第12章. 大脳皮質
The Cerebral Cortex第12章. 大脳皮質 The Cerebral Cortex |

大脳皮質は前脳胞の吻側部に由来する終脳より発達し形成されるが、構造的にも機能的にも、独立して存在しない。例えば、嗅球と直接あるいは間接的に線維結合する領域もあり、このような系統発生学的に古い領域をおおざっぱに「嗅脳rhinencephalon」と呼ぶ。より下等な脊椎動物では、この領域がかなりの部分を占める(第10章)のに対し、哺乳類ではその他の皮質領域が著しく発達する。ここで出現する大脳半球は「新皮質neocortex or neopallium」と呼ばれる。そして新皮質は高等哺乳類やヒトでは大きさを増し、その表面積は体積に比べて増大する。
大脳皮質の構造: 細胞構築と領野の局在 Structure of the cerebral cortex. Cytoarchitecture and areal parcellation. |
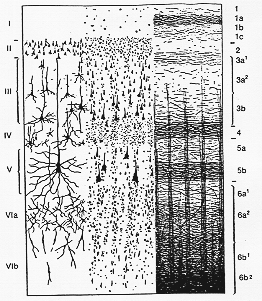
FIG. 12-1 Diagram of the structure of the cerebral cortex. To the left, form a Golgi preparation; center, from a Nissl preparation; to the right, from a myelin sheath preparation. I: lamina zonalis; II: lamina granularis externa; III: lamina pyramidalis; IV: lamina granularis interna; V: lamina ganglionaris; VI: lamina multiformis. After Brodmann and O. Vogt
新皮質はどの領域でも同様の発達を遂げることから、しばしば「等皮質
isocortex (O. Vogt) or homogenetic cortex (Brodmann)」と呼ばれる。領域毎の差はあるものの(後述)、一般的に6つの層からなる(図12-1)。Ⅰ 分子層は軟膜直下にあり、線維が豊富だが細胞には乏しい。小型の神経細胞が散在し、
Cajal細胞は皮質の表層に水平に向いている。Ⅱ 外顆粒層は錐体型・丸形・星形をした小型の細胞から成る。
Ⅲ 錐体層は主に中程度の大きさの錐体細胞から成る。この層の深部では、より巨大なものも見られる。
Ⅳ 内顆粒層は小型の細胞が密に集合している。その多くは星形をしているが、錐体型をしているものも多い。この層では、水平向きの線維が豊富に存在する(外バイヤルジェ
Baillarger線)。Ⅴ 神経節層
The ganglionic layerは主に中型ないしは大型の錐体細胞から成る。他の錐体層と同様、分子層に向けて長い尖端樹状突起を伸ばし、水平に走る基底樹状突起を出す。Ⅵ 多形細胞層は主として紡錘型の細胞を含む。しばしば外側のⅥ
a層とⅥb層に分けられるが後者は次第に白質に移行する。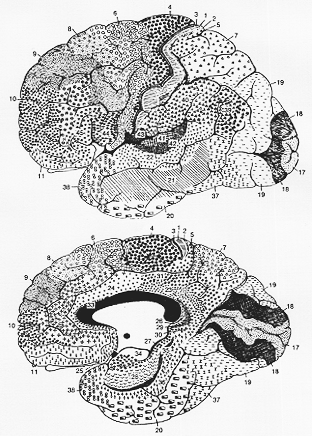
FIG.12-2 Brodmann's cytoarchitectural map of the human brain. The various areas are labeled with different symbols and their numbers indicated by figures.
一方より詳細に観察すると、多くの小さな部位毎に明確な差が認められることが
Campbell (1905)・ Brodmann (1909)・von Economo (1927)らによって認識された。これらの細胞構築の違いに基づきBrodmannは皮質を50の領野に、またvon Economoはさらにその2倍の数の領野に分割した。図12-2はBrodmannによるヒト大脳皮質の細胞構築別の地図を示す(von Economoも同様の地図を作製している)。この図によると、各領野の間の境界は大脳溝の位置と一致しない。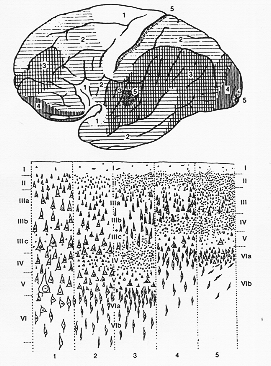
FIG. 12-3 The different principal types of cortex (below) and their distribution in the hemisphere (above). 1: agranular cortex; 2: frontal type of cortex; 5: granular cortex, koniocortex (see text). After von Economo from Kornmuller and Janzen (1939)
細胞構築が共通する領野を元に、
von Economoは5つのグループに大別した(図12-3)。下図の2・3・4型は典型的な皮質の6層を持つのに対し、その他の型(1・5型)は各層の発達が同程度でない。前者はhomotypicalなのに対し、後者はheterotypicalであり、皮質が十分に発達しても6層が明確に識別できない(しかし、Brodmannによると胎生期は6層が形成される。それ故、これらも等皮質に属する)。図中で基本型1は顆粒細胞層(第2・4層)が欠落しているのに対し第3・5層はよく発達していることで区別できる。この
agranular cortexは中心溝よりも前方から前頭葉後部端で見られる。第4野と第6野のほか、第8野・第44野がこれに属する。これと同じ型の皮質構造がparacentral lobeの大脳半球内側面・帯状回の前半部・終板よりも前方・海馬ならびに鈎の一部にも観察される。agranular cortexの特殊型は第4野で見られ、ここでは第5層のより大型の錐体細胞の間にBetzの巨大錐体細胞がある(第4章参照)。この領野からは、遠心性の投射系が圧倒的に多く出る。(図8-7a参照)。それ故、agranular cortexは運動野皮質のプロトタイプであると考えられている。大脳皮質では血管供給の領域差もあることが、
Pfeifer (1940)によって示された。毛細血管の密度・その配列・口径その他の血管構築が大脳皮質の層や部位によっても異なり、この領域差は、多くの点で大脳皮質における領野の局在に関する結果と一致する。これと対応するように、グリア細胞の分布や配列は領野ごとに異なるが、その詳細は完全には明らかでない。近年の研究の中には、大脳皮質の化学構築について扱ったものがある。様々な酵素の化学構成や量に関して、領域ならびに層の間で差があることが明らかとなっている。このような研究は動物でもヒトでもなされ、そこから大脳皮質の病態(例えば老人性痴呆症、Pope, Hess, and Lewin, 1964)に関する情報を得ようという試みがなされてきた。この分野は将来が期待されるが、まだ始まったばかりである。ヒト大脳皮質における神経細胞の数は膨大で、
14×109個(von Economo and Koskinas, 1925)と見積もられている。それに対し、Pakkenberg (1966)は若い成人の脳全体で2.6×109(26億)個と報告した。このような研究者による数の違いは、組織学的手続きによるものと考えられるが、ヒト脳の個人差も要因となり得るようだ。
| The fiber connections of the cerebral cortex. |
大脳皮質の線維結合には、4種のものがある。すなわち、(1)皮質遠心性の投射線維、(2)皮質求心性の線維、(3)同側半球の異なる領域を結ぶ連合線維、(4)左右半球を結ぶ交連線維である。
(1)については、錐体路が広範に調べられており、皮質脊髄路や皮質延髄路は多くの皮質領野から起こるが、その大部分は中心前・後回にある(第4章)。また皮質橋路もヒトではよく発達している(第5章参照)。かなり広範囲な皮質領域から尾状核や被核への
topicalに配列した投射、赤核や毛様体への遠心線維については、第4―6章を参照。その他の遠心線維は後柱核dorsal column nucleus(第2章参照)・下オリーブ(第5章参照)・subthalamic nucleus(第4章参照)へも、わずかながら投射する。また、とりわけ後頭葉からの遠心線維は上丘や視蓋へと走行する(第7章参照)。最後に、皮質視床線維が豊富にある。これらの大部分は、視床からの求心線維を受ける皮質の領野から出て、視床の核へと走行する。皮質の異なる領野の活動は、大脳皮質に多量に存在する連合線維によって起こると考えられる。その中には非常に短く、灰白質内を通り抜けて隣接する領野と相互に結合するものもある(各機能領野の短連合線維については、第2・4・8・9章を参照)。その他の線維は幾分か長く、主に白質内を走行する。長連合線維のあるものは同側大脳半球の領野を結び、またあるものは正中を交叉し両半球の領野を結ぶ。
| Long cortical intrahemispheric and interhemispheric (commissural) association connections and some notes on the cortical “association” areas. |
大脳半球間を結ぶ連合線維はよく交連線維と呼ばれ、その大部分は脳梁である。これに対し、半球内の(同側の)長連合線維、特に大脳皮質のいわゆる連合野どうしを結ぶことを中心に考察する。初期の研究(
Polyak, 1927b; Mettler, 1935; Jansen, 1937その他)より、一般には機能的に密接な相関関係を持つ領野同士は連合線維によって最も密接に結合すると考えられたが、このことは後の研究でも確かめられた。近年は、サル(一部はネコ)にHRPを用い、連合線維の起始細胞について多くの情報が得られるようになった。サルとネコとの間には種差がいくらか存在するようで(Kawamura, 1973c参照)、長連合線維はサルの方がネコよりも発達しているようだ。一方ヒトでは連合線維はもっと重要であると信じられているにもかかわらず、有用な情報はほとんどない。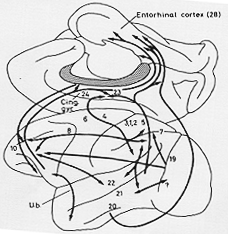
FIG. 12-4 A simplified sketch of some of the long association fiber connections of the "association areas" of the cerebral cortex, and their connections with parts of the allocortex, plotted on diagrams of the monkeys's brain. Only some main features are shown. Fibers from the association areas to area 6 are not included (see text). Arrows point to sites of termination of fibers. Note that most association connections are reciprocally organizaed. (The fibers passing from the orbital surface of the frontal lobe to the entorhinal area are shown as passing in the cingulum bundle for the sake of clarity of the illustration, while they have been found to pass directly and to take off from the uncinate bundle (U.b.) between the orbitofrontal cortex and the temporal pole.) Numbers refer to apporoximate positions of some of Brodmann's areas.
長半球内連合線維
long intrahemispheric association fibersは一様に起こるにも関わらず、かなり複雑なパターンで正確に構築される。こうしてある連合線維束内では、特異的な投射パターンが通常存在するのである(例:側頭葉―前頭葉間投射)(Jacobson and Trojanowski, 1977a)。上側頭溝により隔てられるそれぞれの部位は、その入力のパターンが異なり(Seltzer and Pandya, 1978)、また前頭葉でも同様に、連合線維のパターンが領野によって違う。また、皮質の連合野間の線維結合は大部分が相互に行われている(例:前頭葉―側頭葉間ならびに側頭葉―前頭葉間結合)(図12-4)。不等皮質との連合線維結合も存在する。長線維は大脳皮質の異なる「連合野」と特に相互結合するようにみえるのに対し、これらの領域からの遠心性線維は、より特異的に結合する(とりわけ前運動野である第6野)。これは「peristriate belt」(Pandya and Kuypers, 1969)・頭頂葉の第5野(Jones and Powell, 1970a)・聴覚野皮質(Diamond, Jones, and Powell, 1968b)から長連合線維を受けることが発見されている。
大脳皮質内部の構成とシナプス結合 Intrinsic organization and synaptology of the cerebral cortex. |
かなり初期から、感覚機能と顆粒細胞層の発達、また運動機能と錐体細胞層(特に第5層)の間にある種の相関関係があるに違いないことが結論づけられていた。この見解は、まもなく支持され、大脳皮質の外層すなわち第Ⅰ―Ⅳ層は主として受容・連合の機能を持つのに対し、深層の第5・6層は効果器としての機能を担うという考えが一般に受け入れられた。しかしこの知見は概要に過ぎず、軸索や樹状突起の層構築まで含めたより詳細なことは何も分かっていなかった。そこに挑戦したのが
Lorente de No (1949)で、ゴルジ法を用いて体系的にCajalの仕事を拡張していった。軸索の分布について、
Lorente de Noは大脳皮質の神経細胞を4つの基本型に分類した(図12-6参照)。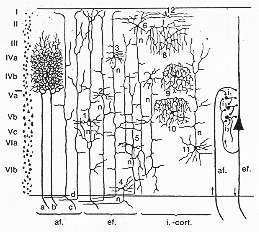
FIG. 12-6 A very simplified diagram of different types of neurons in the cereberal cortex, with axons, dendrites, and collaterals. af: axons of afferent fibers; a,b, and c: fibers from the thalamus; d: association fibers; ef: neurons sending their axons out of the cortex, from the cell labeled 1 as a projection fiber, from cells 2-5 as association fibers; i.-cort: cells whose axons arborize intracortically, those of cells 2-5 as association fibers; i. -cort: cells whose axons arborize intracortically, those of cells 6 and 7 in the deeper layers, of cells 8 to 10 mainly in their own layer, cell 11 with its axon to more superficial layers, and celll 12 with a horizontal course of the axon (see text). To the right a simplified diagram of the passage of impulses within the cortex. af. and ef.: afferent and efferent fibers; i1, i2, and i3 : intercalated neurons. redwawn and simplified from diagrams of Lorente de No (1949).
①白質に向かって軸索を伸ばし、投射線維として深部の構造や皮質の他の領域に続く(図12―6の1-5)。
②唯一短い軸索を持ち、これが細胞体にすぐ近接して分布する(図
12―6の8-10)。③軸索を表層に伸ばし、軸索やその側枝が皮質の一つあるいはそれ以上の層にわたって分布する。(図12―6の11)。
④軸索を水平に伸ばす神経細胞(図
12―6の12)。大脳皮質細胞への神経インパルスは視床の細胞の軸索や側枝に由来するほか、一部は同側の皮質の他の部位にある細胞(連合線維)または反対側(交連線維)からで、加えてする隣接する皮質の細胞からも来ている。
図
12―6でLorente de Noによると、視床からの求心線維(一番左)はその大部分が感覚路の一部を構成しながら、内顆粒層(第Ⅳ層)に終末の分枝を広げて終わる(図12―6a、bの線維)。彼が「特異的求心線維specific afferent fibers」と呼んだように、この線維群は皮質の深層を通過する際、側枝を全く出さない。その他の視床皮質線維は、図中の線維cによって示されたように、分子層(第Ⅰ層)に至るまで、幾つかの層にわたり側枝を送る。このような型の求心線維は「非特異的nonspecific」(第2章参照)と呼ばれる。同様にして、他の皮質の領野からの連合線維は全層を通過するが、その側枝の大部分は皮質表層部とりわけ第Ⅱ・Ⅲ層の細胞に終わるようだ。軸索を灰白質から送る細胞の中には、錐体路のように、離れた神経核群に向かって伸長する線維を出すものもある。このタイプの細胞は特に第Ⅴ層に存在する(図
12―6の1)。その他の細胞からの軸索は同側半球の他の皮質の領野に向けて経路を取る連合線維か、又は反対側の大脳半球に向かう交連線維である(図12―6の2-5)。このタイプの細胞はとりわけ第Ⅲ・Ⅴ・Ⅵ層で見られる。後者のタイプの細胞では、軸索が皮質内を走行する途中で多くの側枝を伸ばすため、軸索が皮質深層を通過する際に神経細胞に対して影響を与えることができる。第Ⅴ層の神経線維そうは、ここからの遠心性軸索の側枝がかなりの部分をなす。皮質深層にある細胞の中には、より表層に向かい反回側枝を送るものもある(図12―6、細胞5)。それ故、第Ⅴ層ならびにⅥ層の「effector」細胞はより表層の細胞に対して影響を与えることができるのであろう。大脳皮質の全層にわたり、軸索や側枝が皮質内にのみ分布する細胞がある。ゴルジタイプⅡ細胞が特に豊富で、これらは紡錘細胞と呼ばれる(図
12―6の8-10)。その軸索は核周囲部に密接して広く枝分かれする。軸索が表層に向かって伸び、1層ないしは複数の層にわたって分布する細胞も、全層でみられる。このことは、皮質深層にある細胞が作動するよう仕向けられると、インパルスを表層に送ることができることを示唆する。水平な軸索を持つ皮質内のニューロンは、第Ⅰ層で有意に存在する(図12―6の12)。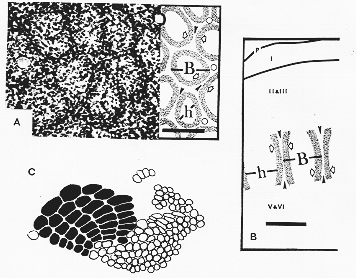
FIG. 12-7 Illustrations of some main features of the anatomy of the cortical barrels in the face region of the primary sensory cortex in the mouse. A: A photomicrograph of a tangential section of cortical layer IV in the anterior barrel field. The explanatory diagram shows that each barrel (B) is made up of a ring of densely lying cells (stippled)p, which surrounds a less cellular area, the hollow (h). Septa (black arrowheads) separate the barrels. Bar=100μm. B: A diagram of the somewhat larger barrels in the posteromedial barrel field, represening th emystacial vibrissae, as seen from the lateral aspect (constructed on the basis of coronal sections). C: A camera lucida drawing of the barrel field of a mouse cerebral hemisphere. The barrels (black) in th eposteromedial barrel field, related to the mystacial vibrissae are arranged in five distinct rows and are larger than the others. For descriptions see pp. 516-517. From Woolsey and Van der Loos (1970).
幾つかの動物において、一次感覚野皮質の顔面領域認められる第Ⅳ層のバレル
barrelsは、コラム状の機能的構築の代表例である(図12―7)。各mystecial vibrissaが一つのバレルに投射することが明らかにされている(第7章参照)が、このバレル構造が果たして縦方向の機能的コラム構造に相当するのかは、不明である。大脳皮質の主要な細胞のタイプである錐体細胞と紡錘細胞の区別は明確でないようだ。前者の中には、皮質から出ない軸索を持つものもあり(
Valverde, 1976; Tombol, 1978)。紡錘細胞あるいは顆粒細胞(ゴルジⅡ型細胞の不均一な細胞群)はいくつかの異なる種類がある。この分類は研究者によって異なっているが、これは両タイプ間を移行するものが存在するためであろう。紡錘細胞の一般的な特徴は、少なくともその大部分が介在ニューロンであり、その軸索が皮質から出ないことのようである。第3のタイプである籠細胞basket cellはCajalによって最初に示され、主に水平方向にのびる軸索を持ち、その側枝は終末ボタンをもち垂直方向に伸びているのが特徴である。その側枝はより深層に位置する錐体細胞の細胞体を取り囲み、それが籠のように見える。また樹状突起は水平並びに垂直方向に走行する。籠細胞のうち最大のものは、第Ⅲ・Ⅳ・Ⅴ層にみられる。これらの細胞はヒトの運動野皮質にみられ、とりわけ第Ⅳ層に多い(Marin-Padilla, 1969)。籠細胞は明らかにいくつかの錐体細胞上で作用するに違いない。
| The cerebral cortex in phylogenesis. |
系統発生学的に高次になるほど、皮質内の神経の伝達もより精密になっていく。とりわけ連合野の細胞の比率が著しく上昇し、ヒトではこれらの細胞が大脳皮質全体の大部分を占める。哺乳類の大脳皮質では、細胞構築と髄鞘形成において分化の程度が上昇している。連合野として機能すると考えられる部位は、ヒトでは特に進歩的な発達を示しており、それはとりわけ頭頂葉や側頭葉で広いようである。それに対し投射線維を送るあるいは受ける領域は増加せず、大脳表面積全体でも比較的少ない部分を占めるに過ぎない。
| Functions of the cerebral cortex. | |
| The frontal lobe. |
運動性失語
motor aphasiaは、優位半球の前頭葉(顔面運動野のすぐ前にある領野)の損傷で起こったと考えられる言語障害である。1861年にブローカBrocaによって初めて報告されたことから、この部位はブローカ野Broca’s areaと呼ばれる。この言語障害は、自分の考えを言葉で表現できなくなる。しかし、発声に関係する顔面筋・舌筋・喉頭筋などは障害されていない。また運動性失語の中には、指の機能が障害されておらず正常に機能するのに書くことができない(失書)ものもある。運動性失語は、言語機能の障害のなかでも唯一、脳の損傷で観察される(後述)。「
Prefrontal」 granular cortexは片側の前頭葉の切除ではほとんど症状が出ないが、両側性に切除することで特徴的な症状が現れることがかなり早くから分かっている。
| Cerebral dominance. |
「大脳半球の優位性」という言葉は、大脳半球のいずれか一方がある機能に関して主として働く半球のことを言うのに用いられ、その差は複雑な言語機能に関して最も顕著である。「
leading hemisphere」という概念は右利きの人で運動性失語が頻繁に起こることから、1869年にHughlings Jacksonによって初めて示された(Zangwill, 1960参照)。左利きのヒトでは状況が逆になるとよく言われる(右半球の損傷で、失語症になりうる)がこれは必ずしも成り立たない。左利きの人の大部分が、言語機能に関しては左半球が優位であることが明らかとなった。根拠の蓄積とともに、人では右半球よりも左半球の片側が優位であるという概念はさびれ、両半球が相補的に細分化しているという概念にとって替わるに違いない。つまり左半球はある機能に関しては優位あるいは「先導的leading」で、右半球はその他の機能とりわけ「空間機能」に対しては優位なようである。言語機能に関しては、大脳の半球優位性は遺伝的に決定されるようである(
Geshwind, 1974b参照;以下の脳の解剖学的不均衡性についての記述も)。しかし、利き手は少なくとも一部は環境によって決定されるようである。多かれ少なかれ、両手利きの人は少なくない。これは、脳の半側にのみ損傷を受けた左利きの患者で言語障害の発生を調べたZwangwillの研究(1960)からで、PenfieldとRoberts (1959)なども同様の結論を導いている。
| The cerebral cortex and language functions. |
言語機能の障害は、もし明らかに存在するならば、優位半球の障害によって起こることが言われている。このような障害は失語症
aphasiaと呼ばれ、その症状は話し言葉や書き言葉の理解は保持されるのに、発話において生じる障害(運動性失語motor aphasia)と、主に言語の受容(見る・聞く)機能の障害(感覚性失語sensory aphasia)との区別が重要としてなされた。これらの事例では、理解の障害が表現の困難を引き起こすため、それぞれ表現性失語expressive aphasia、受容性失語receptive aphasiaとも呼ばれる。しかし一般には失語症の患者を詳細に調べても、純粋なタイプの失語に出会うことは決してないと言われる。言語機能の障害は単に損傷の部位や程度のみで決まるものではなく、生前のその人のパーソナリティー・知的水準・知識量などによっても左右される。そのため、WeisenburgとMcBride (1935)が述べたように、失語症には個人差があり、全ての失語症を説明できるような仮説は存在しない。側頭葉てんかんを持つ患者を対象とした言語中枢の電気刺激による実験によって、
Penfieldらは多くの情報を提供してきた。それらを要約すると、側頭葉なかでも上側頭回は知覚(主に視覚や聴覚)した対象を再生recallしたり、聴覚あるいは視覚による信号を解釈したりするのに特別な役割を果たすと結論される。そのため、この領野はPenfieldによって「interpretive cortex」と呼ばれ、言語機能に関与すると考えられる領野とは別物であることが強調された。
| The parietal lobe. |
側頭葉の病変による症状には、感覚症状(第2章)ともう一つ、体の反対側半身の筋萎縮(上肢で著しい)がある。これには中心前回が付随的に関与するからであるとよく説明される。側頭葉病変による筋萎縮の十分な説明がこれまで与えられていないが、その出現についてはある程度診断に利用できる。
側頭葉障害の中には
agnosias(認識の欠如)と呼ばれるものがあり、多くのタイプが存在する。これらに共通した特徴としては、感覚刺激の認識に障害を持つことである。体の反対側半分に与えた刺激や反対側半分の視野を無視する(または気づかない)。一般に、欠損は空間関係の認識を必要とする課題で特に著しい。頭頂葉の機能を調べる課題はサルよりもヒトの方が明らかに複雑であり、このことを示すのがヒト脳における機能の側方化
lateralizationである。しかし、以前ほどには優位半球と非優位半球の区別は明確でなくなり、むしろ両半球特に左右の頭頂葉の間の協調が、機能を発揮するのに非常に重要であることを示唆する。このことはさらに、大脳皮質の交連線維の重要性を示す近年の研究からも支持されている。
| Functions of the corpus callosum. |
系統発生的に、哺乳類に於いて新皮質の大きさが増大することにより、脳梁の大きさも並行して増加している。脳梁の機能は両大脳半球が協調的に機能するのを助けることであり、ヒトに於いてはこの機能が特に重要であると仮説されてきたが、これについてはほとんど分かっていない。脳梁の発達が十分でない(脳梁の形成不全)人間についての研究がある。このような事例では、他の発達障害はあるものの、神経学的な症状はたいてい、全く表出しない(
Ettlinger, Blakemore, Milner, Wilson, 1974ならびに近年の研究を参照)。てんかん発作の拡大を防ぐために行われる脳梁の切断では、患者は目立った行動の異常を示さない(Smith, Akelaitis, 1942; Brigman, Smith, 1945などを参照)。このような患者では、脳梁がヒト脳の機能にとりわけ重要であるとはほとんどいえない。しかし現在では、このようなネガティブな実験結果の理由として、脳梁の機能を評価するのに相応しいテストが利用されていなかったことが明らかとなっている。近年では、ヒトで少なくとも脳梁切断後に観察された変化の大部分は、後部1/3の線維を切断したためであることが示されている。
Grodon, Bogen, Sperry (1971)は、交連線維の切断範囲が前交連や脳梁前部に限局すると、情報が両半球の間でなお転移することを発見した。
| Evaluation of clinical changes in lesion of the cerebral cortex. |
脳にできた腫瘍を外科的に切除すると、その一部は左半球にあり、このことが幾つかの症状を引き起こしている可能性がある。そのうえ腫瘍の繁殖は頻繁に起こり、切除による損傷は皮質下の構造に影響を与える。
| Cortical localization. |
例えば運動や体性感覚といった領野を明確に区別できないように、古典的な、明瞭な機能局在という概念は支持され得ない。しかし、特定の領野は特定の機能と関係しない一方、ある皮質の領野は多かれ少なかれ幾つかの機能と密接に関係する実際、近年の研究では、視覚機能に於いて第
17・18・19野は同一でなく、それぞれ異なる機能を持つことが明らかとなっている。
最終更新日:2002/09/13
