| 第2章.体性(知覚)の求心路 |
要約:慶應義塾大学解剖学教室 川村光毅
・皮膚受容器の機能的分類
・関節の感覚Joint sensitivity or kinesthesia (運動[感]覚)
・関節の神経支配
・求心性の知覚性線維
・後根および前根の中の求心線維
・脊髄の構成の2・3の特徴、後根線維の終止
・分節的な知覚支配、The
dermatomes (皮膚)
・分節的な知覚支配、The
dermatomes (皮節)
・後索と内側毛帯の繊維
・脊髄視床路
・Spinocervicothalamic tract脊髄・脛・視床路
・視床と視床皮質路
・内側毛帯と脊髄視床路の視床における終止

種々の知覚様態
一般体性求心系(cp. 特殊性の体性求心系-目、耳から)exteroceptive, proprioceptive
受容器
皮膚受容器の形態
free or unencapsulatedの神経終末
encepsulatedの終末
Merkelのdisc(自由終末の多少特殊化されたタイプ)
Encepsulated:
Pacinian corpuscles (2-1A図)
Corpuscle of Meissner (2-1A図)
Corpuscle of Ruffini (2-1A図)
Krause and bulbs
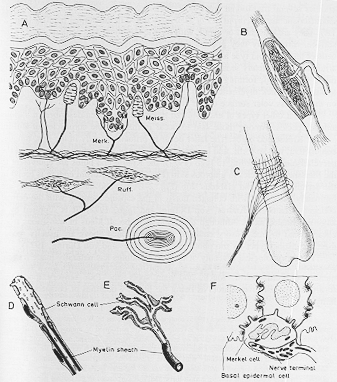
FIG. 2-1 Diagrammatic drawings of various types of cutaneous receptors. For description, see text. A: Typical distribution of receptors in hairless skin. Ruffini endings are found in the dermis, while the Pacinian corpuscles occur subcutaneously B: Drawing of a Ruffini ending (redrawn from Bannister, 1976). The capsule has been opened, and the nerve fibers can be seen to course between the collagen fibrils. C: Thke arrangement of myelinated nerve fibers around a hair root. D,E: Drawings of free nerve terminals, based on electron micreographs (redwawn from Andres and von During, 1973). The nerve fiber is enclosed by Schwann cells except at small regions near its end. At such places the axoplasm shows an increased granularity ("receptor matrix"). F: Dwawing based on an electron micrograph of a Merkel's disc (redrawn from lggo and Muir, 1969).
| 皮膚受容器の機能的分類 |
Mechanoreceptors
Meissner
Merkel
Ruffini
われわれが普通tactile stimuliと呼んでいるのに4-5のreceptoraタイプが関連している。
Thermoreceptors
構造上温度受容器と同定された特別のものはない。が被覆されたものではないことは確からしい。冷覚受容器は、細い有髄線維の自由終末(生理とEMの仕事)(Hensel,’73)
Pain receptors or Nociceptors cp. Noceo害を加える。
fine, freely ending fibers
low intensityで神経が直接刺激うけたとき、太い有髄求心線維のみがactivateされて、触覚様感覚tactilelike sensationが起こるが痛み(-)。電気刺激の強度を増していくとthinnest myelinated線維もactivatedされ痛みを感じるようになる。 *(C-group, C-fibers)
- TorebojorkやHallin 1973-1974の仕事。-
皮膚の痛みに関して主観的に2つのタイプ
pricking, fast, first, pain細い有髄繊維 (group Aδ)
stinging, slow, second, pain細い無髄線維
他に、痛覚刺激に対する神経繊維の感度に2つの主要なタイプがあるようだ(Burgss と Perl, ’73; Lym, ’77)
Pacinian corpuscleファーテル・パチニー層板小体
Vater - Pacini’ sches Lamellen Korperehen
皮下組織など。いくつかの点で皮膚受容器に入れられるよう。単一神経繊維により支配、大きい。 圧→他の種類の刺激にも対応。esp. 振動vibration
高頻度振動>100サイクル/秒
| 関節の感覚joint sensitivity or kinesthesia (運動[感]覚) |
運動・重量、位置などを関知する感覚、普通はとくに、関節の角度の変化の知覚に関して用いられる。
muscle spindle
Golgi tendon organs
筋紡錘からの情報は筋の長さによるものばかりでなく。fusimotor activityのレベルにも依るので、筋紡錘は関節の絶対位置を記録することはできない。
| 関節の神経支配 “articular neurology” |
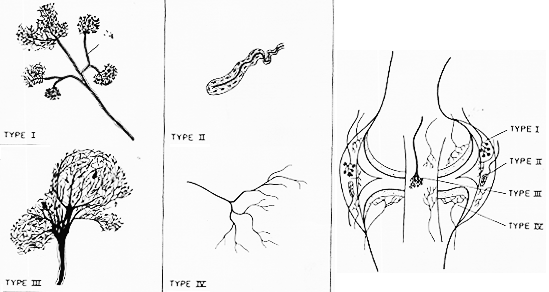
FIG. 2-2 Above, semischematic illustration of the four joint receptors usually distinguished (based on Skoglund, Polaced, 1956; Polacek, 1966; Freeman and Wyke, 1964). For description see text. To the left, a diagram of the knee joint, showing the distribution of the various receptor types in the ccapsule and ligaments of the joint. the menisci are free from nerve fibers except at their attachment to the fibrous capsule.
e.g. FreemanとWyke (’67) 関節の受容器終末を形態的に4つのタイプに分けた。図2-2.各々のタイプは各々特殊の機能的性質をもっている如くであるがすべてtension receptorである。 (張力、緊張、伸張、伸展)受容器
Head’s theory of protopathic (primtive) and epicritic sensibilityベッドの理論。(Rivers と Head, 1908; Head, 1920) 反論があるが(Walsche, 1942aの批判的総論をみよ)。なお支持者が多い。
受容器からのインパルス脊髄神経筋内にsomaをもつ求心性知覚線維により中枢へ伝えられる。末梢神経中の線維の太さは非常に細い無髄線維から太い有髄線維までは幅広い。太い線維は大きい脊髄神経節細胞に属し小→小cell。一般に鈍皮膚神経とその枝は運動神経や混合神経の幹よりも細い無髄線維を豊富に含んでいる。
ex. ヒト尺骨 神経深い所 無髄線維/有髄線維 0.69/1
表在枝 無髄線維/有髄線維 2.12/1
Pacini小体関節受容器のいくつかMeissner, 筋紡錘、腱器質・・・を支配するものは比較的太く有髄
表皮や真皮のsubepidermal層あるものは大部分非常に細い
somatic afferent体性求心線維は、前述の如く、皮膚、筋、腱(粘膜)etcからインパスルを後根から脊髄に伝える。そしてその偽単極性の細胞体を脊髄神経筋内にもつ。加えに、多数の無髄の求心線維が前根から入ってくるという可成りの証拠が現在ある。この問題は後で考察されよう。顔面からの求心線維は三叉神経を通じて入り、細胞体は半月神経節内にある。後根を構成する線維の厚(太)さは大変に幅があり、径が20 μmにも至る太い有髄鞘のものから、2 μmにも至らない細い無髄線維まである。後根の全線維数は相応する前根中の線維に比して可成り大きい。(ヒトについてはRexed 1944あり)。
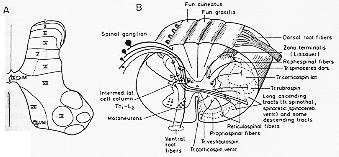
FIG. 2-3 A: Diagram of a transverse section through the 6th cervical ssegment of the spinal cord of the cat, illustrating the cytoarchitectonic subdivision of the gray matter into approximately horizontally arranged laminae (I to X) according to Rexed (1954). LM and VM: lateromedial and ventromedial groups of motoneurons. B: Schematic drawing of an isolated segment of the spinal cord, showing the klocation of some cell groups and the main ascending tracts. Not all tracts are included, and the borders between adjacent tracts are not sharp. a: Neuron in Claarke's column, giving origin to the dorsal spinocerebellar tract; b: neurons giving off spinoreticular and spinothalamic fibers.
後根中の線維は一連の細(長)い束としてdorsolateral sulcusで脊髄に入ってくる。ここで通例に従いこれらの線維を2つの群に分けよう。第1の小部分の外側群:細い線維タイプで大部分が無髄線維。脊髄に入ってすぐに、2分し上行性線維と下行性線維となり両者共、側枝を腹側に与えて分布し、後角に到り、そこで分枝を出しすぐに終始する。大部分の上行性線維も下行性線維も脊髄の1~2 segment以上は走行せず。しかし、あるものは前方3-6分節、後方4-5分節達するものもある。最も細い線維のうちあるものは、zona terminalis, tract of Lissauer, or後外索に含まれる(2-3B図)。リッソウェル氏辺縁帯他の第2群は大きい太い線維の内側群で大部分が有髄。
この内側群線維は、後角のすぐ内側部で白質に入り、外側群の線維同様に、白質中で2分し上、下行する。上行線維の若干は非常に長く前方に延髄の尾部まで至り後索核、gracilis, cuneatusに終わる。
後索核 薄束核(med) Goll
楔状束核(let) Burdach
この長い走行径過中側枝はほんの僅かしか出さない。それも主に始まりの部分の内側群線維の他の線維はほんのみじかい上行性と下行性の枝を出し、たくさんの側枝をもっている。これらの線維は後角の灰白質内に入り、種々の群に分かれて神経細胞とシナプス結合をもつ。
今や前根から求心線維が脊髄に入ることは確立されたと思われる。以前から示唆はされていたが、たくさんの動物でヒトを含んで、無髄線維が驚くほどの高率前根に見られることがEMで分かった。ヒトでは前根中の軸索の28%(平均して)がunmyelinated。明らかに、ネコでは、この50%が自律系の節前線維である(Applebaumら、1976)。しかし脊髄根の色々のレベルでこの比率には可成りの幅がある。前根中の無髄線維の多くが求心線維であるという証拠は色々の実験から明らかである。例えば、前根を切り、近位で変性がおこる(Walker d.)が遠位では(-)。より決定的な証拠は神経節より近位で後根を切った後HRPを脊髄内に注入すると脊髄神経節内にlabelされた細胞が見い出された(Maynard et al ’77, しかしYamamotoら’77をみよ)注入前に前根も切ってしまうと“殆ど陽性細胞はみられない”。生理実験は少ない。
| 脊髄の構成の2・3の特徴、後根線維の終止 |
横断面でみた脊髄の灰白質の構造はどこでも一様である訳ではない。いくつかの細胞群は可成りはっきりしている。たとえば前角の多極性細胞群や(Th1-L2)で後角内側部にみられるClarke柱はそうであるが、他方、他の群は輪郭がはっきりしないものもある。厚いNissl標本を用いて細胞構築をBaseにしてRexed (1952, 1954)はネコ脊髄灰白質が10ヶのlargely horizontal,帯または層に分けられることを示した(2-3A図)。機能的知見から3群に分けると。
afferent terminal fieldをみると特定の層に多く終わるが(層の機能分化を思わせる)介在ニューロンの突起の拡がりを見ると多くの層にわたり複雑な構成をしている(故に特定の層)
多くのintersegmental fibersは灰白質の近くに見られ、massive fasciculi proprii (sometimes called propriospinal fibers)と呼ばれる(Fig. 2-3)。この線維は前索、側索、後索にもみられる。特に背外(側)索dorsolateral fasciculus (Lissauer路)をnotedしておかねばならぬが、大多数の細い線維が含まれintrinsicで1-2分節を走る。ネコではLissauerの繊維の75%がこのタイプ(Earle, 1952, Szentagothai 1964aもみよ)。しかしLa Motte (1977)は猿の脛髄をARG法で調べ、Lissauerの50%の繊維は一次求心性繊維であると結論づけている。ヒトの脊髄固有繊維は、NathanとSmith (1959; La Motte 1977も)より詳しく調べられている。
隣接脊髄分節間結合に加えるに右と左のpossible cooperationを作るに正中を横切る。軸索をもつ細胞がある。VIII層にある細胞に多い。これらは腹側および背側の交連を通る。
変性研究で太い後根求心繊維中の運動ニューロン間に終止するもの -一次性筋紡錘求心繊維と同定(Iles, 1976; BrownとFyffe, 1978)。
後根繊維のうち灰白質を通過してIX層の大型前角細胞に終わるものがある(図2-3でみるように)。又、後根繊維の側枝がクラーク柱の細胞にContactしているのも変性実験でみられている。他の後根繊維とその側枝は後角と中間帯に終わる。
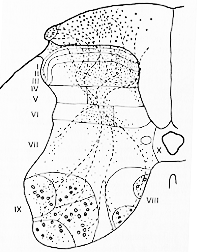
FIG. 2-4 Drawing from the 6th lumbar segment in the spinal cord of the cat to show the sites of termination of degenerating fibers 5 days following transection of the ipsilateral root of L5 as seen in Nauta preparation. Degenerating fibers in the dorsal column are indicated by coarse dots, those in the tract of Lissauer by intermediatesized dots. Small dots indicate sites of termination. Fibers of passage are shown as dashed lines. The laminae of Rexed are indicated (see text). From Sprague and Ha (1964).
Spragueら(1958, 1964)によれば腰髄レベルで全層に終止が見られる(図2-4)、しかしとくにIV層とVI層(VI層では主に中央部)に多い。
少数の後根繊維は正中をこえて対側の後角に終わることが昔から知られていた(Schimert, 1939: SpragueとHa, 1964)。最近の解剖・生理共同研究によれば、これらの繊維は皮膚内のnociceptors由来でI層V層に終わる(LightとPwerl,1979a,b)。この章の後半部で求められるが。これらの層にある多くの細胞は対側脊髄視床路にaxonを送る。故に、(二重交叉するので)このcrossing dorsal root fibersは求心性入力に対して同側に上行する“痛覚路”に貢献すると考えられる。
求心性繊維の後柱膠様質(Rexedの2層)への終止。確実に実験的に証明された。この所見の解釈は様々だが、動物により分布領と密度に差が見られる。
Brown, RoseとSnow (‘77b)単一軸索HRP注入法により
flame-shaped arbors →主としてIII層に終わる。僅少がII層。
LightとPerl (’79a,b). HRP法(後根に注入)
細い有髄繊維→I層、II層の内側(Inner)部。III層
無髄繊維→主としてII層の外側(outer)部
太い有髄繊維→III層に終わる(I,II層には終わらぬ)
膠様質substantia gelatinosaの機能的役割については時代により意見があった。知覚(感覚)情報の過程で特別の役割を演ずる如くであるが以前に考えられていたように、“痛覚路”における最初のシナプス部位ではない。最近ますますはっきりしてきたが、後根から脊髄に入ってsensory informationは、後角ないで可成りの修飾をうけ、この修飾motificationは異なる条件下で変わるものである。Golgi, Ag法、EM法(研究者略)によれば、膠様質の構成の主要な特徴は以下の如く要約される。すなわち、
HRP法によれば少数のII層とIII層にある細胞はラットでlateral cervical核や下部脳幹に軸索を送る(Gieslerら’78)。又、サルでは視床までも、この後者のタイプは感覚(知覚)伝導に直接関与しているだろうけれども、残りの膠様質にある細胞が、脳幹や視床に投射している細胞の活動を修飾しているexact wayはナゾとして残っている(remains enigmatic)。
The gate control説 (MelzackとWall, ’65)
後根入力繊維にII層の細胞がpresynaptic結合し、抑制してclose, or open・・・少し誤りがある。
結論を出すにはデータ不十分。(II層のメカニズムを理解する上で)
前にふれたように、生理学的研究は、後角の層間に機能上の差異が存在するという見解に対して証拠を与えた。Wall (1967)は、皮膚と関節に刺激を与えた後、IV-VI層でunitsを研究し、これら3つの層におけるunitsは皮膚刺激に反応するが、VIでのunitsのみ運動movementにsensitiveである。更に、IV内には内外に配列する体性局在性パターンがあるようである(Wall, ’67; BrownとFuchs, ’75)。しかし、小さい受容野をもっている細胞は、毛の動き、皮膚の触覚、冷覚刺激のようなdifferent皮膚刺激に応答する。V層の細胞はIV層の細胞の軸索によってactivateされる。V層の細胞は異なる様態の収斂する入力converging inputsをうけ、多くは痛みの刺激によりactivateされる。特に興味を引くのは、多くの細胞は内臓性と体性からのcoverging inputをうけている。生理と解剖の所見と一致する。しかしながら、情報は、むしろ複雑でどの層も1つの特定の感覚“様態”に関連しているということはできない。多くの細胞が痛覚刺激にのみactivateされることから、このrule規則の例外にほぼ近づくと云えようか(ChristensenとPerl: 70; 他)。
| 分節的な知覚支配、the dermatomes (皮膚) |
somite体節、重複あり、個体差あり、触・痛・温-性格に同一のデマトームでない。触に関しては、温・痛に関してよりもデルマトームはより広い。
前にもふれたように、生理学的研究は、広角の層間に機能上の差違が存在するという見解に対して証拠を与えた。Wall (1967)は、皮膚と関節に刺激を与えたのち、IV-VI層でunitsを研究し、これら3つの層におけるunitsは皮膚刺激に反応するが、VIでのunitsのみ運動movementにsensitiveである。更に、IV内には内外に配列する体性局在パタンがあるようである(Wall, ’67; Brown と Fuchs, ’75)。しかし、小さい受容野をもっている細胞は、毛の動き、皮膚の触覚、冷覚刺激のようなdifferent皮膚刺激に応答する。V層の細胞は、IV層の細胞の軸索によってactivateされる。V層の細胞は異なる様態の収斂する入力converging inputsをうけ、多くは痛みの刺激によりactivateされる。特に興味を引くのは、多くの細胞は内臓性と体性からのconverging inputをうけている。生理と解剖の所見と一致する。しかしながら、情況はむしろ複雑でどの層も1つの特定の感覚“様態”に関しているということはできない。多くの細胞が痛覚刺激にのみactivateされることから、このrule規則の例外ほぼ近いといえようか(ChristensenとPerl, ’70: 他)。
| 分節的な知覚支配、the dermatomes (皮節) |
Somite体節、重複あり、個体差あり、触・痛・温。-性格に同一デルマトームでない。
触に関しては、温・痛・に関してよりもデルマトームはより広い。
延髄にある後索核に上行して終わる太い有髄の後根繊維は、後根繊維の内側群の可成りの部分を占める。
頸髄上部でみると最内側部(後索の)は仙髄からのもの(内に押される)次に腰部の後根(下肢)から。上肢からのものは楔状束の外側部後角に近い所に。このようにsomatopicallyあり、後索核終止も然り、single-unit recordingsでも確かめられた。しかし上行するとつれて多少の繊維のre-sortingはみられる。
後索中の繊維はその他にあり、上にみるよりももっとheterogeneousである。多くの一次求心性の繊維が脊髄レベルに終わる(propriospinal fibers)。この他に後角にニューロンをもつ第2次ニューロンの多くのaxonsが後索(柱)を上行し後束核に終わるものもある。更に後索核に(→IV層に起始細胞のものが脛・腰レベルで多い(Pustioni, 1976サル)”spinal border cell”の前角の最外部にも少しある。)起こる下行繊維もこのdorsal column中にあることも知って(→V層に終わる)多く必要あり(BurtonとLoewy, 1977)。
主として臨床経験から、後索を上行するインパルスは、触覚、深部圧覚、振動覚、関節の位置覚をmediate伝えるものでありこれらのインパルスは知覚を識別するのに特に重要であるというのが古典的見解であった。しかしよくみると後索のみの傷害というのは稀で他の部位も貫かれ少なかれ傷害されており、CookとBrowder (1965)の報告例-後索切載術例-では上述の感覚に殆ど永久的な傷害lossはみられぬほど反証がある。完全な説明とはほど遠いが、studyで後索は比較的(むしろ)複雑な識別的taskを行うに必要な感覚信号を伝えると云えよう。文字を皮膚の上に書いてみて簡単にテストする(後索の傷害を)。-皮膚上の動きの方法をdetectする能わず。また後索は関節の感覚や振動覚にとっては決定的に重要なものではないように思われる。
後索覚:細胞構築的に区別あり、主に2つの(細胞タイプ)(サル・ネコ)
後索核Single unit study. “modality-specific” (Perlら,1962, etc.)
特定の種類のreceptorからの刺激にのみ反応する。
触覚の受容野は小”place-specificity”
この空間の厳確性は皮膚受容器からsupplyされる細胞のlateral inhibition (inhibition of the “surround type”)によります。
前にのべたように後索核は細胞構築上も求心繊維分布上も同質性でない。遠心性結合についてもこれはあてはまるようだ。とは云っても種間の相違があり一般化することは難しいが。
視床に投射するニューロンは大型で主に”cluster zone” にある(Kuypers と Tuerk, 1964; Berkley, 1975その他)。サル後索核では、しかしながら、視床投射の比率にはこのような明らかな差は見られない。Rustioniら(1979)によれば、HRP法、thalamus注入後:前方”reticular” zoneで
ネコではHRP(+)細胞:HRP(-)が1:1
サルではHRP(+)細胞:HRP(-)が5:1
“cluster zone”への投射とは異なり、”reticular zone”への投射はサルとネコでは同様の求心繊維を受けていることを考えると、内側毛帯経由視床へ送られる情報はネコよりもサルでは多種(orより変化に富むmore varied)による亙るようだ。例えば第2次ニューロン(nonprimary afferents)により後索核に到着する情報は、そのうち若干は広い受容野をもち、多種刺激に反応するが、これらの核を経由して視床に送られるのだろう。
内側毛帯の繊維は全交叉。RFに側枝を出さないようだ(脊髄視床路とよい対照である)。体節的体性局在構成(後索や後索核でみられた)がこの知覚路の第2のlink,内側毛帯、でも又その終止する核においても保たれている。
| 脊髄視床路 |
後索核―内側毛帯ルートに加えて、脊髄視床路は意識にのぼる知覚である体の感覚インパルスを伝える2番目の経路である。解剖学的にも機能的にも第1番目の系とはいくつかの点で異なるが、全体としてのわれわれの知識はより少ない。(簡単化された図で2-12図をみよ)。
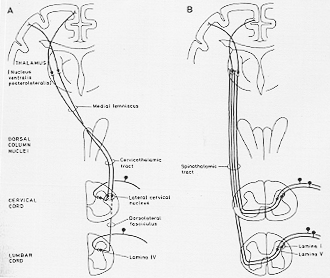
FIG. 2-12 Simplified diagrams which show the main anatomical features of the spinocervicothalamic (A) and the spinothalami (B) tracts. The termination of spinothalamic fibers in other parts of the thalamus than the VPL is not shown.
spinothalamic tractを特に
他の脊髄からの上行繊維とmixしている。
RFなどに側枝を出している。HRPで全く予期せぬことに同側C2 VII-VIIIで(+)ではっきりした起始細胞群あり。体側性のものはI,V,VII,VIII層に主として(+)
生理的仕事と相応する。
以前から(Petren, 1910; 他)ヒト前側索の傷害で身体の対側の温痛覚喪失が知られていた。 後索の前切載後で触覚と深圧核が残っており、また、前側索切載後にも一般に多少の触覚欠損を作る(Foerster と Gagel, 1932)ことから、脊髄視床路も触覚と深部圧覚を伝えるインパルスを伝えるに関与と結論されておった。
I層の細胞…強い機械的刺激や痛みを伴う熱感painful heatingの刺激でactivateされる。
V層の細胞…弱い触覚刺激より強い有害(noxious)刺激
V,VII,VIII層の細胞…deep structureからの刺激(muscular afferentが主として研究されている。)これらは又、加え、皮膚受容野をもっていることもしばしばである。
ある細胞は…関節の回転でactivateされこれらは、関節が固定位置に保たれているときslowly adapting dischargeを示す。
1つのタイプの刺激でのちactivateされるものや触覚刺激でも強い機械的刺激でも応ずるものもある(“wide dynamic range neurons”)
surroundタイプの抑制(lateral inhibition)は、内側毛帯系はたいへん特徴的なものであったが、脊髄視床路ではless commonである。しかしいくつかのunitsは反対側の身体の相応する受容野の刺激で特異な形の抑制がみられる(Willisら、1974)
脊髄視床路内で臨床データで感覚の性質に従った配列があるとFoersterとGagel(1932)が示唆したがサルで調べたところこのような局在はみられない。
しかし、繊維は体性局在的に配列している。これは診断に大切だ。前方に行く程はっきりしなくなるが、延髄や橋のレベルでも残っているようにみえる。
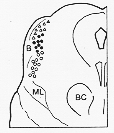
FIG. 2-13 Drawing showing the position of the spinothalamic tract and its somatotopic pattern in the mesencephalon according to Walker (1943). Triangles: lower extremity: dots: upper extremitoory; circles: face; B: brachium quadrigeminum inferius; BC: brachium conjunctivum; ML: medial lemniscus.
中脳レベルのspinothalamic tractsと局在の図(Fig 2-13, p.89)
このtractの腹側に三叉神経核からの繊維もVPLに体性局在パタンを示して終わる。
後索核と丁度同じく、脊髄視床路の起死核も大脳皮質から繊維を受け、細胞活動は影響をうける。
HagbarthとKerr(1954)はspinothalamic路を返る中枢への伝達は、大脳皮質の知覚運動野、小脳の前葉、RFからの刺激により抑制をうける。
後索核→R.M.起始核←縫線核
自然の痛み抑制のメカニズム、痛みの抑制に関連して近年注目されている。(内因性のモルフィン様物質を恐らく含んでいる?)
| Spinocervicothalamic tract脊髄・頚・視床路 |
(cp. Rexedの外側頸髄核Lateral cervical nucleus(→LCN) of Rexed C1,C2,)若干の解剖学者例えばRansonら(1932)によっても記載はされてはいたものの、外側頸髄核が特定の存在(実在)entityとして認められたのはRexedとBrodal(1951)の記載後からである。以来多少とも広範囲に研究された。この核は、後角のすぐ腹外側にC1,C2 segmentで(外)側索の内にある多極性細胞からなる細長い柱を形成している。ネコや2・3の動物にみられるが、他の形態を作って存在するものもあれば全く存在しない動物種もある。HaとMorin(1964)はヒトでは存在するが非常に小さいと主張するが、一方、Truexら(1970)の調べでは16ヒト標本中9例にみられたという。サルでは常に存在するようだが、恐らくネコよりも小さい形をとる(Ha と Morin, 1964; Mizunoら1967)しかしながら、サルやヒトではLCNの細胞のあるものは隣接する後角に包含されているという可能性を除くことができない。
LCNが小脳に繊維を送るという説(Rexed と Brodal, 1951)は、反証をあげられた(Grant, Boivie と Brodal, 1968)。-松下・細谷でごく少数+-HRPネコ-この遠心性繊維は頸上部で交叉し、内側毛帯の繊維と一緒になって上行する(Busch, 1961)変性実験とHRPで:LCNの大部分の細胞は対側の視床に投射し、VPLに体性局在をもって終わり、いくつかの他の核群にも終わる(後述)。2-12Aはspinocervicothalamic tractの模式図。
LCNへの主な求心系のソースは脊髄で、Brodal と Rexed (1953)によれば脊髄のすべてのレベルからの繊維がこの核に終わり、繊維は一次求心繊維ではなくて、脊髄灰白質にある細胞から来る。この繊維は、側枝の背外側を上行し、LCN全体に細かい側枝のprofuse plexusをもって終わる。Clarke柱には起始細胞はない。
HRPをLCNに注すると(ネコとイヌで)大部分の反応細胞は同側後角のIV層にみられ、少数がこれより腹側の層にみられる(Craig, 1978)。生理学的観察とも一致する。
LCNへの“下行性”の結合は、後索核の”reticular” part (Craig, 1978; Burton と Loewy, 1977)。大脳皮質の知覚運動領を刺激すると、通常spinocervicothalamic路の伝達を抑制するようである。しかしその効果は選択的である。つまり、nociceptorからspinocervical cellsへの入力は強く抑制されるが、一方毛包からの求心性入力は影響をうけない(Brownら、1977)。
脊髄(外側)頸(髄)核路は皮膚由来のインパルスを仲介するというRexed と Strom (1952)による最初の証明は、ひきつづく細胞外および細胞内記録を用いた生理学的研究のなかで確かめられおし進められてきた。脊髄頸核路spinocervical tractの起始細胞は、皮膚由来の太い有髄の求心繊維から単シナプス興奮性入力と、筋と皮膚からの(無髄繊維を含む)細い求心繊維のポリシナプス興奮性入力をうける(Brown, 1973: Boivie と Perl, 1975)。
多数の脊髄頸核細胞は、例えば毛の受容器に皮膚の直接的且つ、より強い刺激を要求するような受容器など種々のタイプの受容器からの収斂性の入力をうける。外側頸髄核の細胞はこれに相応する生理学的性質をもっている(Craig と Tapper, 1978; 他、をみよ)食肉類でみられるように、外側頸髄核(LCN)の細胞の3/4が、毛髪の運動で興奮させられるが、残りの他の細胞は皮膚への強い圧力やsqueeze(圧しつぶす。握る)を必要とする。
昔の研究で関節や筋肉からの入力を云々するものがあったが、LCNの細胞から直接記録してみるとこれは証明されなかった。
他の上行性知覚路におけると同様、spinocervicothalamic fibersの受容野は身体の近位部においてよりも遠位(末端)部でより小である。体部位局在もあることを生理学的に示している。
このようにspinocervicothalamic tractは弱い(軽い)皮膚刺激と侵害性刺激の相方の体部位的に構成された。情報を伝えることができるように思われる。感覚信号の全体的な伝達の内にあってのこの経路の正確な機能的役割は、しかしながら、不明である。
第3のニューロン連結(長・上行性・体性・知覚・繊維・系)Thalamic nuclei視床核脊髄視床路と内側毛帯の繊維を受け◎→大脳皮質へaxonsを出す。
ここで全体的に視床とその結合を一般的にのべることにする。(広範な視床皮質投射のうちごく限られた一部である)。
視床の多くのsubdivisionは機能的に異なるもの、2つの卵形の視床、III脳室で左右にわかれる。
3つの灰白質anterior (視床核)
medial
lateral(後方にのびてpulvinarを含む)。
これらは白質のlaminae板によって分けれている。これは横断切で、Y状にみられ、internal medullary lamina内髄板といわれる。(内に髄板内核あり)。intralaminar nuclei
正中部近くいわれるnuclei of the midline正中核がある。視床のほとんど全体の外表に沿って視床毛様核reticular nucleus of the thalamusと呼ばれる薄い細胞層ある。これと視床の主たる核との間には外髄板external medullary laminaとよばれる白質の薄い層がある。視床核と若干の亜核と結合関係が2-14図に示される。
動物間で原則的とは同じパタンを示が、あるものは、相対的に発達している。たとえばMD. LP.LD. Pulvinar CM (中心正中核)がヒトを含む霊長類でよく発達を遂げている。
Walker(1938c)。サルとチンパンジーの研究に主としてよっている-最も普通に認められたスキーマ(2-15図)。Hassler (1959) extendしてヒトで仕事。
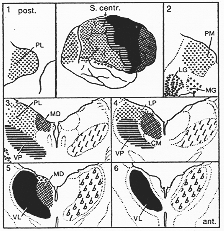
FIG. 2-15 Walker's diagram of the thalamocortical projection in the chmppanzee. 1-6: Representative sections through the thalamus, in a posteroanterior sequence. Corresponding areas of the thalamus and cerebral cortex marked with indentical symbols. LG: lateral geniculate body; LP: nucleus lateralis posterior; MD: nucleus medialis dorsalis; MG: medial geniculate body; PL and PM: nucleus pulvcinaris laterlis and medialis; VL: nucleus ventralis lateralis; VP: nucleus ventralis posterior; I: the area of termination of the fibers of the medial lemniscus and the spinothalamic tract. b: the area of termination of the fibers of the brachium conjunctivum. Slightly altered from Walker (1938c). Subsequent studcies have necessitated certain alterations as to details in the diagram (see text).
視床核の種々の分類に関する短い総説としては、Ajmone Marsan (1965)をみよ。前核anterior thalamic nuclei (3分)→すべて大脳皮質の内側面への投射。
内側核MD.→前頭蓋の種々の領域、および主としてorbital cortex外側核の下及び腹側部VA (ventralis anterior), VL(Ventralis lateralis), VP (Ventralis posterior)
外側核の背側部
VL→precentral中心前回
VPL,VPM→postcentral中心後回
LD→帯状回後部、頭頂葉の一部(恐らく)
LP→主に頭頂葉
Pulvinar→ 頭頂;側頭皮質(12章で詳述する)。
LG→中継核視(8章)
MG→中継核聴(9章)
体性知覚情報の視床中継の問題で特に関連して、posterior complexがある。この術語はM. Rose (1935) と J.E. Rose (1942)によって導入され、中脳-間脳連結部で視床の後部にある輪郭のはっきりし特に細胞周囲につけられた名称である。
RoseとWoolsey (1958)が、このareaから聴覚領およびその隣接領域にdiffuseに投射があることを示唆してから新たな関心がたかまり、その後Poggio と Mountcastlel (1960)が生理学的に、この領野は中枢性の痛覚機構上重要であるらしいと示唆を与えた。
この見解は、視床後部域posterior thalamic regionが脊髄視床の重要な終止域であることを示した多くの実験形態学的研究において相当の支配を得た(後述、文献はBoivie, 1971b, 1979; をみよ。且つ、Jones と Burton, 1974)。良染された正常ニッスル標本でP0-groupの境界をより正確に決めることができるようになった(Moore と Goldberg, 1963; Rinvik, 1968a; Jones と Powell, 1971; Heath と Jones, 1971a; Robertson と Rinvik, 1973; Jones と Burton, 1974; Burton と Jones, 1976; Boivie, 1978, 1979)
Posterior complexは均一質の構造物ではなくSG(suprageniculate), limitans核、MGBの大細胞性部分、Pulvinarのある部分を含んだ、いくつかの多方ともはっきりした細胞群から成っている(Olszewski, 1952により分がされたように)。Posterior complexはまた、VPとLPにはさまれた混じった細胞型の領域をも含み、内側方にcentrum medianum (CM)にひろがる。繊維結合状からもこの視床核はheterogeneousである(ことを示している)。すなわち、サルARGの仕事でBurtonとJones (1976)は、彼らのいうPosterior核の内側部はSIIの後方にあるretroinsular fieldに投射するが、外側部は、post-auditory皮質領野に投射する。SGとLimitans核はgranular insular areaに繊維を送る。
皮質⇔視床核間の相互結合
すべて皮質視床繊維はinfragranular layer顆粒下層の細胞から起こるが一方
視床皮質繊維の終止域は視床における起始域により相違なる如し(Jacobson と Trojanwski 1975)。
量的に大きい視床核は大脳皮質と密接関連を持っている(Macchi と Rinvik 1976の総説をみよ)。小細胞群の若干は、普通には、”cortical-dependent”でないと考えられているが、少なくとも若干の繊維を皮質に送る(Murray 1966, Scheibel 1967をみよ)。HRPで、髄板内核は、密な線状体投射の他に広くdiffuseに、しかしごく僅かだが、皮質に繊維を送ることが示された。この文脈の中で(考えてみて)、髄板内核への皮質からの遠心性繊維がよりMassiveである(と思われる)(Petras, 1972; Rinvik, 1972)のは興味深い。
皮質の特定域に強い投射をし対応する皮質からも繊維を受けている視床核は、本章を次章で考察されるが、可成りよく特徴づけられたextrathalmic源(体性感覚、小脳、淡蒼球、視床下部)からの求心性刺激によりbe acted upon活性化することは注意しておくべきである。
習慣的にある視床核をspecific, 他をnonspecificと呼ばれてきたが、これは、大体において機能的、電気生理学的観察を基礎として(にもとづいた)区別である。
非特殊核に共通するのは、電気的に刺激をうけると、長い潜時の後に両半球の広範の領域に活動電位が起こり、”recruitment”の現象を示す。これらの皮質反応は、特殊核刺激後に起こるよくlocalizedされた(局部的に限局された)急速に起こってくる電位Potentialと比較してよい対照を示す。非特殊核の大部分は正中部又はその近傍あるいは内髄板の内に位置している(図2-14, 図6-12)。視床網様核(R. Fig. 2-14)は一般に非特殊性と見倣されている。いくつかの視床核を“非特殊核”としてひとまとめに取り扱うことは教えるには便利だが、新しい解剖・生理学的知見が集積されるにつれて、視床核を”specific” と “nonspecific”とに分離することは簡単化しすぎであることが益々明白となった。これらの問題を“非特殊核”のことは6章の網様体のところで考察されよう。
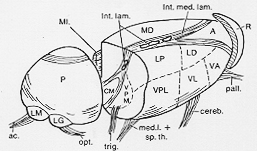
FIG. 2-14 Diagram of a three-dimensional reconstuction of the right humn thalamus seen from the dorsolateral aspect. The posterior part is separated from the rest by a cut to display some features of the internal structure. Only the rostral tip of the reticular nucleus is includes. The main afferent contingents to some of the major nuclei are indicated. Abbreviations for thalamic nuclei: A: anterior; CM: centromedian; Int. Lam: intralaminar; LD and LP: lateralis dorsalis and posterior; LG: lateral geniculate body; LM: medial geniculate body; MD: dorsomedial; MI: midlline; P: pulvinar; R: reticular; VA: ventralis anterior; VL: ventralis lateralis; VPL and VPM: ventralis posterior lateralis and medialis.
三叉神経主核→VPM(7章をみよ)
1970年代初期まではVPLは脊髄視床繊維の主たる終止域でもあった一般に認めれていたが、最近数年この考え方は深刻なる挑戦をうけた。今や内側毛帯と脊髄視床路は視床における終止に置いても何点か異なり、他にも各々の系においてインパルスを生じる末梢性刺激のタイプに関しても興味あること違う。
ネコ後索核
サル後索核 →(Lm fiber)規則正しいくさむら状終止パタン VPLの他にPOmにも終わる(Boivie 1970a, Jones Burton 1974)
VLP投射は多く体部位局在を示すがPOm投射は少なく局在を示さぬ。側枝かどうかもまだ明らかでない。いずれにせよVPLへの繊維は太く、POmへの繊維は細い。
脊髄視床路の正確な終止域は文献的に論争のあるところであった。
→VPL 他にPO. MGBの大部分。SG. CL (intralaminar)
この数年間で(Boivie, 1971b)(1970)
ネコspinothalamic→POm, CL, CLとVPO(吻側部)の間(HasslerのVim)(nucleus, ventralis intermedius)(VLPは-)
このareaはBrodal, Pompeiano (1957) の区核からの終止域に一致する(Grant, Boivie と Silfvenius, 1973)ネコ下肢からのgroup II筋球新繊維のインパルスの中継
サルspinothalamic POm somatotopic(-) に多数終わる(ネコより)
VPL somatotopic(+)全体に終わる(Boivie; Jones と Burton)
サルとネコの差(脊髄視床路)は一見和解し難く見えるが、サルのspinothalamicのVPL終止パタンとネコのcervicothalamic tractのVPL終止パタン酷似(Boivie 1970,1971).
Spinothalamic と cervicothalamic (食肉類)→進化のプロセスでjointサルのVPLへの房状のspinothalamic投射はネコのcervicothalamic投射にhomologous)!
この数年間の解剖学的が強く示すところによると、若干の重複がPOmと恐らくVPLの末梢部(上をみよ)にあるけれども全体としてみれば内側毛帯と脊髄視床路の繊維は異なる視床域に終わる。しかしながら、後索核からのインパルスは主にVPLで中継されはするが、VPLを内側毛帯-知覚皮質経路における純粋な中継核と考えることは極端な単純化であることを認識しておくことは重要なことである。不幸にして、この誤った見解はしばしば今の神経系の教科書にのっている。想起すべきは、後索核と脊髄からの求心繊維に加えるVPLは、他からも、例えば中脳RF、正中視床核、内側前頭束(medial forebrain bundle)(Scheibel と Scheibel, 1966e)、それに視床の網様核(6章)からも繊維をうける。それに、VPLは体性知覚皮質から大変多くの繊維をうける。それに:VPLは体性知覚皮質から大変多くの繊維をうける。この観点から、Ralston (1967)がネコ、EMでVPLでみられる終末ブトン中たった8%がLemniscal afferentに属している凍結論を思い起こすことは意味あることだ。
1950年-60年代のMountcastleと共同研究者の仕事 -single units stimulus- as well as place - specificityがVDL(cortexでも)大体に置いて維持されているらしい。VPL無麻酔サル1000units.
POm: central painメカニズムVPL: nociceptive cellsは大変少ない。
最終更新日:2002/09/13
