| 第3章.末梢性運動ニューロン |
要約:慶應義塾大学解剖学教室 川村光毅
・脊髄前角の運動細胞
・運動性ニューロンの配列
・運動単位
・運動単位の機能、筋電図
・筋肉内の受容器
・伸展反射と筋のコントロール
・伸展反射
・筋緊張

臨床神経学で普通に“運動系”とよばれるものの検索は神経系疾患における症候の解析の基本的部分である。注意深く検査すると、種々のタイプの機能障害が観察されるが、症状の原因となっている病理的過程の局在と型についての価値ある情報を与えるものである。末梢性運動装置-末梢運動ニューロンと身体の横紋筋-と中枢性運動の経路と系に区別される。後者は次章で考察される。
| 脊髄前角の運動細胞 |
末梢性運動ニューロン、しばしば運動ニューロンmotoneuronsと呼ばれるものは細胞体が脊髄前角又は脳幹の若干の脳神経の運動核内にみられる。後者は7章で詳述するが、原則として、前角の運動細胞と同型である。筋紡錘の遠心性支配が発見されて以後、大型と小型の運動ニューロン(通常α(アルファ)γ(ガンマ)運動細胞といわれる)に区別するのが習慣となっている。後者については後で又戻ってのべることにする。
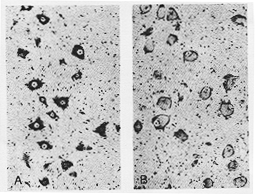
FIG. 1-12 Photomicrographs of motor ventral horn cells in the adult monkey. A: normal; B: 6 days following transection of axons. All cells present typical acute retrograde changes. ×68. From Bodian (1947).
α-motoneuron: 多角形で多極性で、細胞質に多量の虚班顆粒をもつ(1-12A図)。その多数の樹状突起が灰白質内主として前角の灰白質内で分枝している。軸索は腹側に走り、脊髄を出て前根に入る。この末梢神経はついで筋肉に至る。(すべての末梢神経がこのような体性遠心性の運動繊維を含んでいる訳ではない。すなわち、その大部分は、体性求心繊維と内臓性繊維をも有している。)
IX(Rexed)層内にある。しかし、多数の小型細胞を混じている。実際小型細胞の数の方がはるかに多い。ある群ではその比は1:16.5。前根繊維の約30%が筋紡錘支配である。従って、IX層にある小型細胞の大多数が筋紡錘への繊維への繊維の細胞体ではあり得ないことになる。若干のものは、確実に単に小型の運動ニューロンである。若干のものはRenshow細胞であるらしいけれども、他が介在ニューロンであるかどうか定まっていない。この主題は後述される。
脊髄前角及び運動性脳神経核内の運動ニューロンは、身体の骨格筋につたえれるすべてのインパレスにとって“最終共通路”(Sherrington)をなすものである。それらの運動ニューロンはたくさんの源泉からのインパルスにより影響をうける。前述したように、一次知覚繊維がそれらに終わることが見い出されている。それに、大脳皮質及び脳幹諸核から脊髄を下行する繊維によって、主として介在ニューロンを介して、活発化される(acted upon)。(4章で論ぜられる)。故に運動ニューロンの行動の究極的効果として起こる運動は、運動前角細胞に収斂するインパルスによって規定されている。そしてこれらのインパルス(効果)は、いわば、異なる系列より演じられているのである。
種々の運動の多様性は例外を除いて運動ニューロン自身によって第一義的に決定されているのではない。
| 運動性ニューロンmotoneuronの配列 |
IX層の運動細胞は、脳神経運動核のあるものと同様、特徴的仕方で配属している。前角にいくつかの細胞群として脊髄横断切片状でみられる(3-1A図)。-縦方向に配列。この柱は一定の分節内にみる。逆行性細胞変性法で調べれてきた。ヒトの四肢切断例や神経損傷れでも。Sharrard (1955).ポリオ例の麻痺の分布と前角細胞消失を注意深く調べ明らかにされた1つの基本点は、内側細胞群は体幹と頸部の筋を支配する。これらの細胞群(より正確には腹内側柱のみ)は脊髄全長にみられる。他方、外側細胞群は主として頸および腰膨大部にあり、四肢を支配する。この柱内にもパタンがあり、四肢の遠位の筋を支配するものは、近位筋支配ニューロンよりも通常dorsalに位置している。四肢の遠位部に関連する細胞群は膨大部の尾方部にのみ発達することは注目されよう。motoneuronは、したがって体性局在的配列をする。(Fig.3-1Bをみよ)このような配列があるので、ある特定レベルの前角or運動ニューロン局所障害をおこすと、大節的に区分あれた不全麻酔や(完全)麻痺paralysisをおこす(例えば、C5-6が傷害された時(ポリオなどで、-灰白髄炎)三角筋、棘上筋、上腕二頭筋、烏口腕筋、腕橈骨筋、上腕筋の麻痺がおこる。3-1B図や2-4図の皮節dermatomeマップと比較せよ)。
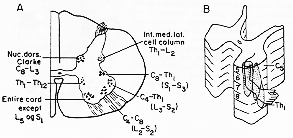
FIG. 3-1 A: Diagram of the human spinal cord presenting in a transverse section the position of the various cellular columns. The letters and figures indicate the segmental distribution of the cellular columns. Redwawn from Kappers, Huber, and Crosby (1936). B: Three-dimensional diagram of the gray matter in the cervical cord, showing the somatotopic arrangement of the groups of motor ventral horn cells which supply different parts of the upper extremity. The hand should cover a larger area than shown in the diagram, since the distal parts of the upper extremity have a far ampler motor supply (cf. small motor units) than the proximal parts. See text.
Romanes (1953)はこの主題を調べ上げた上で以下の如く結んでいる。明らかに運動細胞群は筋群の形態学的区分を部分的に現わす。が、また、大部分の哺乳類で動かされた関節に関連する局在的意味もある。そして時としていくつかの部分は単一の筋を現している。
| 運動単位motor units |
Sherringtonにより導入された言葉で、極めて有用な術語である。一つの運動単位は、1つのα-運動ニューロンとその軸索とそれが支配する筋繊維(複数)である。1つの運動前角細胞が放電dischargeしたとき、この単位に属するすべての筋繊維群は収縮する。この単位に属するすべての筋繊維群は収縮する。1つのmotoneuronはvery fewから1000又はそれ以上の筋繊維を支配する(ものである)。一本の神経(突起)従って1つの運動ニューロンあたりの(支配する)筋繊維の数をしらべる。筋の内でも、特に外眼筋や手の小節などは運動単位は小である。霊長類では、手の小筋や喉頭筋、外眼筋のような細かい分化した運動に使われる筋では運動単位は最小であると思われる。
ex. Platysma 25
gastrocnemius 1600-1900
小さい運動単位の神経の径は径も小である。軸索の枝分かれにより多数の筋繊維を支配。この枝分かれは、神経繊維が筋に隣接したときはまた筋肉でおこる。
1つの運動単位の筋繊維は筋の全長にわたって直列分布をしているようだ。筋の一定部に集まるに非ずして (cooper 1929の生理実験から考え)
cp. グリマーゲンdepleting消耗法(Edstrom と Kugelberg, 1968)頻回刺激→PAS染色。
運動神経繊維の終末分枝(岐)は、いわゆる運動終板motor end platesの一部として筋肉内にみられる。運動終板は2部より成る。一つは神経性muscular筋性であり20-30nmの隙(すきま)で隔てられている。この筋・神経結合myoneuronal junctionは多くの点でCNS中のシナプスに似ている。終板の筋肉部はいくつかの核と多(無)数ミトコンドリアを含んでいるが、収縮性のcontractile myofibrils筋原繊維を欠く。神経部は多くのぶどう状のふくらみがみられ、さながら終末ブトンの如くである。この各々は筋繊維の中に入り込み”dips into”(侵入)し、シナプッス小胞とミトコンドリアやを含む。アセチルコリンacetylcholineが終板で伝達物質として働くことが確立したアセチルコリンは、間隙には入り、筋の細胞膜と結合(Bound)し、このイオン透過性を変える。
一つのインパルス→1つの終板終板電位(ポテンシャル)を筋繊維に起こす。充分な高さに達したとき1つの活動電位action potentialを生ぜしめ、これが繊維沿って伝播し、propagate、筋繊維の短縮を引き起こす。Ach:セチルコリンエステラーゼ(酵素)により速やかに破壊される。組織学的方法により、終板がこの酵素の存在のために可視化される。
神経筋接合部での刺激伝達はいろいろな影響をうける。
クラーレcurare, e.g., 終板電位を減少させ、遂に活動電位の出現を妨げる。→筋の麻痺が起こる。
(クラーレ様物質を筋弛緩を得るために麻酔中に使用することを比較せよ)
重症筋無力症の本格的傷害は、終板における伝達の欠陥である。EMGで記録されたポテンシャルは上述した筋繊維の活動電位である。
通常、一つの筋繊維は、一つの軸索によってのみ支配を受けており、単一の運動終板を有している(Feindelら、1952をみよ)。
運動単位(数)は生後減少する。恐らくはじめは各筋繊維は複数の運動ニューロンで支配を受けているため(Brown, Jansen とvan Essen, 1976)。成人パタンを獲得した後(1軸索-1筋繊維)(ラットで生後2週位)運動単位のサイズ(数)は一定で不変となる。
| 運動単位の機能、筋電図 |
運動単位は、運動装置locomotor apparatusの最小の機能単位である。運動における種々のことがら(差)-その範囲、強さ、タイプ-究極的には、運動単位の相互作用と共同作用の相違により決定される。これらの基本単位の機能に関する情報は、収縮中の筋肉の電気的変化の記録である筋電図を使用することによって特別に得られる。
皮膚上に表面電極、筋肉は表在性のもの(e.g. 背側骨間筋 muscle interossei dorsales)
深部の筋には微細な双極電極針を刺入して誘導した電位の変化を記録する。電位は1つのmotor unitの活動を反映すると考える。
筋収縮力は、一部はactivateされた(recruitment)運動単位数に、一部は運動ニューロンが支配する筋繊維に送るインパルスの頻度に依頼する。
EEGはα-運動ニューロンの活動を反映するが、その活動に関与する情報は比較的粗雑で不完全である。
現今の総説本としてMatthew’s(1972)をよまれたし:Handbook of Sensory Plysiol. Vol. Ⅷ/2. “筋受容器”たくさんの未解決の問題が残されている。骨格筋にはいくつかのタイプの受容器が備わっている(3-3A図)。すなわち、筋紡錘muscle spindle,腱器官(ゴルジー器官)、Vater-Pacinian小体、関節の滑液色capsules of the synovial jointsにあるいくつかの終末タイプ、および自由神経終末である。これらのうちはじめの2つだけ、ここで考えることにしよう。しかしながら、皮膚にある他の受容器のタイプも運動ニューロンに影響を与えることが明らかにされている。パチニー小体と関節の受容器についてはすでに第2章で考察した。
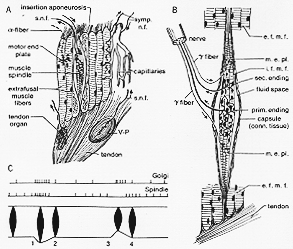
FIG. 3-3 A: Diagrammatic drawings of the three main receptors in skeletal muscles: a muscle spindle, a tendon organ, and a Vater-Pacinian corpuscle (V-P). Note that the spindle is arranged in parallel with the extrafusal muscle fibers, while the tendon organ is arranged in series with them. This has iomportant consequences for rtheir function. B: Very simplified diagram of a muscle spindle to show its principal components (see text). Abbreviations: e.f.m.f., extrafusal muscle fibers; i.f.m.f., intrafusal muscle fibers; m.e.pl., motor endings. C: A diagram illustrating the response of the muscle. The muslce is dwawn below in black, between lines indicating its insertiion. Action potentials are shown in two top lines. Further explanation in text. Fig. C from Granit (1955a).
これらの受容器のうちで筋紡錘が形態学的にも機能上でも最も緻密(5)で複雑なorganizationである。Ruffiniによりほぼ完全に記載されている。すべての筋(運動装置としての)にあるようだ。喉頭筋、咀嚼筋、舌、外眼筋など運動性脳神経支配の多くの筋にもある。筋紡錘存在の密度とは差がある。ヒトの 1 gr. あたりの数
(短母指外転筋) abductor pollicis brevis (小筋) 80ヶ 29.3 デリケートの運動
(広背筋)latissimus dorsi (大筋) 368ヶ 1.4 粗大な運動
伸張受容器、muscle spindleの名前はそのshape形に由来している。構造の基本(主要)点は、(3-3B図をみよ)。
紡錘状の形をした結合識のの包capsuleが、数ミリメーターの長さにわたり、数本3-8(ヒトでは平均10本Cooper, 1960)の細い筋繊維を包む。この筋がいわゆるintrafusal fiberの筋内繊維は、その端が包capsuleの結合識に付着しており、その間のスペースには液体が充たされている。
このように、錘内(筋)繊維は、普通の錘外筋繊維extrafusal muscle fiberと並列的(同方向に)に配列されている。そして両者は、結局は、直接又は関節に非弾性inelasticのコラーゲン繊維膠原繊維より腱に付着している。(3-3A図をみよ)。筋外転繊維が収縮するとき、筋紡錘の繊維は張力(tension)が減じられ、一方、筋全体の伸展は錘内筋繊維の長さと張力(tension)が増大する。錘内繊維の赤道部中心は収縮せず、多くの核を有す。その部分は螺旋状の神経繊維である(第)一次(知覚)(感覚)or annulospiral終末(螺旋形終末)は、単一の比較的厚い求心繊維(ネコ、12-14 μm)から成る。(第)二次(知覚)終末又は、flower-spray ending散形終末はより細い求心繊維で、環螺旋形終末の1側又は両側とみられる。更に筋紡錘は運動支配を受けている。これは比較的細い繊維繊維で以前に(から)γ-繊維(or fusimotor錘運動繊維)、と呼ばれ前角より来る。この繊維は錘内筋繊維の遠位のcross-striatedの部分に終わり、これが収縮する。これからわかる(云えること)は、γ-繊維の刺激に反応して、起こる錘内筋繊維の遠位部の収縮が、錘内筋繊維の中心部の“感覚”部分の伸展を引き起こし、知覚繊維を結果的に刺激する。同時に筋全体の伸展をともなう如くに。腱器官は、筋紡錘よりも一層簡単(な構造)を示している。本質的に(本来は)腱器官は、ふつう筋、腱結合部近くにある腱の一群の膠原繊維間に散形の細い終末をもって終わる太い有髄神経繊維(12-18 μm)の一群の枝から成っている(3-3A図)。単一の繊維が1つ以上の腱器官支配することもある。これらは普通デリケートな結合識の包capsuleで被われている。腱器官の機能を理解するためには、それが錘外筋と直列in seriesに配列されているということを知っておくことが必要である。筋肉が収縮したときも伸張されても、腱器官は刺激を受ける。なぜなら、いずれの場合にも,腱の張力は増大するから。
この形態からの類推は生理学的に確かめられた。(筋紡錘と腱器官に関する見解)
後根単一繊維からの記録(活動電位)
これらの所見から腱器官は、張力の記録係(tension recorder)であり、一方筋紡錘は筋(肉)の長さに関する情報を提供すると云える。これらの重要な知見はMatthew’s (1933)によりえられ、後年他の人たちにより確証された(Granit, 1955a;1975, -for reviews).
筋紡錘の遠心性支配に関しては、Leksell (1945)前根の細い繊維から複合活動電位を最初に導き出すことに成功した(20-38 m/secの伝導速度を持つ)。γ繊維を刺激しても筋の収縮は起こらぬが、求心性の活動電位は筋神経から記録できる。この電位は、筋紡錘から起こるものと解釈された。γ-繊維が活動したとき遠位の収縮部分が収縮することによって錘筋内繊維の中央の感覚域が引っ張られることにより起こる。Leksell (1945)の所見は、幾度となく単一繊維法により確かめられたが、筋紡錘はCNSからの支配下にあることを明らかにした。muscle spindleの遠位収縮部の収縮が中央感覚部を伸展させ刺激閾値を下げるので(ことから)。筋紡錘のγ-繊維の刺激の程度は伸長に対するニューロンは多くの源からのインパルスで興奮する。これが又、spindleの活動に影響を与える(下述)。
Eccles, Eccles, Iggo と Lundberg (1960)細胞内記録法
γ細胞は同じ筋肉を支配する運動ニューロンに混在する。
Nyberg-Hansen (1965b) modified Gudden法で証明、Burkeと共同研究者(1977) HRP. IX層内、大小の標識細胞、最も小型の細胞10-38 μmの径、 γ-ニューロンを構成していると推量された(Pellegrini, Pompeiano と Corvajaをもみよ)HRP注入で(ネコのsoleus, 内側gastrocnemius)これらの筋を支配している細胞柱内の96%が標識された故、IX層には介在ニューロンはあったとしても非常に少ない。
その後の研究で、筋紡錘の構造と機能は複雑となった。
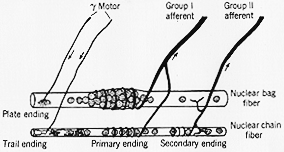
FIG. 3-4 Simplified diagram of the central region of a muscle spindle, showing the two kinds of intrafusal muscle fibers and their sensory and motor innervation. The presence of two types of bag fibers in not shown. The nuclear bag fiber in the diagram may be taken to represent a bag1 fiber while the nuclear chain fiber represents both chain and bag2 fibers. (Slightly changed from Matthews, 1964.)
ネコでmuscle spindle赤筋slow twitch, fatigue-resistant型に殆ど必ず存在する。運動単位は小の筋、省エネルギー・デリケート運動
白筋、実際上存在せず(muscle spindleは)。
表在性のmuscle spindleは深在性のmuscle spindleに比べて感受性が劣る。
20年前に錘内筋繊維(3-4図)はとりわけ直径の異なる2つのタイプありとされる。その核の配列に関して
太い繊維:nuclear bag fibers核袋繊維
細い繊維:nuclear chain fibers核鎖繊維
最近bag fibersにはbag1とbag2の2種が組織化学的、電顕的に区別される。(Barkerら1976をみよ)。
機能上も異なるという証拠もある。 Bar上もmuscle spindle支配について複雑となり。
一次(感覚)(らせん形)終末は核袋・核鎖の両繊維にみられ
二次(感覚)(散形)終末は、主に、しかし常にではないが、核鎖繊維のみに存在する如く。ヒトのmuscle spindleはネコのmuscle spindleと本質的に類似している(Cooper と Daniel, 1963, 1963; Swath と Fox, 1972)。伸展に対する筋紡錘の反応特性は単にlength recorder (筋の長さの記録係)だけではないことを以前の研究者はみていた。Jansen と Matthews(1962a)は、一次求心繊維から記録された電位は筋の長さのみならず伸長速度に関する情報も与えることを強調した。前者の反応は静的反応static responseと呼ばれ、筋の維持された伸びの際にみられる。動的反応dynamic responseは、実際に伸ばされ(つつあ)ることに対する受容器の反応である。種々の理由から著者らは以下を示唆した。動的反応は核袋繊維に起因し、静的反応は核鎖繊維の伸展による:この両タイプの繊維とも一次終末により支配されている。これに一致して最近
Boyd (1979)ら bag1 fibers主に動的反応動的相 (初期)速度感受性
bag2 fibers
chain fibers 静的反応、静的相 位置感受性
二次終末は多くの点で一次終末と機能が似ているようだが、長さの変化、とくに急速変化に対する感受性が低い。(Bessou と Laporte,1962; Matthews, 1964)。又tonic stretch reflex緊張性(or持続性)伸張反応、に実質的に貢献はいくつかの間接的証明あり(Matthews, 1969, 1972)。
更に生理学的にγ-繊維にいくつかのタイプであり。
Jansen と Matthews (1962a,b)。一次終末の動的及び静的感受性が除脳ネコで独立にコントロールされる。Matthews (1962)・・・static, dynamic fusimotor fibersと機能的に異なるBarker et al (1976)グリマーゲン消失法で繊維の型を分けて命名した。
伸展に対する動的な特性を高める・・・bag1型-動的γ繊維のみにより影響される。
特定の長さまで(錘内)筋をのばしたときに発射のレベルをたしかめる。・・・bag2型と核鎖型繊維-静的γ繊維のみにより影響される。
筋紡鎖へのγ繊維の終止(末)。2種類あり、 Barker (1962, 1966)命名
ガンマ板状終末plate endings 錘外筋の運動終板状
ガンマトレイル終末trail endings 細い軸索突起の広範な散形のネットワークで小さく終末が膨らんでいる。
生理と形態の技法を結べつけて少なくとも通例として以下の如く確立された。
plate endings mediate the dynamic fusimotor effect (or bag1 fiber)
trail endings are responsible for the static effect (on bag2 と chain fibers)
β(ベータ)繊維:
錘外筋と錘内筋に共に分枝を出している遠心性繊維組織的に証明(Adal と Barker, 1965)。αとγ運動ニューロンが共に、特別な筋の随意筋収縮の際に、activateされる(Vallbo、1970a,b,1974a,b)。-これがGranit (1955a)のいわゆるαとγ共同作業、α と γcoactivationである。随意収縮中のγ-運動ニューロンの活動は、筋紡錘の感覚域のshorteningを妨げる。こうしてこの(γ-)運動ニューロンを刺激してmuscle spindleからのインパルスの流れが、筋を力の減少を恐らく妨いで短縮させながら、維持される。これに一致して、筋神経中の細い繊維(その中にγ-繊維がある)が選択的にブロックされると、運動の力と速度が減ずる。しかし、これらは共に、筋腹に振動vibratingを与えると回復する(Hagbarthら、1970; 1975)。この方法は筋紡錘からの求心性をactivateする強力な一つの方法である(Granit と Henatsch, 1956; Matthews, 1966; Burke, Hagbarth et al., 1976 a. b.).
振動を与えると→筋の特異な反射的収縮が起こる→tonic vibration reflex
長い潜的(30秒以上)とゆっくりした(緩除)なrise and full
この収縮contractionに伴って、拮抗筋antagonistが抑制され、振動を受けた筋の腱反射も抑制を受ける。時にこの緊張性(持続性)振動反射に随意的に完全におさえられるが、検者の注意を他に向けるようにするとすぐに立ち戻ってくる。tonic vibration reflexでspindle afferentsが主たる求心性のlinkを構成していることは明らかであるが、その中枢性のメカニズムは分かっていない。とりわけ、その長い潜時を有していることから、polysynaptic pathwayにちがいなく、恐らくtranscortical皮質を通るものであろう。
筋紡錘はCNSに筋の長さと伸張のスピード(速度)を知らせ、こうして、脊髄反射装置の適切な活動に必要な情報を供給する。加えて、筋紡錘は、supraspinal level例えば大脳皮質にメッセージを送ることにより筋をコントロールすることに貢献する。われわれは、伸展反射と筋のtone(緊張)を考察するときに再び筋紡錘に立ち戻ろう。
腱器官は筋紡錘ほどには詳しく調べられていない。前述の如く、腱器官は筋の収縮によるものであれ、伸張によるものであれ、筋内の張力tensionをrecord記録するようにarrangeされている。以前に過度の伸張に対しての保護が主な役目とされていたが、よりはっきりした(分化した)機能を持っているようだ。そして事実、筋伸張で起こる張力に対するよりも筋収縮により起こる張力に対してはるかに感受性を有している。
Pacinian corpusclesは振動受容器と思われる。
| 伸展反射stretch reflexと筋のコントロール |
他の脳から分離されても脊髄は体性および自律性の多数の反射を仲介するmediateことができる。一つの神経反射の形態的基盤は一般に反射弓reflex arcと伝われている。きっと簡単な形で5つのlinksから成る。
ここでは脊髄反射、骨格筋-体性反射
基本原理(脊髄の反射活動を支配している)はSherringtonによって明らかにされた。
Flexor reflex (nociceptive) - withdrawal reflex
extension reflex
Scratch reflex
伸展されたことに反応して単一筋が収縮する。おそらくすべての筋で起こる基本的な反射である。
伸展反射は多くのいわゆる姿勢反射(第4章参照)の基礎を作るもので、大づかみに云って、身体の正しい姿勢の維持をし、種々の要求に適応することを目的としている。それが外的な力によるものであれ、生物体自体によりなされた運動によるものであれ。
伸展反射とその解剖学的基礎を論ずるに、比較的簡単なモデル、patellar reflex膝蓋腱反射から始めよう。単シナプス反射、恐らく機能的意義は全くといって良い程ない。必要とする刺激があまりにも非生理学的であるので。又、この反射はすべての筋を起こるものでもないし、健康人でも全く欠如するものもある。
単シナプス以上の長い潜時で起こる筋の伸展に対する反射反応はreflex (responses)は明らかにより大きな機能的意義がありHammond (1958) の記載以後注目された。これらの多シナプス伸展反射の関与するpathwaysについてはよく分かっていない(以下をみよ)。上述の反射は臨床で腱反射or深部反射といわれる。どちらも真に適切なものとは云えない。以下でみるように。myotatic(筋張力性…腱伸展…)反射という名称がしばしば用いられるが、これは、適切である。しかし伸展反射stretch reflexという術語が今日最もよく使用されている。図3-5でpatellar-reflexの説明あり、(p.164-165)。phasic相同性stretch reflex
長い潜時の伸展反射
ヒト二頭筋の伸展 25 msec(単シナプス性)
50 msec(多シナプス性)→大脳皮質が関与する(Philips, 1969,が最初に示唆した)
muscle spindleからの情報が大脳皮質に達することはよく知られている(cp.2章)一次終末も二次終末も。transcortical reflex loopの侵害でこの長潜時の伸展反射はヒトで消失する。緊張性tonic stretch reflex?
姿勢における緊張性伸展反射によって演じられる正確な役割と姿勢適応postural adjustmentはまだ明らかにされていない。
休止時正常筋一定度の緊張tensionをもっている。(さわってみてわかる)。
この緊張は受動運動の際にも感じられる。この正常時の筋緊張状態と通常muscle tone(筋緊張)、更に適確にはresting tone。緊張又hypertonus hypotonus
臨床検査
ex. 内包性麻痺
γ―ニューロン spindleの感受性を変える。
γ-ニューロンの活動が亢まるとspindleをよりactiveなレベルに”set”して錘外筋の緊張を増大させる。使用される筋の状態を決定しそのコントロールを動かせるにγ-ニューロンは大切。(cp.α-ニューロンは収縮させるだけ)
又γとαのcollaboration(運動を行うにあたり)は研究さるべきこと。
前述の如く
CNSは静的γ-ニューロン動的γニューロン、αニューロンを個々にコントロールすることが可能なり。しかしどの程度まで?
腱器官からのafferentsはmotoneuronにdirectにつかぬ。介在ニューロン→α-motoneuron(図3-5A)。
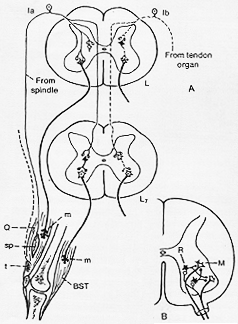
FIG. 3-5 A: Diagram showing (to the left) the pathways involved in a stretch reflex. Impulses arising in the muscle spindles are transmitted in group Ia fibers. To the right are shown (dotted) pathways arising in the tendon organs and passing in group Ib fibers. Excitatory cells and synapses are shown as open structures, inhibitory as black. BST: flexors of the knee; m: motor end plate; Q: quadriceps muscle; sp: muscle spindle; t: tendon organ (cf. text). Slightly modified from eccles (1957). B: A diagram of the proposed anatomical basis of the inhibition of motoneurons (M), occurring through their recurrent collaterals to Renshaw cells (R). Modified from Eccles (1957).
最終更新日:2002/09/13

{kind=link}