第23回 大脳基底核研究会
「基底核とその周辺」
2008年7月5,6日
参考資料1:基底核についての要約と解釈
(参考資料2「小脳、扁桃体」および3「辺縁系一般」は研究会における補足資料)
サル大脳皮質線条体線維の起始細胞は皮質第5層にあり、前頭葉内の主溝を中心としたその周囲領域からは尾状核への、その背側(6野, 前頭前野)や眼窩面に近い腹側野からは尾状核および被殻の両方への投射がみられ、一次運動野、体性感覚野を中心とする領野からは被殻へ投射する。その他に頭頂葉(尾状核と被殻へ)や側頭葉(とくに尾状核へ)からも線条体への投射が存在することが知られている [Kemp and Powell, 1970; Jones et al., 1977] 。
線条体は細胞構築上、アセチルコリンエステラーゼ(AChE)に強く染まる部分と弱く染まる部分に区分される。前者はmatrix compartment (matrix)、後者はpatch compartment (patch) とよばれる。パッチはstriosomesとも呼ばれ、マトリックスにより糸巻き状に囲まれた複雑な配列をしている。ラットの線条体では、胎生13~15日(E13~E15)でパッチが、E18~E20でマトリックスが形成される。その際に黒質からのドーパミン含有入力が最初のパッチ形成に強く関与すると言われている。線条体への入力としては、辺縁皮質関連野からは主にパッチに、新皮質からは主にマトリックスに、また、ドーパミン投射に関しては、腹側被蓋野(VTA)、黒質緻密質(SNc)の背側部からマトリックスに、SNcの腹側部と黒質網様部からパッチへの投射がみられる。なお、腹側線条体に投射するVTAのドーパミン細胞はコレチストキニン(CCK)、ニューロテンシン(NT)を共有する。
これらのパッチ/マトリックス(patch/matrix)構造および伝達物質、含有ペプチドや受容体に関して、意味づけをすることがどの程度できるのか?
線条体内には1種類の投射ニューロンと4種類の介在ニューロンが明らかにされている。投射ニューロンは多数の樹状突起棘をもった中等大の細胞(medium spiny neuron)で GABA作動性、全体の約80%を占め、淡蒼球に線維を送っている。介在ニューロンにはGABA/Pv含有細胞、SOM/NOS含有細胞、カルレチニン/含有細胞およびACh含有細胞がある。そのうちのコリン性ニューロンは大型無棘の細胞(large aspiny neuron)で、全体の2%を占めるに過ぎないが、線条体全体にAChを供給し、その放射状に長く伸びた樹状突起と軸索を介して投射ニューロンに影響を与えており、手続き記憶や条件づけ感覚運動学習に関わる運動制御に大きな役割を演じているという。
大脳基底核・視床・大脳皮質を構成する神経回路のシステムについて、形態、機能ごとに並列的チャンネルを作っており(parallel channeling)、個別的、並列的な情報処理(parallel processing)をする場である考えられている[Alexander et al., 1986]。
この系は閉鎖回路を形成し、便宜上、運動系、連合系および辺縁系の3つの系に分けられる。また、機能・構成上、①感覚運動線条体、②連合線条体、③辺縁線条体に分類される。なお③の辺縁線条体は尾状核頭の腹側部で主に側坐核と嗅結節の深層部に相当し、組織化学的に運動系と連絡しているが、辺縁系のインターフェイスとしても働く中心部 core と、視床下部、扁桃体など辺縁系と連絡する周辺部 shellに分けられる。以下の注を参照。
注:
Ⅰ.運動系ループ
1)運動感覚系ループ(この系は運動の高次機能に関与する)
知覚運動野→被殻(運動系線条体)→淡蒼球外節/内節(GPe/GPi)(外側部、運動系-淡蒼球))→VLo→運動野
2)固有補足運動野(SMA-proper)系ループ
SMA-proper→被殻→GPe/GPiの中間部(補足運動野関連淡蒼球)→VLo内側部→SMA-proper
3)前補足運動野(pre-SMA)系ループ
pre-SMA→尾状核(CN)の外側部→GPe/Gpiの中間部→VApcの外側部→pre-SMA(pre-SMAはヒエラルキーが最も上位にあり、このループは運動のプログラム、準備に深く関与する)
4)運動前野(PM)系ループ
PM→CNの外側部→GPe/GPiの背内側部→VApcの内側部→PM
5)眼球運動系ループ
前頭眼野(FEF)/補足前野眼野(SFEF)→CNの中央部→黒質網様部(SNr)の外側部→VAmc,MDpl→FEF/SFEF
Ⅱ.連合系ループ(この系は認知などの高次の脳機能に関与する)
前頭連合野/頭頂連合野→CNの外側と腹内側を除く大部分および被殻の前部(連合線条体)→SNrおよびGPe/GPiの背内側部(連合系淡蒼球)→MDpcの中央部と一部VAmc→連合野
Ⅲ.辺縁系ループ(この系は動機づけ、情動行動に関与する)
辺縁皮質・扁桃体・海馬→辺縁(または腹側)線条体→腹側淡蒼球→MDmc内側部→辺縁皮質
この情報処理概念を発展させ得る所見として、最近、ミドルトンとストリックはヘルペスウイルスを利用したシナプス越え逆行性軸索輸送法を用いて、サルの前頭前野-基底核-視床-皮質を経由する神経回路を調べ、その中で前頭前野への5つの投射系を明らかにした[Middleton and Strick , 2002]。すなわち、大脳基底核、視床核を経由して、前頭連合皮質内の9野の内側(area 9 m)と外側(area 9 l)、46 野の背側(area 46 d)と腹側(area 46 v)および12野の外側(area 12 l)への投射が存在すると提唱した。
さらにマックファーランドとハーバーは、同じくサルの脳に種々の軸索流マーカーを注入して、このシステムの大脳基底核を中継とする視床と皮質との間の結合様式について、たとえば、VL核は一次運動野、補足運動野、運動前野後部に対して、VA核は帯状回前部、前補足運動野を含む運動前野前部に対して、MD核は前頭前野の背外側部および眼窩面皮質に対して各々投射することを明らかにした[McFarland and Haber, 2002]。
近い将来に脳機能の能動的活動、あるいは精神機能に関する研究の中心をなす課題の一つがここに成立しつつある。実際、カミングスらは、前頭皮質-大脳基底核-視床の神経回路を、ヒト前頭葉内の ①補足運動野、②前頭眼野、③背外側前頭前野、④前頭眼窩面外側部、⑤後帯状回前部を起始とする5系列に分けて、運動機能、認知、情動、動機づけなどの行動認知や精神機能について考察し、その障害として、実行系障害、性格障害、無感動に関わるものがあると推論し、それらを精神異常に結び付けようとしている[Cummings, 1993, 1995, Tekin and Cummings, 2002]。
なお、近年、彦坂グループ[Miyachi et al., 1997; Nakamura et al., 1998, 1999] は、この大脳皮質-大脳基底核連関に注目して、その手続き的(procedural) 運動記憶とその実行系の神経機構を明らかにすべく、このサルの生理学的実験(細胞外記録)を行なってニューロン活動を調べ、① (視覚空間座標でコードされる)学習の初期、つまり新しい課題を獲得しようとする段階では、前頭前野/前補足運動野(pre-SMA)/頂頭葉内側部などの連合皮質-基底核前部ループ(視覚ループ)が関与し、次に ②(運動座標でコードされる)学習がさらに進んで蓄えられた記憶を保持し、読み出し、スキルになった手続きを実行する段階になると、補足運動野(SMA)などの運動関連皮質-大脳基底核中央部ループ(運動ループ)が関与するようになるという結果を示した [Hikosaka et al., 1999; 中村, 2001] 。
これは空間座標と運動座標が並行してセットされているという意味で、「パラレル・ニューラル・ネットワーク(parallel neural network)」と名付けられた理論で、運動記憶が、なんらかの事柄への習熟の度を高め、自在に適応可能になる過程で少なくとも2つの、つまり複数の独立した前方および後方の「大脳皮質-大脳基底核回路」が働き、漸次一方の系列に移行する機構が存在することを予測させる。
大脳基底核は、大脳新皮質、大脳辺縁系、中脳ドーパミン系との密接な結びつきから考えて、認知、運動(能動的活動)、意欲、情動などの「精神機能」を統合する上で重要な位置を占めるにもかかわらず、なお未開拓の研究分野である。大脳基底核の背側部は黒質線条体(A9)系の主な標的部位として感覚運動機能に関係し、他方、腹側部は中脳辺縁ドーパミン(A10)系の主な標的部位として海馬や扁桃体、さらに視床下部と共に辺縁系の中核を成していると以前からみなされてきた。
なお、線条体入力軸索終末には少なくとも大脳皮質由来のグルタミン酸作動性(その受容体はNMDAタイプで、樹状突起棘/スパインの頭部にある)のものと黒質由来のドーパミン作動性(その受容体はD1, D2 グループで、樹状突起棘/スパインの頚部にある)のものがある。これらの入力を受けてGABA 作動性の抑制性投射ニューロンが視床ニューロンおよび大脳基底核あるいは前脳内側基底部の細胞群(マイネルト基底核、ブローカの対角帯、無名質など)内のコリン作動性ニューロンに神経終末を与えている。そして、これらの視床ニューロンおよび大脳基底核ニューロンは、ともに広く大脳皮質に興奮性の出力を与えており、フィードバック的に大脳皮質の働きを制御している。
さらに一言すると、抗精神病薬の作用点(伝達物質の受容体)や疾患モデル動物を用いた薬物作用機序の研究の結果も考慮されて、統合失調症における情動処理障害仮説―視床フィルター機能不全仮説(thalamic filter hypothesis)―が カールソンらによって提唱されて久しい[Carlsson and Carlsson , 1988] 。大脳皮質-基底核-視床-大脳皮質という「再帰性」神経回路(網)のなかで情報統合処理が障害されるというこの仮説は、20年も前に統合失調症障害を見据えた点は注目に値する。現在の神経科学の進歩の上に立って再検討したい。
大脳(新)皮質から直接投射をうける尾状核と被殻(同質の構造体で両者はまとめて新線条体と呼ばれる)から入力をうける淡蒼球(外節と内節とに区分される)からは、視床の運動核(VA/VL)へ投射がある他に髄板内核群や視床網様核(両者とも新線条体にフィードバック様に投射して閉回路を作っている)にも投射がなされている点であろう。とくにこの視床網様核は上行性網様体賦活系(ascending reticular activating system, Moruzzi and Magoun, 1949)の間脳における中継核に相当し、視床から大脳皮質に広汎な覚醒刺激が送られる際に抑制過程として関与すると言われている [Masson et al., 2002] 。
大脳基底核の機能をより簡明に解釈すれば、以下のようになろう。すなわち、運動系においても、知覚系においても、さらには認識・情動系においても、体外および体内からの刺激による過剰で不適切な入力を阻止し、コントロールして適正な覚醒状態を保ち、新皮質とくに能動性機能の場である前頭葉を活性化させることによって、霊長類とくにヒトが日々の行動、生活において注意を集中することに大きく関与するものと思われる。この情報入力に対する抑制機能ないしフィルター機能が傷害されると、雑多な情報を適切に選択することができなくなり、過剰な感覚刺激を処理できず、その結果として能動機能が低下し、思路は乱れて、奇妙な思考形態をとることにもなる。 今後、精神疾患の病態の解析をも新たな視点に据えたい。
以上。
参考資料2:小脳
小脳出力系 ―上行性と下行性
小脳は脳幹、脊髄の広範囲の領野から受ける種々の感覚および運動情報を処理して、それらの結果を前脳、中脳、後脳内の様々な神経細胞群へ送り返している。考えられるすべての神経機能の ―その計算(metric)された上でしかも自動的(automatic)な働きを通して― 円滑な活動に、大きな影響を与えていると言える。
「古い」小脳に属する「前庭小脳」(片葉と小節)のプルキンエ細胞は前庭神経核に直接線維を送っている。これに対して、「脊髄小脳」、「橋核小脳」と呼ばれる領域のプルキンエ細胞は軸索線維をすべて小脳核に送っている。上述の微小帯域(マイクロゾーン)に小脳核/前庭核の細小域が相互に結合することにより密接な対応関係が生じ、機能単位としてのループを構成しており、小脳核でニューロンを代えて後、小脳核からの遠心性線維は、上小脳脚(結合腕)および下小脳脚(索状体)を通って、小脳の外に出る。上小脳脚内の線維群は上行枝(赤核、上丘、視床核、視蓋前域へ投射)と下行枝(一部が延髄まで)に分かれる。下小脳脚の大部分は小脳求心性の線維で構成されているが、小部分が前庭神経核、橋延髄網様体、上部頸髄へ投射する遠心性線維を含んでいる。小脳外側核および中位核から出た軸索線維は上小脳脚を通り、中脳下部で交叉した後、その大部分は反対側の赤核(外側核からは赤核の小細胞部へ、中位核からは大細胞部へ)および視床の運動核(ネコのVL核、VA核;サルのCL核、VPLo核、VLc核)に終わる。内側核(室頂核)から出る線維の主たる標的は、前庭神経核外側核(ダイテルス核)および延髄の巨大細胞網様体核と尾部橋網様体核で部位局在の関係が明らかである。小脳には運動性のほかあらゆる感覚様態の情報が入力されている。出力系としては、視床核からは上行性に大脳皮質に興奮が伝達されるが、赤核、上丘、脳幹網様体、前庭神経核などからは、下行性に脊髄や下オリーブ核などに投射がなされているので、小脳皮質・小脳核・脳幹諸核が構成する神経回路ループの活動は低次から高次にいたるまでの精神活動の形態基盤として重要な視点を提供するものである。
小脳の運動および思考モデル
以上の古典的制御系に上述の皮質マイクロゾーン(微小帯域)-小脳核/前庭核を包括する機能単位(小脳チップ)モデル- をこれから作ろうとする雛形となる元の系と並列的につないで、かつその元の系と小脳チップモデルとの間の出力の差を誤差信号として小脳チップに与えるように組み込む。誤差信号を与え続けると小脳チップの動特性が次第に元の系の動特性に近づいて古典的な制御系を適応制御系に変えることができる。この原理で筋肉骨格系のモデルを小脳内に作り上げ、これに大脳皮質が働いて、小脳を通る内部フィードバックによって、外部フィードバックを置き換えてしまう。これができてしまうと、外部ループなしに小脳の内部モデルの働きだけで筋肉骨格系の動特性を忠実に再現させることができるようになる(「ダイナミクスモデル運動制御系」, forward model)[Ito, 1970、]。その後、川人と五味によって、小脳の中に筋肉骨格系の動特性の逆数を現わす「逆ダイナミクスモデル運動制御系」(inverse model)というもう一つのモデル制御の方式が提案された[Kawato and Gomi, 1992a, 1992b]。伊藤は小脳に見られる上の2つの制御様式の差について次のように説明している。
すなわち、 「ダイナミクスモデル運動制御系は大脳運動野と小脳の中間部との間の関係と、逆ダイナミクスモデル運動制御系は運動前野と小脳外側部の関係と対応しており、運動練習の二つの段階に当てはまるものと思われる。前者の場合は意識的だが、目をつぶったまま、結果を見ないでも正確に思った通り運動できるようになり、後者では運動を繰り返し練習すると、無意識のままでも正確な運動ができるようになると想定できる。」
伊藤はこの運動に関するモデルを思考過程にも拡張、発展させて、モデル思考制御系の考えを提示した(Ito, 1993)。「思考モデルの順モデルを組み込んだ思考系」と「思考モデルの逆モデルを組み込んだ思考系」と呼ばれるもので、モデル作図の上では、「運動野」を「前頭前野」に、「筋肉骨格系の運動作用」を「後連合野の認知作用」に入れ替えた形になっている。この運動から思考へのステップは非常に大きいものがある。すなわち、「①前頭前野が後連合野内の思考モデルに働きかける。②この思考モデルの動特性をシミュレートするモデルを小脳内に作る。③前頭前野はこの小脳内思考モデルに働く。」この過程を繰り返し続けることにより、自動的思考が可能になり、思考モデルの逆モデルが小脳内にできれば、無意識に思考過程が進行する。
長い間、小脳はもっぱら運動機能の制御に関わるとして他の機能の発現については看過されてきた嫌いがあるが、小脳の自律機能/情動機能への関与は下等な動物からみられる。すなわち、これらの動物において既に小脳[Haines, 1984] や扁桃体 [小野と西条、2002]から視床下部への投射の存在が証明されている。なお、霊長類、とくにヒトになると、大脳・小脳関連ループの中に、運動系に加えて思考機能系の問題を改めて設定する必要があろう。サルにおいて小脳外側核(歯状核)から視床を介して前頭葉に到達する神経路は佐々木らによって電気生理学的によく調べられている[Sasaki et al.,1979; Sasaki and Gemba, 1993]。さらに玄番らは、ヒトの発語との類似性を考慮に入れ、サルの小脳の皮質除去手術などを行ない、その際、発声に先行して大脳皮質内に出現または消失する活動電位(field potential) を指標として、小脳からの興奮(皮質表層陰性, s-Nかつ深層陽性, d-Pの電位)を伝える遅い波が前頭葉内、とくにその左側の運動前野、補足運動野および弓状溝下枝後壁(ヒトのブローカ野に相当する領野という説がある)に出現することを観察した。またさらに、この結果を基に、その存在が予測される小脳・大脳投射の持つ役割を発声、発語との関わりのなかで考察した[Gemba et al., 1995, 1997]。
伊藤は「思考は脳内にあるモデル化された概念を動かすことである」という意味の心理学用語を用いて「思考モデル」と呼び、小脳において提示された運動制御系を大脳のモデル思考制御系に適用(対応)させて提案した。すなわち、大脳皮質内でブローカ野を含む前連合野が、認知思考の要素の貯蔵庫であるウェルニッケ野を含む後連合野内に存在する思考モデルに繰り返し働きかけた結果として、ヒトが大脳皮質内活動として思考、すなわちさまざまに考えることをくり返すうちに、小脳と大脳皮質との間を両方向性に密接に結ぶ結合[Evarts and Thach, 1969; Allen and Tsukahara, 1974; Sasaki, 1979]を使って小脳内にそのシミュレートされた思考モデルが形成されてしまえば、何度か、既に経験された思考に関して、改めて大脳皮質内活動をすることなく自動的に思考が進むことになる。
ヒトが何かを考える場合、そこには意識を集中して働かせて自意識にまで発展させ得る、論理的・構築的に行なわれる思考と、自動的、無意識的に行なわれるより単純な思考とがある。しかしいずれにせよ、こうした思考のための情報処理は進化的に最も新しい大脳新皮質で行なわれる。とはいえ小脳のほかに、① 皮質の働きを安定化させる大脳基底核、② 皮質の活動モードを調節する視床、③ 記憶、価値判断、動機づけに関与する辺縁系など、種々の機能を発揮する多くの下部構造がこれを支えているのである。ここで思考作用を脳内の「思考モデル」を操作する一種の行動と考えると、心に浮かぶ様々な要素的な思考過程を組織化して伝達するという、この思考という名の運動が、いわゆる一般の、身体的運動遂行の操作と同様に考えられることになる。
また、その際に、大脳辺縁系と結びつきの強い側頭連合野は思考内容についての喜怒哀楽を伴った価値評価に深く関与することになる。広義の反射、種々の知覚作用、習熟した手続き記憶、情動を伴う条件付けなどの諸要素を変数とする関数として扱うことによって、これらを脳の高次機能を種々の異なる種類の神経回路モジュールが作り出す動的な信号として捉えることができよう。認識や記憶や情動・意欲の行動をこれらモジュールの複雑な組み合わせとして理解することが、すなわち脳の高次システムを解明することにつながる一つの道であろう。
以上みてきたように、協調には仮にそれが言語であれ、思考であれ、行動であれ、意図的、随意的な協調と、自動的、不随意的な協調という2つのシステムがあり、これらを結びつけて統一する形態的な構成系として、小脳・大脳関連ループ系の存在が重要なものとして注目される。サルでの所見ではあるが、小脳半球の外側部は大脳皮質の運動野外側部、運動前野および前頭前野との間に、他方、小脳の中間部は大脳皮質の全運動野(とくにその中間部)との間に相互結合が認められる[Sasaki,1979, 1986]。そして、この小脳・大脳関連ループは前脳の発達を待って成熟したものとなる。それだけに、この系、とくに小脳の外側部と大脳の前頭前野を結ぶ系は、とりわけてヒトの脳において特徴的かつ重要な系となるものである。
「メトリカル」なリズムと「ノンメトリカル」なリズム
小脳が備えている認知機能(注意、記憶、言語、感覚、刺激受容)に関係する、もう一方のリズムないし時間情報の問題について考察する。すなわち、坂井らはヒトに7つの音を異なる時間間隔で聞かせ、12秒後にそのリズムをボタン押しによって再生させた。このようにしてリズムを短期に記憶し、保持しているときの脳内の活動状態を同じくヒトでf MRIを用いて研究した(Sakai et al.,1999a)。 彼らの研究結果から解釈すれば、リズムはインターバル比率に依存しており、その構成の比率により2通りのパタンがあるというが、基本的にはインターバル比率、つまり時間を一定比率の打点として捉えたものを人間はリズムと呼んで来たのであり、しかもこのリズムを感じ取って二様に区分して来たのである。だがともかく問題の実験によれば―
①左側の運動前野、頭頂葉と右側の小脳前葉外側部は1: 2: 4および1: 2: 3のリズムに反応し、②右側の前頭前野、運動前野、頭頂葉および両側の小脳後葉半球部は1: 2.5: 3.5のリズムに反応する(ニューロン群を持つ)。
①②は共に音と音との、座標で言うならば横軸の関係に相当するもので、①はその場合の時間間隔が一定(metrical)であり、②は不定(non-metrical)である。ただし、①の一定比率の打点(時間)は、当然打たれるべき次の点に対する予測を前提させるから、その予測からして、打点、すなわち音そのものの存在を予測させることになる。これを言い換えれば、次の打点(音)の存在(非存在)の表象に関わる神経作用であると言うことになる。つまり、当然あるべきものがあるべき場所にあるかないかの判断ないし表象に関するものであり、この、あるべき場に表われるべき点(音)をくり返し予測(感知)して行なう認知作用を一種の、一定時間比率による認知作用の積み上げ式(階層的表象構造)と呼ぶことができる。
それに対して②の不定的、非一定比率の場合は、①にみられる一定の予測が立たないため、①に比してより多くの緊張を伴って、むしろ非時間的に、ということは予測認知活動がmetrical(リズミカル)に予測点を打って自動的に進行して行くのとは大きく異なり、不測的な次の打点を待ち受け、打点される毎に、その点がこれに先立つ打点とどのような関係(比率)にあるかを計算し、割り出して行く認知活動になる。そのため、個々の打点に即して、その都度打点が、待ち構える認知によってキャッチされ、その都度異なる待ち時間が認知されて行くことになる。これは①に対して明らかに非階層的=非積み上げ的であり、それだけに神経作用のエネルギーは大となり、緊張・興奮が伴う。
新奇なリズムと耳慣れたリズム(小脳後葉と小脳前葉)
これを一般的に解釈することはかなり難しいが、日常的には、①はわれわれの生活(環境)に自然な馴染みのある「整数倍」リズムであり、長期記憶として小脳前葉とそれに関連する領域(左側大脳皮質運動前野など)に蓄えられているのであろう。これに対して、②はそうした自然の秩序から外れる、日常にはない一瞬戸惑いを感じさせる「非整数倍」リズムである。この「新奇」なリズムは小脳後葉でキャッチされ、そのリズムが馴染んで自動化されると小脳内の活動が前葉に移行する。この際に活動する大脳皮質領域は右側前頭・頭頂葉である[Sakai et al. 1999, 2000]。ここで新旧の事項の運動記憶ないし認知機構に関わる神経回路の問題として、彦坂グループによって明らかにされた、手続き記憶学習の獲得と実行に関わる大脳皮質-基底核ループに属する複数の独立した系「パラレル・ニューラル・ネットワーク」[Hikosaka et al., 1999;坂井, 2000]と結びつけることができよう。
小脳系・線条体系・大脳皮質運動系
すなわち、一方は ①課題に慣れて習熟した段階に関わる、いわば運動実行系の、右側小脳前葉外側部―左側補足運動野などの運動関連皮質―大脳基底核中央(および後)部のループ(運動座標系)、他方は②運動のタイミングの調節などの新しい課題を獲得しようとする段階に関わる、両側小脳後葉半球部―右側側頭葉内側部などの連合皮質―大脳基底核前部のループ(視覚座標系)という2種類の機能系列に属する回路(ループ)が脳内機構として存在することになり、それらは小脳系、線条体系および前補足運動野や運動前野が関与する大脳皮質運動系という形態的に異なる3つの系が互いに関連づけられて考察され得るようになる。小脳の推尺作用として考えられてきたいわゆる空間的概念のみの捉え方は、筋肉群が連続的に空間・時間的に変化する運動のほかに、言語や思考の分野においても、時間の制御機構ないし小脳による時間的統御の機能としても新たな視点が当てられる段階に到達しているように思われる。
随意運動系と不随意運動系に認知・情動系を包括する「運動のゲシュタルト」
ここで強調して置きたいことは、感覚性言語野と結びついた受動的な認知と情動表現が、運動性言語野と関わりをもつ能動的な情動行為と結びついて、その結果この前頭前野内で「組み換え処理」がなされ、然る後に情報が前補足運動野に伝えられ、次いで意欲に関わる帯状回(帯状皮質運動野前部および後部)との結びつきをもった補足運動野(supplementary motor area)、そして第一次運動領(4野)へと興奮が入力され、錐体路系が作働される点である。
やや詳しくなるが、前補足運動野から補足運動野を介して第一次運動領への神経回路の間に間接的に腹側および背側運動前野(ventral and dorsal premotor areas)が関与していることは留意すべき点である(丹治、1999)。なお、4野以外の広い範囲の皮質領域からも皮質下の運動系組織である線条体への投射が存在する(動物が高等になるにつれて運動系関連皮質以外のところからも投射されるようになる)。
この線条体運動系は他に小脳系、網様体系の「不随意運動系システム」をも含めて錐体外路系と呼ばれ、全体的な身体的なバランスを統御・調節的に司るところである。言葉を換えていえば、「運動系のゲシュタルト機構」といえよう。このように運動系は単に随意運動を司る錐体路系のみでなく、上述の高次運動野といわれる運動関連野皮質および大脳基底核を中心とする大脳皮質・線条体路、黒質・線条体路、腹側被蓋野・側坐核路、扁桃体遠心路(視床下部、側坐核、中隔核などへ線維を送る)と密接に関連しているので、芸術や思索に結びつく高次神経活動、すなわち、精神活動を研究する際にこの点を含めて総括的に考案することは重要な視点である。
付言すれば、ピアノやヴァイオリンを学習するタイプのいわゆる「体で覚える技術的性質をもった」記憶の習得は小脳・脳幹を中心になされており、その学習の習熟のプロセスは視床運動核(VA, VL)を介して順次大脳皮質の運動関連皮質に伝えられるものである。複雑なことにVA/VL核には小脳から小脳核を介して興奮性入力が、また大脳基底核から淡蒼球(内節)を介して抑制性入力が情報として集まって来る。しかし、この2つの入力系は互いに独立に処理されていると考えられ、両者が1つの視床ニューロンに収束することは稀である[Ilinsky and Kultas-Ilinsky, 1984]。
さらに視床―皮質投射にもこの独立性はおおよそ維持されており、主として、大脳基底核からの信号はVA/VLを介して補足運動野に、小脳からの信号はVPLoを介して運動野およびVLmを介して運動前野に送られるとされてきた[Schell and Strick, 1984]。この見解は大筋で正しいが、細かいところではなお議論の余地が残されている。すなわち、その後のマテリらの研究によれば、従来の補足運動野はさらにF3(固有の補足運動野、SMA-proper)とF6(前補足運動野、pre-SMA)との独立した2つの領野に区別されるが、両領野ともに淡蒼球からも小脳からも入力を受けるという(Matelli et al., 1995)。
このように、小脳系と線条体(または大脳基底核)系とは独立性が強く、大脳皮質を介する回路で結びついていると考えられてきたが、2つの錐体外路運動系はシナプスを超えて直接的につながっていることが、最近のラット(Ichinohe et al., 2000)およびサル(Hoshi et al., 2005)を用いた研究で明らかにされた。とくに、Hoshi ら(2005)は赤毛ザルの基底核である、被殻と淡蒼球に狂犬病ウイルスを微量注入して、超シナプス性逆行性に標識される神経細胞を歯状核を主とするすべての小脳核に認めることにより、霊長類において小脳-基底核投射がかなりの強さで存在することを証明した。この発見の意義は大きく、「能動性」高次機能の新しい展開の基礎となるであろう画期的な仕事として注目される。
総体としてみた運動発現のシステム
以下に大脳皮質から脊髄に至る運動下行系の幹線神経回路を示す。視・聴覚系入力によって運動系に刺激が加わり、テンポとリズムがヒトの身体内に発現する。発生学的にみると、線条体/間脳→脳幹→脊髄のルートが始めに歩行もしくはプリミテイヴな踊りのような身体の活動として歩行機構と結びついた形で発生し、辺縁系・情動系により駆動され、感情移入された情動の昇華の源が見い出されることになる。実際、尾状核の視覚性記憶ニューロンは、運動の報酬スケジュールによって強く修飾されることが示されている[Kawagoe et al.,1998]。サルを用いたこの実験は、最初に視野の中心部を注視させておいて、光を視野の上、下、左、右にランダムに点灯させて眼球運動を起こさせ、この際の大脳基底核ニューロンの活動を記録するものである。この時、或る特定の方向に眼球を動かした(外眼筋を自己の意志により活動させた)時のみ甘いジュースという報酬が与えられる。
視空間認知に価値判断(動機)がリンクし、大脳基底核が関与して運動系へ変換される。
この実験のパラダイムは、視覚系が空間を認知し、価値判断や動機とリンクさせて運動系に変換する時、どのように大脳基底核のニューロンが関与しているかを調べようとしたものである。この大脳基底核領域は、扁桃体から情動系の入力が、またさらに新奇物体に対する意欲的行為に関与する中脳の黒質からのドーパミン入力が、終止する場所である。
実験結果は以下のようである。すなわち、この領域のニューロンは、サルが、形象を自己にとって意味のある空間内に認知し、その方向に向かって意欲的に行動を開始したときに活動し、大脳基底核(実験として尾状核を用いた)は報酬を期待することによって動機づけを評価し、その結果眼球を動かすという運動系の決定に関与していることを示すものであった。
報酬期待時における前頭前野ニューロンと大脳基底核ニューロンの活動性の違い
これに関連して触れておきたいこととして、このように尾状核では、報酬が予知され、期待されたときに、ニューロンの活動性が全体として(底上げされた形で)上昇するが、これに対して、前頭前野外側部のニューロンは動機によって全体的に活動性が上昇するのではなく、条件/状況に応じて弁別性が高まるという結果が同じく彦坂グループ[Lauwereyns et al., 2002; Kobayashi et al., 2002]によって明らかにされている点である。
情動報酬系が大脳皮質言語系の段階にまで進む
ここでコメントしておくが、彦坂らはサルを用いた実験で、手足や体幹など全身的運動を調べているのではなく、眼球運動としての外眼筋の活動に代表させて、認識や動機づけによって行動がコントロール(制御)されるか否かを調べているのである。これは実験データを得る際のニューロンの背景の活動による乱れ、いわゆる「雑音」、を除去して観察する点で優れた手法である。既に、中脳ドーパミン系の大多数のニューロンは、報酬期待性の視覚性のみならず聴覚性の刺激に対しても相動性(phasic)の発火(活動)を示すというデータが得られており、情動報酬の過程と接近行動の学習に関わっていることが知られている[Schultz et al.,1997; Schultz, 1998]。従って、線条体内に聴覚性記憶ニューロンも同様に存在し、情動に関わる系によって(リズムを持った運動としての、たとえば演奏を含む)行動が強く影響されると推察される。このように考えると、視覚系における形や色の表現に類似した脳内機構、すなわち言語の発音(語ること)の機構とは異なるニューロンのコントロールセンターなるものが、聴覚系における音の表現にも存在するように思われる。もっとも、このコントロールセンターを特定するためには、なんらかの響き(振動)が、何を根拠としたときには言語として、また別のときには言語とは異なるものとして発現するかを明確にする必要があろう。果して人間は、どのような振動形態を言語と呼ぶのであろうか?
このように、大脳皮質とくに新皮質が発達した動物-哺乳類-の段階になると、その動物の脳髄には、リズム・テンポの表現に加えて、動と静、速と遅、長と短、さらに美と醜、純と不純、協和と不協和など対比的関係を持った質的に発展した表現方法が発現されるような機構が形成されてくると考えられる。最も上位にある大脳新皮質はこのプロセスの中で下位の線条体の運動を中心とした並列系ループの中でこの系をコントロールする役目を演じる。聴覚領、視覚領の持つ知覚、認知、認識記憶のシステムは上にみたように、言語系(第二信号系の確立)と結びついて、後連合野の内でより高い段階に昇り、それが背側路および腹側路を通り、前頭葉、とくに前頭前野に伝わり、そこで文脈の中に含蓄されている意味内容(connotation)の変換ないし組み換えを起こして、運動系をゆり動かす。
扁桃体への入力(系):
扁桃体には、味覚、嗅覚、内臓感覚、聴覚、視覚、体性感覚などあらゆる種類の刺激が、嗅球や脳幹から直接的に、そして視床核(視覚、聴覚などの特殊核近傍ニューロン群)を介して間接的に入力される。その他に、大脳皮質内で処理された情報および海馬からの情報が扁桃体に入ってくる(ちなみに、これらの領域間の情報の流れは両方向性である)。この後者の入力は梨状前皮質、嗅内野(28野)、海馬台(ヒトの海馬旁回近傍)、帯状回(とくに24野)、側頭葉、前頭前野から扁桃体の基底外側核に至るもので(Kawamura & Norita, 1980; Aggleton ら,1980)、前者の皮質下からの入力に比べて時間的に遅れて、より適正かつ精緻な情報が入力される。これら2種類の粗と精、「低位」と「高位」の経路を通過した情報が扁桃体で出会う。そこでは、環境に対して瞬間的、反射的に反応した生得的な生体反応は、皮質レベルで認知された「高次」の情報に基づいて、益(報酬性)か害(嫌悪性)であるかが環境適応的に判断され修正される。同時に、海馬は新皮質からの認知情報を受ける前に情動的情報を含んだ扁桃体からの入力を受ける。この海馬・扁桃体間の関係は相補的である。
なお、モノアミン系の入力として、
1)ドーパミン系: 黒質緻密部(A9)および腹側被蓋野(A10)から起こる中脳辺縁皮質系の一部が、
2)セロトニン系: 中脳背側縫線核(B7)および正中縫線核(B8)から起こる上行性セロトニン束の一部が、
3)ノルエピネフリン(=ノルアドレナリン)系:青斑核(A6)から、4) コリン系:マイネルト基底核(Ch4)からの入力が扁桃体に送られてくる。
扁桃体への入力系
扁桃体には、味覚、嗅覚、内臓感覚、聴覚、視覚、体性感覚などあらゆる種類の刺激が、皮質感覚野を経過せずに、脳幹レベルから直接的にまたは間脳の視床核を介して間接的に入ってくる。環境からの情報を将にそのまま感覚的に受容する生(なま)の粗な感覚である。その他に、大脳皮質を経由していわば高次元で処理され、知覚され、認知された結果が扁桃体に入ってくるものがある。この後者の入力系はその伝達経路の故に必然的に、時間的に僅かに遅れて伝達されるが、そのことによって、適正かつ精密な情報として入ってくる。すなわち、扁桃体の基底外側核は、梨状前皮質、嗅内野(28野)、帯状回(とくに24野)、側頭葉、前頭前野からの皮質遠心性線維を受ける(Norita and Kawamura, 1980)。以上の粗と精、原始的と識別的、低次と高次という2種の情報が扁桃核内で遭着する。
扁桃体からの出力(系):
大別して3つを挙げる。すなわち、①扁桃体の中心核(および一部、内側皮質核)から起こり尾状核と視床の境界部に沿って背後部から前腹方へと走って中隔核、視床下部(前核、腹内側核、弓状核)、内側視索前核などへ終止する分界条(stria terminalis) という神経線維束、および②主として基底外側核を出て内側に走り、側坐核や外側視床下部から内側部にかけて分散状に分布する腹側投射系(ventral pathway、または腹側扁桃体遠心路ventral amygdalofugal projection)と呼ばれる投射路、さらに③広範囲の大脳領域へ終わる皮質投射がある。とくに側頭葉(TE野、Boninと Bailey, 1947,による略字命名、以下同様)梨状葉皮質、前帯状回、眼窩面皮質(OF野)との結びつきは強い。側頭葉については、扁桃体の内側基底核はTE野の腹側域とのみ、外側核はTE野全体と各々結びついている(Saleem, 2000)。
分界条(扁桃体の情報を視床下部へ伝える経路)
扁桃体の働きに、たとえば芸術→情動→療法といった、現在社会的関心を集めている音楽療法といったものの経緯の有効性を探る鍵の一つが存在していることは間違いあるまい。ともかくも、ここで行なわれるこの価値判断の機能の結果は、主として扁桃体の皮質内側核から起こる分界条(stria terminalis)を経由して中隔核、視床下部(前核、腹内側核、弓状核)、内側視索前核などへ伝えられ、小部分は主として基底外側核から起こる腹側扁桃体遠心路(腹側投射系、ventral pathway)により視床下部外側部に伝えられる。
視床下部は摂食、性機能、水分代謝などの自律機能や脳下垂体の機能を促進または抑制する内分泌調節などを司る生命維持に関わる重要な領域で、外的または内的な誘因が存在するとき、視床下部への刺激が来たとき周囲への働きかけの動因となり、これによって動物は行動(行為)を起こすことになる。扁桃体から入力される線維束である分界条の形成については相沢らによれば、マウス胎生14-16日で核集団を形成中のニューロンから軸索が束化を形成し、或る特定の細胞群(LOT細胞)をマイルストーンとして尾状核と視床の境界部に沿って背後部へ、また前方および腹方へと走り、形成過程の視床下部領域に侵入する。そして腹内側部(「満腹中枢」)や外側野(「摂食中枢」)のニューロンとシナプスを作る。
これらの領域にあるニューロンはグルコースに反応するが、同時に身体の脂肪細胞から分泌される肥満因子レプチンに対して腹内側部には促進的に、外側野には抑制的に反応する受容体を持つものも存在する(後述参照, 25節)。このほか視床下部には、大脳皮質、海馬、網膜、脳幹などから種々の線維が入力する。すなわち大脳皮質とくに前頭葉(6野)からは内側乳頭体核や腹内側核へ、海馬からは脳弓を経由して外側乳頭体核へ、網膜からは視交叉上核へ、さらに脳幹からはアミン系線維の視床下部の広い領域への投射が行なわれている。このうち、とくに大脳皮質-視床下部神経路に関しては、ラット、ネコ、サル、ヒトとおそらく動物の発達段階に応じた線維結合の状態変化が認められると考えるが、今後の研究に期待したい。
参考資料3:辺縁系
海馬―嗅覚、てんかん、記憶―未知なる皮質の原型
情動機能と認知機能はともに、脳の高次神経の活動を担う諸要素の中でも主要な二大機能を成している。われわれの脳は、外部環境および内部環境に関する情報のなかから生体にとって意味のある情報を認知し、過去の体験や記憶情報と照合して、その事象が自分にとってどのような意味を持つのか、すなわち外界からの刺激によってもたらされる情報が報酬性(益、快情動)か嫌悪性(害、不快情動)かを判断している。記憶や情動は海馬や扁桃体を主たる構成領域成分とする大脳辺縁系の機能と密接に関係している。
海馬(hippocampus、ギリシア神話上の動物)はアンモン角(Cornu Ammonis,エジプトの神)とも呼称され、原始皮質 (archicortex) と呼ばれる系統発生的に古い皮質に属している。ヒトでは側脳室下角の床部に沿って前方・内方に折り込まれた形に入りこんでおり、内側壁の大部分を占める。細胞構築学的には、表層から深層(脳室面)へと順に、(外)叢状叢、錐体細胞層、多形細胞層の3層構造から構成されている。それぞれ新皮質の4,5,6層に相当すると考えられている。一般に叢状層に求心性線維が入り、錐体細胞層および多形細胞層から遠心性線維が出ている。髄鞘構築学的にはより複雑な外観を呈する。すなわち、側脳室の方から順に、脳室上衣層、海馬白板、上昇(行)層または内叢状層、錐体細胞層、放射(線)状層、網状層、分子層または外叢状層、内髄層または帯状層に分けられる。
組織学的に歯状回は、表層から深層へ、辺縁層、分子層、顆粒層、多形細胞層または錐体細胞層と区分された明瞭な層構造を呈しており、この錐体細胞層は海馬のCA4の同名の層に移行している。他方、海馬旁回は皮質の幅も広くなり層的分化も進み6層構造を呈するが古い皮質に属し、その4-5層は海馬のCA1の錐体細胞層(Lorente de No, 1934)に連続している。なお、嗅内野は梨状葉皮質の後部の大部をつくりあげている。
注]: 解剖学名(ラテン語)を参考に。
海馬: ①脳室上衣層 str. ependymale、②海馬白板 alveus hippocampi、③上昇(行)層str. oriensまたは内叢状層internal plexiform layer,④錐体細胞層str. pyramidale,⑤放射(線)状層str. radiatum,⑥網状層str. reticulare s. lacunosum,⑦分子層str. moleculareまたは外叢状層external plexiform layer,⑧内髄層str. medullare involutaまたは帯状層str. zonale。
歯状回: ①辺縁層str. marginale,②分子層str. moleculare,③顆粒層str. granulosum,④多形細胞層または錐体細胞層str.pyramidale。
組織構造および神経回路については他書[たとえば、川村と小野、1987;石塚、2002]を参照されたい。海馬は終脳の蓋板に続く半球内側面の部分が翼板の肥厚によって発生の早い時期に形成される大脳皮質の一部である。哺乳動物の海馬は、発生初期には脳梁の腹側に位置しており、脳梁が背尾方に発達してくる時に同伴して次第に発達し、尾方では脳梁膨大の腹側で狭い小帯回となる。さらに腹側前方に進んで数珠玉を並べたような外観を呈する歯状回となり、また側脳室内に足を突出した形の海馬足(内部の構造が固有の海馬あるいはアンモン角といわれる部分である)を形成する。
海馬の構造と機能
海馬は嗅脳に属するとして、サカナやカエルのレベルで嗅覚との関連がしばしば強調されるが、少なくともヒトの海馬は、嗅覚機能にとって大きな役割を演じるものではない[Brodal,1947]。嗅脳(rhinencephalon)という術語は不幸なことにいつも同じ意味で用いられている訳ではない。系統発生学的研究を基礎にして、嗅脳は通常外套部の原始皮質(または古皮質)と旧皮質および中隔野、嗅結節、嗅索、嗅球など終脳基底域の部分を一括とする。その上この術語は生理学的意味を言外に含んでいる。魚類と両生類では、終脳の主体は嗅覚インパルスを外套部(pallial part)に伝える求心性線維であり、哺乳類にみられるような皮質様構造の特徴を示してない。
比較解剖学的には、両生類の外套部(pallium)は、内側部(archicortex、原始皮質、海馬の原基)、外側部(paleocortex 、梨状葉皮質)、中央部(neocortex、新皮質の背側皮質)の3つに区別される。爬虫類では、皮質構造はなお原始型であるが3区分は一層はっきりする。哺乳類では背側皮質が著明に発達し、嗅覚性インパルスの分布は大方paleocortexとarchicortexに限局されるようになるが、その発達・分化は下等哺乳類どまりで、高等哺乳類でさらに発達することはない。下等動物において第一に嗅覚に関係するとみなされる脳のpaleocortexとarchicortexの部分は、哺乳類そしてとくに高等哺乳類において他の重要な機能にとって代わった。
Archicortexにはヒダが形成され、海馬溝の発達により、海馬として側脳室の内側壁の中に突出する。新皮質の発達の結果、paleocortex, archicortexは内方に押しやられ、ヒトでは大脳半球の内側面にみられる。半球皮質の後頭極が発達すると、paleocortexとarchicortexは後方に引かれ、その後、側頭葉の発達にともない再び前方、腹方に引かれる。その結果ヒトを含む哺乳類のarchicortexとpaleocortexは、環状構造を示す。 archicortexは、海馬と歯状回によって代表されるが、この構造は、この環状の線lineの全長にわたってみられる筈である。しかし、両半球新皮質間を結ぶ脳梁の発達によって、この位置にあるarchicortexの部分は著しく縮小し、脳梁灰白質(induseum griseum)と呼ばれる構造として残る。脳梁膝の下方(腹側)にある終板旁回は海馬の最前部とみなされている。paleopalliumは哺乳類で後方に向かって発達し、梨状葉を形成するが、ヒトでは海馬旁回の大部分がこれに相当する。帯状皮質は旧皮質(paleocortex)と新皮質(neocortex)の移行皮質と考えられている。上述の発生学的記載は模式図的ではあるが、哺乳類やヒトでみられる、長いカーブをえがく脳弓や分界条のような奇妙な線維結合の像を説明するのに役立つであろう。
このように、海馬は後方に発達し、それに伴って前方部は退化している。海馬はその発生初期から乳頭体と一部で結合しているが、新皮質の発達に伴ってこの結合部分の内に割って入った形で脳梁が膨大化し後方に発達するため、この結合も伸張されて脳弓とよばれるアーチが形成される。海馬は短期記憶[Scoville and Milner, 1957; Zola-Morgan and Squire, 1986]や外界との関係で場所を認知する[Morris et al., 1982; O’Keefe and Nadel, 1978]機能をもつ領域として注目されてきたが、最近の研究では場所細胞(place cell)の活動は直前のエピソード(短期記憶)に関わる認知機能が反映される学習機能にも関係することが明らかにされている[中沢、2002、参照]。なお、このような記憶に関する海馬の役割については、されるように、分子、遺伝子レベルでの研究が進んでいる(利根川グループ、α-CaMKII、およびリベイロ(Ribeiro)グループ、zif-268 )。
扁桃体―大脳基底核の仲間で八面六臂の情動の重鎮
一方、哺乳動物の扁桃体(組織学的には扁桃核という)は、尾状核や被殻と同様に、半球胞の腹側壁が側脳室の内腔に隆起状に発達する神経節丘(ganglionic eminence)の最内側部と間脳との間の限局された神経上皮から生じる(マウスでは胎生13~14日頃)[湯浅と八木、1998]。側頭葉が形成されるにつれて神経節丘の腹側が前方に移動し、側脳室下角の前端の前上部に扁桃体が位置するようになる。ヒトの扁桃体は側頭葉前部の海馬旁回鈎(海馬旁回の前端が後外側に曲った部分)のすぐ下にみられる。一般に扁桃体と海馬は「関係が深い」 と考えられているが、発生の過程を調べてみると、たまたま最終的に定着した位置が比較的接近しているに過ぎず、両者は互いに独立分離して発達した構造物である。扁桃核は内側核、外側核、基底外側核、中心核、皮質核などの幾つかの亜域(亜核)に分かれている。比較解剖学的にみて、嗅覚との関連で発達してきたと考えられている扁桃体(およびその周辺皮質部)の亜核間の関係が高等動物になるにつれて、より複雑になっていることは興味深い。動物種間の相違はあるが、扁桃体は、皮質内側核群と基底外側核群とに大別される。基底外側核群(基底核と外側核)は動物が高等化するにつれて発達し、ヒトで著明である。一方、内側核、中心核および皮質核は逆にヒトの場合に発達状況は悪い。なお、鳥類以下の原始線条体(または旧線条体)は哺乳類の扁桃体に相同とされている。
大脳皮質との関係をみると、ネコやサルの所見から腹側側頭皮質、前頭葉眼窩面皮質、帯状回から嗅内野(entorhinal, 28野)や嗅周野(perirhinal, 35野)への投射(注: これは両方向性である)、さらに28野と35野から海馬歯状回への貫通線維の投射が明らかにされている[Goldman-Rakic et al., 1984 ]。また、眼窩面皮質および嗅内野を含む側頭葉皮質と扁桃体との間には直接の相互結合がみられる。種々の感覚性刺激の海馬への入力は嗅内野など海馬周辺皮質を介してみられるが、扁桃体への入力は間脳、中脳の幾つかの神経核(視床諸亜核、視床下部の腹内側核、黒質、縫線核など)や脳幹内の結合腕傍核、青斑核などからの直接の投射が存在する。海馬と扁桃体は発生学的にも機能的にも異なる構造物であるが、この両構造物間の線維連絡は、少なくともサルの段階で明らかに存在する[Aggleton, 1986] 。大略、扁桃体の外側核・副基底核からは嗅内野へ、基底核・副基底核からはCA1/CA3へ投射がみられ、逆に海馬台・CA1からは扁桃体の基底核・皮質核に、嗅内野からは扁桃体の基底核・外側核への投射が存在する[Amaral et al., 1992; Fig.6 を参照]。現在、連合野を含む大脳皮質や海馬・扁桃体の特定領域および視床下部諸亜核との間の連絡を究明すべく多くの研究がなされている[LeDoux,1998; Aggleton, 2000; 松本と小野、2002]。
大脳辺縁系の俯瞰と神経連絡
海馬体(海馬・歯状回・海馬台の総称、hippocampal formation)・扁桃体・視床下部(当然乳頭体も含む)・中隔野・嗅内野など(固有辺縁系領野limbic structures proper)の間の結合関係、さらには、大脳基底核、視床核の一部、中脳辺縁系野・側頭葉極部など(旁辺縁系領域paralimbic areas)まで含めた神経連絡路は複雑である。研究方法による所見の相違や動物種間の差などを遂一考察してまとめあげることは困難である。小異を捨てて重要な点に注目して要述的にまとめてみたい。以下に、大脳辺縁系についての鳥瞰図(図A)を示し短い説明を付けておく。
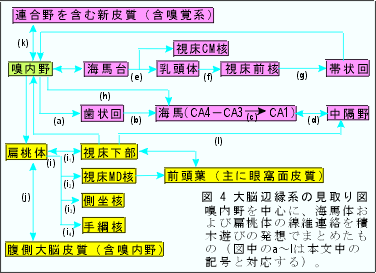
(a):嗅内野(皮質の2層と3層に神経線維の叢がある。詳しくは、細胞構築学的にも線維結合の上からも、内側部-28a野-と外側部-28b野-とに分けられる)からの内側および外側貫通線維perforant path。海馬台を通り抜けて海馬溝を越えるのでこの名がある。主要なものは歯状回の顆粒細胞の尖端樹状突起が存在する分子層の外層(外側貫通線維)と中層(内側貫通線維)に終わる。exteroceptive の情報を運ぶ。なお、内層には対側歯状回からの交連線維が終わる(海馬采からの線維とともに海馬交連として入る)。
(b):歯状回の顆粒細胞からおこりCA4,CA3の錐体細胞の樹状突起に終わる苔状線維とよばれる線維。
(c):CA3と(おそらく)CA4(歯状回のhilus域)の大型錐体細胞の神経突起の分枝がCA1の小型錐体細胞の樹状突起(網状層)に終わる。Schaffer線維という。
(d):主としてCA3よりおこり中隔の外側核に終わる。中隔・海馬路はコリン作働性線維を多く含み内側核からおこり海馬内の広範囲の領域(上行層)に終わる。interoceptiveの情報を運ぶ。
(e):いわゆる海馬乳頭体路で海馬台からおこり脳弓を通って乳頭体外側部に終わる。アンモン角(固有の海馬)からの投射はない。
(f):Vicq d'Azyr束ともよばれ乳頭体の主として内側部からおこる。細かくいえば、視床前核は乳頭体の内側核から同側性に、外側核から両側性に線維を受けている。
(g):視床前核群(とくにAM核、AV核)から帯状回皮質への投射には部位局在の関係が存在する。
(h):嗅内野および(おそらく)海馬台から白板線維(alveus)として海馬(主としてCAl)に終わる。貫通線維の一部も終わる。標的はバスケット細胞と(おそらく)錐体細胞の基底樹状突起。
(i):扁桃体からの皮質遠心性投射(発生的に古い皮質内側核群との結合が強い)
(i1):分界条および内側前脳束を通る。視床下部腹内側核(VMH)に多く、また外側核(LH)にも終わる(他に視索前野、分界条床核にも終わる)。視床下部扁桃体投射は、主として皮質内側核群に終わる。
(i2):下視床脚を通り、MD核の発生的に古い部分である内側部(大細胞性領域)に投射する。この投射域は嗅覚野が存在する眼窩面皮質との結びつきが強い。
(i3):連合縦束を通る。一部が尾状核腹側部、中隔核にも終わるが主たる終止域は側坐核である。なお、側坐核は黒質内側部および中脳腹側被蓋域からドーパミン含有線維を受けており、被殻腹側部を介して中脳網様体の腹内側部へ、線条体からの線維とともに投射している。
(i4):下視床脚を通る弱い投射である。なお、手綱核は視床髄条を介して中隔核、視床前核、外側視床下部域、対角帯核、外側視索前野からの神経線維を受けている。また反屈束(手綱脚間路)を介して中脳の脚間核へ線維を送る。
(j):扁桃体-大脳皮質間結合(発生的に新しい基底外側核との結合が強い)。側頭葉前部、前頭葉眼窩面皮質、帯状回と相互に結合する。
(k):(j)と同様な領域との相互結合の他に比較的広範囲の新皮質領域からも嗅内野への投射がみられる。前頭葉からの線維のうちかなりのものが帯状束内を通路とする。
(l):弱い結合が、おそらく存在する。
大脳辺縁系を、①海馬台-乳頭体系、②海馬-中隔系、③扁桃体-視床下部系の3系に分けることができる。①と②を海馬系としてまとめれば、③の扁桃体系と対比させられて2つに大別できよう。
海馬体(歯状回・海馬・海馬台)の入・出力、(補筆)
上に大脳辺縁系を図解してまとめた形で記述したが、とくに表記のテーマについて改めて説明を試みる[Gloor,1997 ; 松本と小野、2002を参照]。海馬傍回(嗅内野 海馬台の一部)から貫通線維を通って海馬体の入り口の歯状回に入ってきた信号は、歯状回内の顆粒細胞の樹状突起にシナプスを介して情報を伝える。この部分は盛んに神経細胞の新生が行われているところである(別記 前記?)。顆粒細胞からは苔状線維と呼ばれる軸索突起が伸びて、海馬(アンモン角)のCA3の錐体細胞の樹状突起にシナプス結合する。CA3の錐体細胞の軸索突起はシェファー線維と呼ばれCA1の錐体細胞に、一部は海馬采に投射を行う。CA1,CA3領域、とくにCA1領域はLTPの研究が記憶との関連で盛んになされた部位である。エピソード記憶などの陳述記憶や場所を覚える記憶の初期段階の事象がここで完成すると考えられる。この初期記憶の段階が長期的に保持され、想起されるためには次の二つのステップが起こらなければならない。第一はこの記銘と呼ばれる過程(プロセス)が繰り返されること。第二はある段階でその情報が長期記憶として海馬体以外の場所、すなわち新皮質の連合野領域に蓄えられることである。この神経基盤を明らかにするために以下の記述を行う。
第一の回路には、① 海馬体(歯状回→CA3→CA1)⇔ 嗅内野、② 嗅内野→海馬体→乳頭体→視床前核→帯状回→嗅内野があり、第二の回路には、③ 海馬体→嗅内野→大脳皮質連合野、が考えられる。①は貫通線維、苔状線維、シェファー線維が関与している系で、②はペーペッツの回路と呼ばれる反響回路で、③はCA1から一部が直接的に、他が海馬台およびその近傍を経由して間接的に、嗅内野に至る系である。このCA1から嗅内野への出力路については石崎グループによって主にラットを用いて詳しく調べられており、ネコ、サルなどの所見も加えて考察がなされている(本多と石崎、2001; 石崎、2002、参照)。
海馬のtrisynaptic 神経回路と記憶
上に述べたように、海馬における神経回路をみると、嗅内野から貫通線維を通って歯状回へ、歯状回から苔状線維が伸びてCA3へ、さらにCA3からシェファー線維がCA1へ,そしてCA1から元の嗅内野に戻って閉鎖回路ないし反響回路を作っていることが分かる。このように見ると、海馬内の伝達回路は歯状回→CA3→CA1で表わされCA2を跳び越しているように思われる。この領域はネズミでは狭いが、サルやヒトでは領域的に広くなり、動物の進化につれて発達してくるところらしいが、現在まで詳しい研究の対象にならなかった。改めてこのCA2領域の線維結合を調べてみると興味ある事象が浮かんでくる。CA2から他の領域への遠心路はほとんどなく、同じ領域内の長軸方向の結合が大部分を占めると思われる。求心路の主たるものは上乳頭体(supramammillary nucleus)から入ってくる[Haglund et al., 1984; Ochiishi et al., 1999; Saji et al., 2000]。上乳頭体(およびその近傍)は情動系の入力があるところで[Gonzalo-Ruiz et al., 1992, 1999]、不安、緊張などに関連し[Beck and Fibiger, 1995]、温熱[McKitrick, 2000]や環境変化[Wirtshafter et al., 1998]などのストレスが加わったとき、最初期遺伝子のcFos陽性の細胞が増加することでも知られている。このことを考慮すると上乳頭体・CA2入力系は海馬の機能とくに記憶回路に情動性を与えるシステムに関わっているように思われる。
CA1には重要な「抑制ないし負」の入力があり、また構造上おそらくあるのではないかと考えておりました。理由は少なくとも三つありまして、愚考ですが、①CA3とCA1とをmossy fiber と Shafferで構造上きれいに分けていること。②利根川、中沢らのKO mouse/behavor study から2領域間で記憶障害のパタン/仕組みが異なること。③山口陽子先生の海馬神経回路の記憶モデル(位相歳差)でも記憶の貯蔵と取り出しとに分けようとしているらしいこと。 下記の先生曰くの「海馬内長軸」に興奮が拡がるというお考えはよく理解できない(私の電気生理の知識不足のために)のですが、私は「横軸方向」つまりtranseversally cut section で見てCA3とCA1の間がCA2への[[negative input]によって"septum"が形成されて独立な機能が発揮されると勝手に考えていますが、
この考えはどうでしょうか?私が現在考えているストーリーはこうです。
CA2領域にアデノシンA1受容体が多いことで、海馬の中ではこの領域の興奮閾値が高い。
そのために、海馬内の超軸方向への興奮の拡がりは側抑制のような形で狭くなっている。
その状態の時に、上乳頭体核からの入力がある。そして、歯状回とCA2の活動があがる。
CA2の神経活動があがると、海馬内で長超軸方向に興奮が拡がる。このような形で、上乳頭体核からの不安情報がくると、海馬内でさまざまな情報が連合されるのではないかと言うストーリーです。
コルサコフ症候群の記憶障害と海馬損傷の場合の記憶障害と、臨床的にどう違うのかを、きちんと把握したいのですが、臨床データーを知らないのと、論文を読んでも、どこが違うのかがはっきり把握できていません。この投射様式などを見ると、コルサコフの場合、新規環境におかれたときの不安刺激が海馬にこないために、海馬で新しいことを覚えるのが苦手になるのではないかしらと想像しています。
「ペーペッツの回路」と「ヤコブレフの回路」
情動中枢説が支配的であった1930年代に、情動発現が幾つかの脳部位間の関連活動として捉えられて、情動の「ペーペッツの回路」(海馬→脳弓→乳頭体→視床前核→帯状回後部→海馬傍回→海馬)として提唱された[Papez, 1937]ことは、現在の研究から判断して記憶の回路に属するとは言え、やはり特筆すべきである。現在、情動の回路と言うべきものは、「ヤコブレフの回路」がこれに当たる。すなわち、扁桃体→視床背内側核(MD核)→帯状回前部→海馬傍回→扁桃体をめぐる回路、およびこれと並列して扁桃体→(側頭葉極部)→前頭葉眼窩面皮質→帯状回(その前部→後部)→海馬傍回→扁桃体をめぐる回路がそれで、情動や意欲さらには自我の発現にも関わる神経回路網と考えられている[Yakovlev, 1948]。別に述べる(# §6、§38)ように、中古皮質に属する帯状回、とくにその前部は、意欲に関連した行為を行ったとき局所循環血流量が増加する皮質野として知られている領域で、サルの実験でも独自に新しく工夫した有用な行為を施行して報酬を得た時に活動する神経細胞(ニューロン)が存在する [Shima and Tanji, 1998b] 。
動物実験の結果、新皮質から海馬体への入力/情報の経路は必ず嗅内野・海馬台(ヒト海馬旁回近傍)を介していることが知られている[川村と小野、1987]。これは連合線維(皮質-皮質間結合)の鎖である。固有の海馬ないし海馬体は、発生学的には、「大脳の辺縁(le grand lobe limbique, Broca, 1878)」、すなわち前脳胞から発芽して形成される内外二つのリング状の突起部分の内側の環部が室間孔(モンロー孔)周囲の脳の中心部分を縁どって(limbus)いる所にある。一般には、視床下部を含めた脳幹部から入力される、生存に関わる呼吸、循環、心臓の収縮および食欲、性欲などを包括する自律系・内分泌系からの要素(影響)と皮質連合野からの高度な情報を一時的にでも結びつけることにより、一般記憶の記銘過程や空間記憶の保持に関連した作業に関与していると考えられる。これを支持する行動・生理学的な証拠も提示されている[Eifuku et al.,1995; 永福と小野、2001]。
他方、扁桃体と大脳皮質との関係をみると、扁桃体は側頭葉の前部および下部、前頭葉の腹側部および眼窩面皮質など、感情・情緒に関連する皮質と相互に結合している[Kawamura and Norita, 1980]。その上、扁桃核は味や臭いや自律神経に関連する皮質下の諸核と結合している他、亜核内での役割分担も示唆されており、皮質-扁桃核間の神経回路が働くことによって、賞罰、報酬などの意味づけ、動機づけなどが形成される所と考えられる [Ono and Nishijo, 1999]。このようにみてくると、哺乳動物の大脳辺縁系は大別して、記憶の可変器としての「海馬体系」と、感情表出複合体としての「扁桃体系」とから構成されていると一応みなされよう[川村と小野、1987] 。しかしながら、これらの部位には、量質の差があるにせよ、単に嗅覚系のみならずすべての感覚系の刺激が大脳皮質からも脳幹部からも入ってきており、この両系の調節ないし制御の働きを結びつけ得る場を求めるのであれば、その主要な接点は嗅内野ないし海馬台を含めて海馬旁回近傍にあると考えられる。
クリューバー・ビューシー症候群
扁桃体と情動記憶の関連についても研究が進んでいる。両側の扁桃体が破壊されたサルは、対象物の生物学的意味認知が障害され、食べられないものでも手当たり次第に口に運んだり(精神盲、口唇傾向)、同性に対しても交尾行為を仕掛けたり(性行動の亢進)、以前恐れていたヘビやヒトに平気で近づく(情動反応の低下)ようになる(Klüver-Bucy 症候群、1939)。てんかん治療のために(海馬を破壊する目的の)両側側頭葉切除手術を行った後に、(扁桃体の破壊を伴うために)同様の症候群がヒトでも見られたことが報告されている(1955)。このことは、扁桃体が対象物、あるいは感覚刺激の生物学的意味認知に重要な役割を果たしていることを示唆している。扁桃体は、新奇刺激や感覚刺激の生物学的意味が変化したときに、それらの刺激の生物学的な意味を学習していく過程に関与しているからである。
恐怖条件づけ
ラットに音を聞かせたとき電気ショックを与えるという聴覚条件づけ(恐怖条件づけ)を行なうと、刺激条件(音)だけで血圧上昇やすくみ反応が起こるようになる。扁桃体が破壊されると、この条件づけ反応を用いた課題の学習は障害される。神経生理学的に言って、条件づけ学習により扁桃体ニューロンが条件刺激にも応答するようになるのである。 一般に、強く印象を受けた出来事や情動に裏づけられた出来事に関する記憶が長く残りやすいことは知られている。
情動の記憶
このような情動の記憶増強効果は、扁桃体が記憶固定の過程に促進的に作用していることによる。海馬体苔状線維をテタヌス刺激(100Hz, 1秒の高頻度刺激)すると、記憶・学習に関わると考えられている長期増強(LTP)、すなわちシナプスの伝達効率が歯状回において数日にわたって増強されるなどの可塑的な変化が観察されるが、苔状線維のテタヌス刺激時にさらに扁桃体内側部の電気刺激を同時に加えると、歯状回における長期記憶が増強されることが報告されている[Ono et al.,1993 ; 松本と小野、2002, を参照]。
扁桃体は情動記憶の獲得や固定や保持にも関与している。扁桃体には、感覚条件刺激や強化刺激に誘発された快・不快感あるいは嫌悪性の痛覚情報や報酬性の味覚情報が、(条件づけにより)同期的に入力され、長期記憶などの機序により感覚線維-扁桃体ニューロン間のシナプス結合が強化され、扁桃体ニューロンが条件刺激に応答するようになる。また、扁桃体は、中隔-海馬体系における学習性のシナプス応答の変化を増強させ、扁桃体で得られた情動記憶を長期記憶として大脳皮質に移行させる。なお、臨床的に単極性うつ病患者において[Drevets et al.,1992]、また健康人においても悲しみや喜びなどを表わす情動的な陳述記憶を思い浮かべる場合に、扁桃体で脳血流が増加していることがPETやfMRIを用いた研究により報告されている。
扁桃体への入力系
扁桃体には、味覚、嗅覚、内臓感覚、聴覚、視覚、体性感覚などあらゆる種類の刺激が、皮質感覚野を経過せずに、脳幹レベルから直接的にまたは間脳の視床核を介して間接的に入ってくる。環境からの情報を将にそのまま感覚的に受容する生(なま)の粗な感覚である。その他に、大脳皮質を経由していわば高次元で処理され、知覚され、認知された結果が扁桃体に入ってくるものがある。この後者の入力系はその伝達経路の故に必然的に、時間的に僅かに遅れて伝達されるが、そのことによって、適正かつ精密な情報として入ってくる。すなわち、扁桃体の基底外側核は、梨状前皮質、嗅内野(28野)、帯状回(とくに24野)、側頭葉、前頭前野からの皮質遠心性線維を受ける。以上の粗と精、原始的と識別的、低次と高次という2種の情報が扁桃核内で遭着する。そのことにより、環境に対して瞬間的、反射的に反応した生得的な生体反応は、成体の思慮深い知恵により、快か不快か、有益か有害かの判断に基づいて環境適応的に補正・修正されるのである。なお、ここで言う有益か有害かの判断は実生活上の実利的なものに限定されるものではなく、上記のように、情動記憶と扁桃体の関係からして明らかなように、ヒトの場合、周囲(社会)の情報を処理し、広く動因なるものを選定して、また芸術などに接した場合に知覚される高尚な快の情感、つまり精神的な意味で生体にとって有益な情感の判断もここに含められよう。また、自然の景観や宗教的経験における崇高な感情や超越的感動もここに根ざしていると考えられよう。
動物的情念と高尚な情感
こうした高尚・崇高な情感は、いわゆる哲学や美学に言う精神的なものとしてのみ人間に働きかけてくるのではなく、むしろここにみられる粗と精の合体、すなわち原始的・体感的なものと、たとえば前頭前野を経由する高次なものとの総合によってはじめて認知されるものであると言うのが正しいであろう。何か崇高なものを前にして、人間はいわば心身の全体でこれを受け止めるのであり、またそうであればこそ、崇高にして聖なるものが、遍く(低次な)宗教感情としても、高次精神活動の所産である神学上の対象としても認識され得るのである。神聖なものは、まず第一に高度なものなのではなく、(動物的に)限りなく低く、同時に(人間的に)限りなく高いものであることは扁桃体という、脳の一機能の面からも強調されねばならないし、またこうした低くして高いものの認知が、以下のように視床下部の働きに直接し、それによって体調(これをこそアフェクトと言う)が整えられていく点も注意されなければならない。
分界条(扁桃体の情報を視床下部へ伝える経路)
扁桃体の働きに、たとえば芸術→情動→療法といった、現在社会的関心を集めている音楽療法といったものの経緯の有効性を探る鍵の一つが存在していることは間違いあるまい。ともかくも、ここで行われるこの価値判断の機能の結果は、主として扁桃体の皮質内側核から起こる分界条(stria terminalis)を経由して中隔核、視床下部(前核、腹内側核、弓状核)、内側視索前核などへ伝えられ、小部分は主として基底外側核から起こる腹側扁桃体遠心路(腹側投射系、ventral pathway)により視床下部外側部に伝えられる。
メモ] 視床下部:間脳の一部で、その背側に位置する視床とは視床下溝により境される。その前方は明瞭な境界なしに終脳の一部である視索前野(preoptic area)につながり、後方も格別の境無く中脳視蓋の腹側部と中心灰白質(central gray matter)に移行する。腹側を前方から後方にみると、視神経交叉部(その背側に生体リズムに関連する視交叉上核がある)、正中隆起(内部に下垂体前葉ホルモンの放出因子を産出する弓状核と下垂体門脈系および下垂体後葉ホルモンが運ばれる神経分泌系の視床下部下垂体路が存在する)、乳頭体(海馬から起こる脳弓線維が乳頭体内側核に終始する)が認められる。
視床下部は摂食、性機能、水分代謝などの自律機能や脳下垂体の機能を促進または抑制する内分泌調節などを司る生命維持に関わる重要な領域で、外的または内的な誘因が存在するとき、視床下部への刺激が来たとき周囲への働きかけの動因となり、これによって動物は行動(行為)を起こすことになる。扁桃体から入力される線維束、分界条の形成については相沢らによれば、マウス胎生14~16日で核集団を形成中のニューロンから軸索が束化を形成し、或る特定の細胞群(LOT細胞)をマイルストーンとして尾状核と視床の境界部に沿って背後部へ、また前方および腹方へと走り、形成過程の視床下部領域に侵入する[Aizawa et al.,2004]。そして腹内側部(「満腹中枢」)や外側野(「摂食中枢」)のニューロンとシナプスを作る。これらの領域にあるニューロンはグルコースに反応するが、同時に身体の脂肪細胞から分泌される肥満因子レプチンに対して腹内側部には促進的に、外側野には抑制的に反応する受容体を持つものも存在する(後述参照, # §25)。このほか視床下部には、大脳皮質、海馬、網膜、脳幹などからの線維が入力する。すなわち大脳皮質とくに前頭葉(6野)からは内側乳頭体核や腹内側核へ、海馬からは脳弓を経由して外側乳頭体核へ、網膜からは視交叉上核へ、さらに脳幹からはアミン系の投射が視床下部の広い領域へ行われている。このうち、とくに大脳皮質-視床下部神経路に関しては、ラット、ネコ、サル、ヒトとおそらく動物の発達段階に応じた線維結合の状態が認められると考えるが、今後の研究に期待したい。
嗅覚や味覚の情報も扁桃体が処理する
嗅覚や味覚の刺激は脳幹レベルから直接、扁桃体の系統発生的に古い部分に当たる皮質核・内側核群に入力されており[Norita and Kawamura, 1980] 、その情動への関与も無視できない。水棲動物では嗅覚も味覚も同じ化学物質が各感覚細胞に興奮を与える。これらの動物では、聴覚は水圧や波動の感覚や一部味覚と同様に側線系がこれに当たっている。或る種の魚(ゴンズイ)では、味覚が非常によく発達して側線葉と呼ばれる膨らみが生じており、その前方は小脳原基と考えられる部分で、そこでは聴覚系の蝸牛神経核が発達している。イヌが縄張り設定として嗅覚系を用いて自分の尿の同定をすることはよく知られているところである。
このように、嗅覚や味覚は、とくに下等動物において、辺縁系/情動と強い関連をもっている。が、或る知的な女性から「香りに接したとき、様々なことを思い出したり、せつない、悲しい気持ちなったりする」という話を聞かされたことがある。或る味がその人にとって特定の体験およびさます手段になっていることはよくある。そして胸さわぐこともしばしばある。香道やaromatherapyはもっと高級な話で高次の認知機構に関連するものである。
ここで嗅覚と味覚の結びつきについて考えてみると、この両者には互いに補強しあう関係があるようにみえる。いわゆる「風味」といわれるもので、香りと味の組合せである。しかし、感覚器から大脳皮質までの投射路をみると、味覚系は延髄から孤束核→視床VPM核→外側溝壁部皮質へ、嗅覚は嗅結節→扁桃体、視床MD核→前頭葉眼窩面皮質へと全く独立した別のルートをとる。最近 Dalton らはヒトを対象にした実験で、ヒトにサッカリンのような香りのない甘味物質を口に含ませたとき、塩味や辛味の物質を口に含んだ場合と比べて、サクランボやアーモンドの独特の香りを嗅ぎ分ける能力が向上したと言う[Dalton et al.,2000]。この組合せは経験的に西洋料理によく使用されるが、何れにもせよ小脳の扁桃がこの検知閾値の降下(感度が増すこと)に関係するらしいという。一方、視覚と聴覚との関連については、すでに前の章(# §4、§7)で皮質レベルの形態を中心に論じているが、現象面においても、或る種の音楽を聴いて、情景が視覚的に思い浮かぶことは多くの人達が経験するところである。一般に視・聴覚系は、嗅・味覚系に較べてより高い感覚の階層に属し、原始的な生の情動との関連性は低いと考えられているが、網膜から入力される視覚性刺激を基に視床下部域で生体リズム(サーカディアンリズム)が形成されるという点は注目すべきである。情動と感覚の階層性の問題は別に論ずべきテーマである。
てんかん研究の歴史(アンモン角硬化、キンドリング、遺伝子改変など)
精神医学の分野で海馬はてんかんの際の病理所見、すなわちてんかん発作による血行障害、酸素欠乏、萎縮性硬化症の存在が知られている。すなわち、古典的には海馬のCA1,CA3,CA4に限局する細胞脱落とグリオーシスが強調されてアンモン角硬化(Ammonshornsklerose、海馬硬化)(Spielmeyer, 内村、 1927、により、痙攣の際の機能的血行障害による断血性の細胞変化と脱落をおこし、その結果二次的にグリア線維が増殖し瘢痕が形成され組織が萎縮する)と言われたが、後に扁桃体、海馬傍回などにも病変が及ぶことが知られて、現在は内側側頭葉硬化(mesial temporal sclerosis)と呼ばれている[Burton, 1988; Lambert et al., 2003]。また、燃え上がり現象(キンドリング:kindling phenomenon)[Goddard et al., 1969]が実験てんかんのモデルの一つとして知られている[佐藤、1975]。燃え上がり現象とは、動物(ネコやラット)の海馬や扁桃体にけいれん閾値以下の電気刺激を1日1回数秒間与えると、てんかん発作の準備状態が形成され、低閾値の刺激で興奮が誘発されやすくなるという現象である。この神経機序には関連する神経回路内のシナプス伝達の長期増強(LPT)が関与すると考えられている。この異常興奮の機序としては、シナプス抑制の減少、シナプス興奮の増大(とくに貫通路「perforant path」が終止する歯状回顆粒細胞におけるグルタミン酸シナプス伝達の増強)、さらにニューロンの発芽現象(sprouting)およびシナプス再構成が起こる結果とされている[植村ら、1996]。
注]: 歯状回皮質は分子層、顆粒細胞層、多形細胞層の3層から成る。顆粒細胞層にはグルタミン作動性(興奮性)の顆粒細胞とGABA作動性(抑制性)の籠細胞が存在する。後者は前者を抑制する(フィードバック回路)。顆粒細胞の樹状突起は、分子層内の遠位部で嗅内野や嗅周野から起こる貫通線維を、近位部でCA3とCA4から起こる線維を入力として受ける。また、アンモン角(cornu ammonis)は海馬台側からCA1,CA2,CA3,CA4[Lorente de Nó, 1933-4]の順に領域的に区分される。歯状回顆粒細胞の軸索は苔状線維と呼ばれ、CA3,CA4の細胞に興奮性のシナプスを与える。CA3、CA4の錐体細胞の軸索は、主として海馬後部からは海馬采より出て脳弓線維となって視床下部とくに外側乳頭体核に、また主としてその前部からは外側中隔核に終わる。そしてこれらの軸索はシェファー (Schaffer) の側枝を出してCA1細胞にシナプスを作る。CA1からは線維が出て嗅内野に戻る。海馬への求心性線維としては、貫通線維のほかに内側中隔核や対角帯(Broca)や無名質から入るアセチルコリン性の中隔海馬線維、青斑核からのノルアドレナリン線維および縫線核からのセロトニン線維などがある。アルツハイマー病の海馬や新皮質、嗅脳では、ニコチン性アセチルコリン受容体(nAChR)の数が減少している。因みにアセチルコリン受容体はニコチン性(nicotinic、イオンチャンネル型:α,β,γ,δ,ε)の他にムスカリン性(muscarinic, mAChR、G蛋白質共役型:M1-M5)も新皮質、海馬、線条体、上丘を含む脳全体に広く分布しているが、その発現部位に関して、①どの神経投射系にか、②シナプス前終末かシナプス後膜にか、③血管壁にか等々の量的分布の問題も含めて、今後明らかにされるであろう。従来nAChRは骨格筋や自律神経節に、mAChRは脳や副交感神経支配臓器にあるとされてきたが、近年、中枢神経系内におけるnAChRの存在とその重要性が指摘されるようになった。CA3野から出る錐体細胞軸索の側枝が同じCA3野内の神経細胞にシナプスを形成するという構造を示す、反回性経路(recurrent pathway)がみられるが、このようなネットワークの特徴は記憶の蓄積や想起に貢献する構造の仕組みと考えられている。
LTPという現象は、てんかん発症(epileptogenesis)のメカニズムの解明に関連があると思われるが、これまでは記憶や学習との関連で多くの研究がなされてきた。本格的な臨床研究が始まったのは、てんかん治療のために両側海馬切除手術をうけたH.M.という名前の患者が、てんかんの発作は治まったが、予想外に、あるlife event をきっかけにその後の記憶を失うという現象である「前向性健忘症」が発見さて以降のことである[Scoville and Milner, 1957]。
最近は海馬に発現する酵素などの特定の遺伝子を人工的に破壊して、働かないようにして作製した遺伝子に異常のある「ノックアウト」マウスを用いた研究が行われている。たとえば、利根川のグループは、領域特異的に、海馬CA1野(Tsien et al., 1996)およびCA3野(Nakazawa et al., 2002, 2003)においてNMDA受容体(この場合、その活性に必須であるNR1サブユニット)を、またLTPに重要な役割を演じていると考えられるα-CaMKII(α-Caカルモジュリン・タンパク質リン酸化酵素II:この阻害剤を添加するとLTPの発現がブロックされる) を、Cre-loxP法を用いてノックアウトし、シナプスレベルでのLTPの発現機序と行動レベルでの記憶・学習の関連について調べ、その相関を明らかにした。もっとも、LTPとキンドリングはともに電気刺激に対して作られる長期間持続する可塑的変化であるという点では同じではあるが、LTPが反復刺激の短いバースト(burst、群発射)を与えることで作れるのに対して、キンドリングの方は数日から数週かけて毎日繰り返し刺激を与えなければ成立しないという現象である。その点からして、両者は現象としては似てはいるけれども同じ機序として説明することは難しい。実際、この両つの現象の誘導過程において、歯状回の顆粒細胞でc-fos, c-jun, jun-Bなど最初期遺伝子群のmRNAの発現を比較して可塑的変化について調べてみると類似性はあるものの、確かな相違が認められる(Cole et al., 1989)。キンドリング現象は扁桃体に反復電気刺激を与えることによっても成立する(Uno and Ozawa, 1983)。
扁桃体は嗅内野でシナプスを介して歯状回と線維連絡があり(# 前述、本節§21)、神経系の可塑的な変化の故に、キンドリングの形成過程で相互に二次的なてんかん原性(epileptogenicity)を獲得する可能性は十分に考えられる。てんかんの他に、最近は、統合失調症、うつ病、外傷後ストレス障害(post-traumatic stress disorder , PTSD)の患者の脳でも、海馬や扁桃体の機能異常や萎縮、神経細胞の脱落などの形態変化が報告されている。善悪、好悪、是非、賞罰の識別や情緒・気分状態の健全な切り替え、さらには自律神経・内分泌系が深く関与する大脳辺縁系の快楽や恐怖への感知システムに対して、如何に適切に前頭葉皮質の制御機構が働いているのか、現在、治療法の開発と結びついたこれらの研究が進んでいる。それにつけても、大脳辺縁系-視床下部-前頭前野を中心に大脳皮質連合野を基盤とする神経回路の発達障害の原因究明については今後の研究に俟たねばならないが、これらの精神疾患の易再燃性(cf. 臺の履歴現象)について考えるとき、人間の精神のはたらきを深いところで支えている、辺縁系が演じる根源的な役割にあらためて気付かされる。