要約:慶應義塾大学解剖学教室 川村光毅
| 嗅覚受容器 | |
| いくつかの比較解剖学的データ | |
| 嗅球 | |
| 嗅神経入力と糸球層の構造 | |
| 嗅球の結合 | |
| 前嗅核 | |
| 嗅結節 | |
| 扁桃核 | |
| 梨状葉皮質 |
| 第10章.嗅覚路、扁桃体、海馬、扁桃系 要約:慶應義塾大学解剖学教室 川村光毅
|

文明人(教養人、センスのある人間)の生活にとり嗅覚はさほど重要でないということで、臨床上嗅覚は系の重要性は少ない。よりたくみなテスト法が用いられたならとくに、嗅覚検査から一定の診断情報は得られる。故に嗅覚結合の解剖の基本的特徴となる知識は神経学者にとって有益である。哺乳類に於いては、扁桃核の一部だけが嗅覚に関係し、海馬は恐らく全くかかる関連はないが実際的目的から、両者ともここで考察しよう。同様に、中隔といわゆる“辺縁系”も簡単に検討されよう。
| 嗅覚受容器 |
嗅上皮は、鼻腔内で、鼻中隔の後上部と外側壁の相対する部分にある。限局は領域で一部上鼻甲介をためると所にある。この嗅粒膜部は黄味を帯びている。嗅上皮の細長い感覚細胞は支持細胞より結合されている。この感覚毛は“嗅覚棒(杵)、olfactory rod”として、感覚細胞の自由表面からの突出部にある。6~20本の細い毛で被覆している粘液mucusに突出す繊毛(嗅小毛)ciliaの構造を示している。支持細胞には微絨毛microvilliがある。各嗅細胞の基底部から1本の細い糸状の突起がありこれが一定方向にインパルスを伝える。この形態的特徴は、嗅覚上皮は脊椎動物間における感覚細胞の内で原始型primitive typeであることを示し、嗅覚がすべての感覚中で系統発生的にみて最も古く原始的なもの、であるとい見解を支持する。嗅覚細胞の無髄近位繊維は軸索に相当するcomparable(くらべる)ものである。
嗅覚受容細胞間にある程度の組織学上の区別がある。しかし形態学上から特定のタイプにわけることは光顕上(LeGros, Clark, 1956)も電顕上(de Lorenzo, 1963)も成功していない。以下の本を参照。Olfaction and Taste (1975) Taste and Smell in Vertebrates (1970), Douek (1974) monograph。
非常に細い嗅覚上皮からの繊維(径ca 0.2 μm)は合して細い束となり篩骨ethmoid boneの篩板cribriform plateを貫き脳内に入り、嗅球に達する。これらの嗅覚繊維束の全体が第1脳神経すなわち脳神経である(という)。
嗅上皮:多列線毛上皮で3種のcells
| いくつかの比較解剖学的データ |
魚類と両棲類では、終脳の主体は嗅覚インパルスを外套部pallial partに(運ぶ)伝える求心性繊維であり、例えば哺乳類にみられるような皮質様構造の特徴を片鱗も示してない。
比較解剖学的研究から、両棲類に見られる3つに分けられる外套の部分は、高等脊椎動物の外套部palliumに認められる特定部に相当すると推論される。すなわち
内側部→archicortex or海馬の原基
外側部→paleocortex or梨状(葉)皮質
中央部→neocortex = “dorsal area”新皮質の背側皮質
爬虫類では、皮質構造はなお原始型であるが3区分はもっとはっきりしてくる。
哺乳類になりはじめて、背側皮質dorsal cortexが著明に発達する。
この発達の間(発育の経過中)、嗅覚性インパルスの分布paleocortexとarchicortexに限局されるようになる。ここは全体として下等哺乳類で高等の分化のレベルに達したのちは、高等哺乳類で更に発達するこはない。
Archicortexにはヒダが形成されfolded、海馬溝の発達により、海馬として側脳室の内側壁の中に大きくふくれる(突出する)。新皮質の発達の結果、paleocortex, archicortexは遂に内方に押しやられ、ヒトでは10-1,4図にみられるように、全く半球の内側面にみられる。新皮質の発育は他領域の形の変化の原因ともなる。半球皮質の後頭極occipital paleが発達すると、paleocortexとarchicortexは後方に引かれ、遂に、その後、側頭葉の発達にともない再び前方、腹方に引かれる。このためにヒトや多くの哺乳類のarchicortexとpaleocortexの部分は、環状に近いnearly circular構造を示し、室間孔前方域から後方に拡がり、ついで下方にそして前方に曲がり、遂に主発点の下方にあたる脳底に達する。模式的には、archicortexは、主として海馬と歯状回によって代表されるが、これは、この線lineの全長にわたってみられる筈である。しかしながら、脳梁が両半球新皮質間の非常は発達によって現れてくると、この位置にあるarchicortexの部分は著しく縮小する。その部分に相当するものはinduseum griseum脳梁灰白質と一般に考えられている。脳梁膝の下にある脳梁下回(gyrus subcallosus, BNA,JNA,: PNAではGyrus paraterminalis終板旁回)は(いく人かの著者-研究者によって)海馬の最前部と見倣されている。(cp. 10-4図、p.650)。Paleopalliumは哺乳類で後方に向かって発達し、梨状葉を形成するが、ヒトでは海馬回(Gyrus hippocampi, JNA, BNA; PNAではGyrus parahippocampalis, 海馬旁回)の大部分がこれにあたり、その(梨状葉,paleocortex?)残りは帯状回および、帯状回と海馬回(PNAで海馬旁回)を結ぶretrosplenial cortex膨大後部皮質の形で存在すると考えられている。しかしながら、多くの研究者は、帯状皮質は実際はpaleocortexとneocortexの移行皮質と考えている(“辺縁系”の後述をみよ)。paleocortexとarchicortexには種々の細胞構築的区分域あり、この時点で、海馬回(PNAで海馬旁回)内のブロードマン28野 、entorhinal area内嗅野について注意をひくことで充分である。
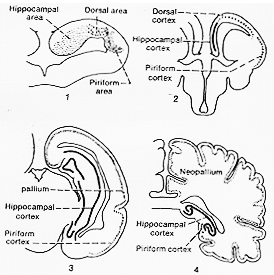
FIG. 10-1 Diagram (after Herrick, 1933) to illustrate the development of the olfactory parts of the brain. In amphibians (drawing 1) a dorsal area is found between the hippocampal area (archipallium) and the piriform area (palaeopallim). In reptiles the dorsal area has the appearance of a cortex (drawing 2); in mammals this develops progressively as the neocortex or neopallim (drawing 2 from opossm) and in man (drawing 4) entirely overshadows the piriform cortex and the hippocampal (archipallial) cortex. Compare text.
上述の記載は甚だ模式図的であるが、系統発生上のこれら構造物の発達の印象を与えるとは役立つだろう。長いカーブをえがく脳弓や分界条stria terminalisのような哺乳類やヒトでみられる2・3の特殊な(奇妙な)繊維結合の像(特徴)を説明する。とりわけ、比較解剖学により、下等脊椎動物では全半球を占める脳の部分(paleocortex archicortex)がいかにして、哺乳類になって、新皮質によって全く被いかくされるようになるかを明らかにしてくれる。下等動物において第一に嗅覚に関係すると見倣される脳のpaleocortexとarchicortexの部分は、哺乳類そしてとくに後頭哺乳類おいて他の重要な機能にとって代わった。
嗅脳rhinencephalonという術語はしばしば使用されるが、不幸なことにいつも同じ意味で用いられている訳ではない。系統発生学的研究を基礎にして、嗅脳は通常外套部pallialのarchicortexとpaleocortexを中隔野、嗅結節、嗅索、嗅球など終脳基底域の部分を一括とする。しかしながら、この術語は生理学的意味を言外に含んでいる。繊維接合分析の結果として、いわゆる嗅脳の多くの部分が、その名称が意味するように特に嗅覚に関係していると考えることはむづかしい(できぬ。not entitled)ことを明らかにしている。このような術語を用いることは不幸である。以下white Jr.の1965b引用。
| 嗅球 |
嗅上皮からの繊維は1対の嗅球に終わる。嗅球に終わる。嗅球は脳の一部でもともと終脳からevaginate(外転、ほんてん、turn inside out, event)した部分である。篩骨篩板上の頭蓋骨面に位置する嗅球から、嗅覚インパルスを中枢に伝える繊維結合が始まる。これらの結合は完全にはまだ判ってない。以下に主たる点のみを記述する。主たる点のみを記述する。嗅球構造はすべての脊椎動物で似たもの。(Allison, 1953bをみよ)。大まかに云うと、嗅感覚細胞からの求心繊維は2次ニューロンである僧帽細胞(mitral)と房飾細胞(tufted)の樹状突起といわゆる糸状体glomeruli内で結合interlockしている。僧帽細胞はたくさんの可成り粗な2次樹状突起をもち、軸索を嗅索に送る。房飾細胞も多少これに似ており、同様に多くの樹状突起を有し、それらのあるものは糸球体に送る。且、少なくとも一部は中枢に投射(axon)するようである。これには疑問(論議)ある所だが、最近Haberly と Price (1977)はHRP法で房飾細胞の軸索がrostral terminal stations of the olfactory tract fibersに達することを確かめた。又、僧帽細胞も房飾細胞も正中を超えて前交連crossして軸索を送ることはないようだ。Golgi標本を基礎に房飾細胞の軸索のあるものは嗅脳内に終わるintrinsicものと主張さる(Valverde, 1965をみよ)。最後にいわゆる外叢状層external plexiform layerより下(深部)に顆粒細胞granule cellsがあり、この細胞は樹状突起を外叢状層に送り、ここで僧帽細胞や房飾細胞の長い樹状突起とdendro-dendriticのシナプスを作る散在性のいわゆるperiglomerular cells (PB細胞)が糸状体の付近にみえる。
嗅受容細胞の軸索が糸状体中の樹状突起状に作るシナプスはasymmetricタイプで丸い小胞をもち、一般に興奮性を考えらる。
僧帽細胞(M)と顆粒細胞間(G)にみられ、dendrodendriticシナプスは相互いにpresynaptic。
M→G(E) (asymmetricタイプ、球状小胞)
G→M(I)(symmetric扁平小胞)
顆粒細胞の樹状突起には棘が密、顆粒細胞は古典的意味での軸索を欠くようだ(網膜上のアマクリン細胞と比較)そして、それ故、そのactionを樹状突起のみで他の細胞に及ぼす。
顆粒細胞は、嗅球に繊維(後出)を送る中枢の構造物からelictされ嗅球のactivity活動の抑制に必須の如し、CNSのある一般原則の公式化formulationを促進 局所回路とその構成ノルアドレナリン終末は顆粒層に存在する如し、以下2頁(X-5, X-6)嗅球の回路(森憲作)-脳の統御機能3“感覚と知覚”より
図1は(嗅球)ちょうど地球が核を中心とする幾層かの層状構造によって構成されているのと同様に、嗅球は顆粒は顆粒細胞層を中心にして5して層がほぼ楕円体系にとりかこんで構成されている。その最外層は嗅神経繊維層(olfactory nerve layer; ONL)で、嗅上皮中の嗅細胞から投射されてきた軸索(嗅神経)からなる層である。嗅神経はさらに、すぐ下層の糸球層(glomerular layer; GL)にある糸球とよばれる神経叢の中に侵入して、ここで終末を形成する。この糸球には、嗅球の出力細胞である僧帽細胞tufted cellの主樹状突起の先端部が細かく分枝して分布しており、嗅神経からシナプスを受ける。糸球のまわりには多数の小型の細胞(periglomerular cell; PG細胞)が存在する。この細胞は樹状突起を糸球内に送っており、またその軸索は糸球層付近に限局して分布する。糸球層から約500 μm深部に僧帽細胞層(mitral cell layer; MCL)があり、ここに僧帽細胞の細胞体がほぼ1列にならんでいる。僧帽細胞は通常1本の主樹状突起と数本の副樹状突起をもっており、主樹状突起は糸球層まで分枝することはなく伸びて、糸球の中で細かく分枝するが:副樹状突起は糸球層と僧帽細胞層とにはさまれた外叢状層(external plexiform layer; EPL)に限局して存在する。僧帽細胞層より深部に顆粒細胞層(granule cell layer; GCL)がある。ここには多数の小さな細胞(顆粒細胞)が密につまって、多くの集落を形成している。顆粒細胞は無軸索細胞であり、peripheral processとよばれる樹状突起を外叢状層に、そしてdeep dendriteとよばれる樹状突起を顆粒細胞層深部に向かって出している。嗅球には上記のニューロンの他に、少数ながら幾種類かの短軸索細胞がまばらに存在する。
| 嗅神経入力と糸球層の構造 |
Allisonらによると、ウサギの一側の嗅球に投射する嗅神経の数は約5千万本と推定されている。これに対して、嗅球には約1,900個の糸球が存在する。嗅神経は嗅球に入るまでほとんど分枝しないことから、1つの糸球は約2万5千本の嗅神経入力をうけることになる。また、嗅球には約4万5千個の僧帽細胞と13万個のtufted cellがあり、それぞれ1本の主樹状突起を糸球に送っている。したがって、1つの糸球には約24個の僧帽細胞と約68個のtufted cellが関与していることになる。さらに、糸球のまわりには数百のPG細胞が存在し、樹状突起を糸球内にだしている。
この嗅覚系の最初のシナプス中継部位である糸球内の神経叢の解析はPinchingとPowellやWhiteらによって電子顕微鏡学的になされてきている。彼らの研究によると、嗅神経は糸球内で僧帽細胞、tufted cellおよびPG細胞の樹状突起上にシナプスを形成する。さらに、糸球の中には嗅球中のニューロンどうしの樹状突起間シナプスも多数存在する。
たとえば、僧帽細胞樹状突起からPG細胞の樹状突起に向かって、球形のシナプス小胞と非対称形のシナプス膜肥厚をもった(おそらく抑制性だと思われる)シナプスが、またPG細胞の樹状突起から僧帽細胞樹状突起に向かっては、扁平型のシナプス小胞と対称形のシナプス膜肥厚をもった(おそらく抑止性だと思われる)シナプスが存在することが報告されている。さらに、これらのシナプスは相反型シナプスもしくは直列型シナプスの形で見い出されてきており、糸球内部での複雑なシナプス相互作用が推測される。この糸球層でのシナプス相互作用、特にPG細胞の働きを電気生理学的に解析しようとする試みは、GetchellとShepherdらによってなされてきたが、まだ十分とはいえず、今後の進展が期待される。
| 嗅球の結合 |
一次嗅覚繊維→嗅球…他の脳の部分へ嗅覚インパルスを伝える経路の出発点。(嗅球への主たる求心繊維で感覚細胞の中枢性突起) 他の求心路については後述する。
嗅球から僧帽細胞と房飾細胞の軸索が出。嗅索olfactory tractを作る(10-3,10-4図)。後方に扁平に延び、外側及び内側嗅条としてやや(比較的に)分離されてある。両嗅条が分かれる所に3角領が嗅三角olfactory trigone, trigonum olfactoriumで、この嗅条からの繊維はいくつかの核に行く繊維を構成する。多くの方法(Marchi, 鍍銀法、Nauta, FH, EM, HRP)で多くの動物で調べられた。動物差に程度の差あり。オポスム、ラット、モルモット、ウサギ、ネコ、ツパイ、サル。
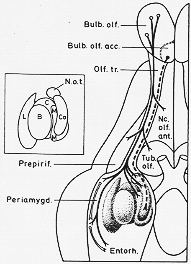
FIG. 10-3 A diagram showing the course and sites of termination of efferent fibers from the main olfactory bulb (uninterrupted line) and from the accessory olfactory bulb (interrupted line). Projections from the periamygdaloid cortex to the entorhinal area (white) are also shown. The inset diagram above is a key to the identification of subdivisions of the amygdaloid complex (see Fig. 10-5). N.o.t.: nucleus of lateral olfactory trafct. Note: olfactory fibers terminate in this and the cortical (and medial) amygdaloid nucleus. Compare text. From Lammers (1972)
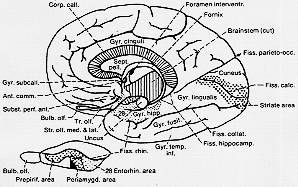
FIG. 10-4 A drawing of the medial aspect of a right hman hemisphere, showing some of the olfactory structures. The primary olfactory cortex, indicated by coarse stippling (like the striate are 17), covers parts of two small gyri which appear to corespond to the prepiriform and periamygdaloid areas (see text). An extension of the primary olfactory cortex into the anterior part of the entorhinal area (fine stippling) as determined in animals, is not indicated. Lower left, a diagram of the basal surface of the rabbit's brain for comparison. The primary olfactory area is indicated according to Rose (1935), and symbols correspond to those in the drawing of the human brain.
ARG法でもラット、ウサギ、ツパイ、サルで行われた。
小さな相違を除き一般に、哺乳類での嗅球からの繊維の終止部位は以下の如し、
前嗅核anterior olfactory nucleus(嗅索に散在する細胞Niimi)
嗅結節olfactory tubercle
扁桃核の一部の亜核amygdaloid nucleus
梨状葉皮質piriform lobe
中隔核
視床下部
以下にこれらの終止について記する(10-3図)。
| 前嗅核anterior olfactory nucleus |
嗅索のbase(底部)にあり、嗅球より繊維をうける。対側のパートナー(前嗅索)に投射。他に遠心繊維が嗅球へ、及び、嗅三角、前梨状皮質、視床下部へ少し(Brodwell, 1975b)。前嗅索は嗅球から求心繊維をうける部位に、一般的に云って、投射するように思われる。そして、これは嗅球からの直接の経路を補足する意味の、一定の領域への嗅覚インパルスに対する間接ルートの中継核である如し。生理総説としてDaval と Levetau (1974)をみよ。
| 嗅結節olfactory tubercle |
明からに(恐らく)前有孔室(ヒトの形態で)に相当し、嗅三角のすぐ後部に位置する。嗅結節は嗅球と前嗅核から繊維をうける。これはmicrosmatic動物において可成り大きく。anosmaticのクジラでさえ可成り大きい。サルでは、嗅核求心繊維は少く。しかし、側頭葉から可成り繊維をうける(20,21,35野)。霊長類では、嗅結節は嗅いと関係が薄く。‘二次的’嗅核領野とはほとんど考えられない如し。
| 扁桃核amygdaloid nucleus |
扁桃核への嗅球からの繊維は、皮質核と内側核に限局される。うち、ARG法により、皮質核が主であることを示した。中心核へもあるらしい。嗅覚繊維が、視床下部とくにそのVM核(ventromedial核)に投射する扁桃亜核に主として終わることは機能学的観点から興味深い(11章をみよ)。かくして嗅覚受容器と視床下部の“食餌(摂食)中枢、feeding center”との間に比較的直接的なシナプスの少ないoligosynapticルートがあるよである。しかしながら、嗅球から直接の視床下部hypothalamusへの投射もある。とくに、その前方域と外側域(Powell, Cowan, Raisman, 1963, 他)嗅覚刺激、生殖および他の種々の行動パタンへの影響。
| 梨状葉皮質piriform lobe |
梨状葉皮質へもrather consistentに投射あり、paleocortexに属する。種間の差もあり。梨状葉の細胞構築上の亜核分類は一致しない。しかし通常的には、吻側から尾側へ3つの主たる領野に分かれる(Valverde, 1965)。
prepiriform area梨状前野
piriform (periamygdalid扁桃周野) uncusの上前面を占める。
entorhinal (Brodmann’s 28)ヒトで最大。…海馬旁回の広い前方部を占める。
以前は梨状前野、梨上野(周扁桃体野)より後方域には感覚繊維は行かぬと思われていたが、内嗅野(Brodmann’s 28)にも終わる。ことが近年の方法で判った。内嗅野は海馬の主たる求心入力の起始部である。
嗅球から海馬固有部への繊維は確証無し。
第一次嗅覚皮質はどう考えるべきか(他の感覚系に対応して)?多くの研究は、嗅球と前嗅覚から直接の繊維がところと限る。故に(こう考えると)prepiriform areaとperiamygdaloid areaと内嗅野の可成りの外側部。
この一次嗅覚皮質はある種の哺乳類(ウサギなど)では比較的大きいが、ヒトでは10-4図に示されているように海馬旁回の前端と鈎に接する小域を占めるにすぎぬ。視覚、聴覚、味覚、体性知覚の第一次知覚領野とは対照的にこの一次嗅覚野はallocortex(12章参照)内にある。この皮質は、他の一次知覚野に特徴的な典型的な顆粒状の外観を欠いている。又、他の面でも異なっている。すなわち、視床内の中継核の後、脳の内部(深部)からこの皮質に達するのではなく。表面から達する。(cp)最近のTakagi groupの仕事→新皮質-眼窩(面)前頭皮質にある。
惟うに、ヒトの一次嗅覚領は、第一に、嗅覚刺激の意識的な認知に関与するらしい。他の終止域は、おそらく嗅覚刺激反応で引き起こされた反射活動と行動反応に主に関与している。しかしながら、一次嗅覚皮質と扁桃核からの二次結合も考慮されねばならぬ(後をみよ)。嗅球へは体側嗅球からも繊維が入る。HRP法でprepiriform cortex “horizontal nucleus of the diagonal band” nucleus of the olfactory tract and the olfactory tubercle. lateral hypothalamus cortical amygdaloid nucleusからのbulbusへのafferentsを証明。このcentrifugal fibersは糸球体まで行く。顆粒細胞とasymmetricalシナプスをつくる。抑制的(Kerr と Hagbarth, 1955)。交連繊維前交連を通る。嗅球間、しかし、少なくとも、これらの大部分の繊維は前嗅覚anterior olfactory nucleusからの対側の嗅球へ。前交連は他の繊維も含む(後出)。[d.h. 左右の嗅球(古皮質)を結ぶ。高等Aでは新皮質の一部も加わる。扁桃体を結ぶ物も少数あり。](by Niimi)
最終更新日:2002/09/13
