| 第4章.脊髄への下行路.大脳基底核 |
要約:慶應義塾大学解剖学教室 川村光毅
・錐体および錐体外の運動系
・錐体路に関する見解の変遷
・錐体路の経路
・皮質脊髄路
・錐体路繊維の起始と終止
・伝達速度
・赤核脊髄路
・前庭脊髄路
・網様体脊髄路
・脊髄への他の下行路
・視蓋脊髄路
・間質核(カハール)脊髄路
・弧束核脊髄路
・基底核といくつかの関連した核群
・線条体
・視床下核(Luys体)
・黒質
・基底核と関連核の繊維結合
・淡蒼球黒質繊維
・他の黒質求心繊維の起始核として
・黒質視蓋投射

読者はこの本に「錐体路」と「錐体外路(系)運動系」の項が含まれていないことに気づくであろう。この項はほとんどの教科書にもあるし、またこの書の第1版に盛られていた。一般使用から離れているので多少異端であるもので少ないコメントの要があろう。
| “錐体”および“錐体外”の運動系 |
大脳皮質と脳幹の多くの核は、直接間接の下行路により脊髄の活動に影響を与え得る。これら諸核は、他から求心線維を受けており(例えば小脳)、それ故これらの核を至る脊髄への“高次レベル”からのインパルスの伝達は神経路がシナプスで絶たれる場所でmodifications修飾をうける。
脊髄に直接至り、その活動に影響を与えるであろうsupraspinal structuresからの神経繊維束のいくつかが4-1図に示してある。機能上も異なる。正常の状態でこれらは統一的方法で協同することが推量される。しかしこの共同作業の分析は可成りむづかしく、多く為すべきことが残っている(山程ある)。
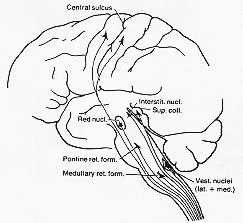
FIG. 4-1 Diagram of fiber systems descending to the cord from supraspinal levels: the corticospinal, rubrospinal, tectospinal, interstitiospinal, pontine and medullary reticulospinal and vestibulospinal tracts. Some minor tracts, such as the solitariospinal tract, are omitted. All of these connections may influence the activity of the peripheral motor neurons.
昔は、一般的に脊髄への下行性繊維系の仕事は、その遠心性機構に働き、体性及び内臓性遠心性ニューロンのインパルのdischarges(発射)を決定づけると考えられていた。1950年代以来、これらの下行繊維系は、この他にもう一つの重要な機能。しなわち、第2章で述べた如く。末梢から脳への知覚インパルスの伝達を調節regulateするという機能をも有していることが結論的(決定的)に証明された。これはHead, Brouwerら臨床家により仮定されていた所だ。
更に、これらの繊維系の多くは脊髄の反射活動に影響を与える。だから下行路を一括して“運動”繊維系とすることはもはや適切ではなく許されさえもしない。このような概念の不支持は、上述のように、脊髄の遠心性機構に対して影響を与える繊維を送っている神経核がその適正な機能遂行に必要な感覚情報を備えた上行繊維を受けているという事実によっても更に支持されている。同時随伴的に起こる感覚現象を考慮せずに“運動”機能をとりあつかうことは一つの理論的抽象である。Jung と Hassler (1960)「感覚(知覚)コントロール無しの“運動システム”は虚構であり、その虚構(つくりこど)には有益性さえもない」。
この2~30年代の進歩で古典的概念の改築が必要にも不向き、メンタンで又つかいやすいことから、不適当と知りつつもいつまでも生き続け心に残る。無知を隠す格好のラベル”pyramidal”, “extrapyramidal” “upper motor neuronsについて少し言及してみる。
錐体路は定義にしたがえば、起始のいづれをを問わず、延髄衰退を縦に走るすべての繊維のこと。大部分皮質脊髄路。corticospinal; corticobulbar(→運津性能神経核→時にreticular formationや他の延髄核へ行くものを含めている。)も-共通術語として、錐体路症候群というのは誤った名称である。
錐体路の傷害のために生じる症候を意味するが、実際には内包や皮質の特定部の傷害でみられる運動性及び反射性の変化が記述されている。しかし、これは錐体路の真の、純粋な傷害でなく。皮質でも内包でも必ず混合した傷害あり、唯一の錐体路症候群を作り出せる傷はpyramis内のみ。錐体外路という言葉は本来解剖学的概念で厳密には“錐体路”以外のすべてを含む。しかしながら、暗黒大の資格(権限)を付与されて、現在。“運動系”(錐体外路運動系)を示し、運動活動に影響を与える錐体路以外のすべての下行路とその介在する神経路包括する名称を意味する如くに使用されている。小脳も含め神経系のいくつかの他の部分の(核)も、運動装置の機能に対して影響を与えることから、どれが‘錐体外路系’の構造物かの概念を限定づけることはむづかしい。ある研究者はその内に、橋核や小脳を含めているが、特に大脳基底核はextrapyramidalのうち1つの統合的部分integrative partをなしていると考えられている。しかしながら、解剖学的にこの‘系’を限定し、定義することはできない。
臨床的には“錐体外路性疾患”…錐体路症状とは異なる運動機能障害。主として、筋の緊張の変化と種々の不随意運動、大脳基底核傷害の後にみられる症候群が含まれる。
Oscar と Cecile Vogt (1920)…‘striate system(線条体系)’と呼んだが。線条体、淡蒼球、視床下核、および他の核と路を含む。後日歯状核などの他の脳部の傷害でも似た症状が現れて錐体外路系の概念が拡大された。症状に多少の共通性はあるものの共通の事項の下でこれらと考察する利点を見い出すに困難を感じる。この上定義の定まらぬ系を云々するのであるからなお更である。“錐体外路疾患”という名前を残しておく唯一の理由といえるものは、理論的に根拠にもとずいて各々の特定の症候を分析する問題を避けられると云う位のものであろう。
“錐体外路系”を“錐体路系”から分けるのは、過去にコントラストをはっきりさせようとの意図で為されたが人為的なものである。両者とも正常機能の働いている生物(有機体、being)では共同(協力)しなくてはならず解剖学的にみても分してしまうことは同じく不可能である。
“上位運動ニューロン, upper motor neuron”は時々使用されるが、特にその用法は錐体路(上位運動細胞疾患)の傷害による症状に関連しているようだ。たくさんの”upper motor neurons”があるので、この意味合いでこの術語を使うのは誤りを導くもので避けることが望ましい。
著者(私)の深く信ずる所に従えば、理論的にも実際上も、錐体外路という言葉を残すことに何らの有益なる目的もない。
そもそも起こりは延髄の錐体pyramisここを繊維が独立(孤立)して走る。Turck (1851)ヒト内包出血-痛性半身麻痺材料 大脳皮質→脊髄 記載FritschとHitzig (1870) イヌ前頭葉の電気刺激後 対側肢の運動 同じ頃Hughlings Jackson (1970) 臨床観察から、体局性の“脳に運動局在”がある筈。正規の変わり目に以上のパイオニアの仕事をいくつかの仕事がでてelaborateした。GrunbaumとSherrington (1902, 1903)サルの前中心回で体性局在パタンを存在を、皮質刺激と剥離で示した。同類の結果が皮質傷害臨床例からでた。HolmesとMay (1909) 上部脊髄で錐体路切断、サル、逆行性細胞変性4野Betz巨大細胞のみに現れた。これで起始が決定さたかにみえた。以後 錐体路の概念が生まれ、その後約40年前に到るまで一般に支持(held)された。即ち、錐体路は中心前回から起こり脊髄に向かい下行する。その繊維は太く有髄。脊髄では末梢性運動細胞と接触する。錐体路は随意のとくにこまかい個別的なきめこまかいdiscrete運動の(伝達)(mediation) or仲介に関与する。この考えに従えば錐体路はむしろ構成が簡単なもので明らかに定まった1つの機能をもっているものである。以後に疑問、LassekとRasmussen (1939)ヒトの錐体で繊維を数えた、この錐体では皮質脊髄路の大部分が離れてしまっている。Ag impreg法で計算したところ、各(一側)錐体に約100万本の繊維がある。最近DeMyer (1959) 749,000~1,391,00で平均1,087,200本、(21例)。4野のBetz細胞は(5層)25,000~30,000 (Campbell, 1905 ; Lassek, 1940)故3-4%にすぎぬ。錐体路の繊維の大多数は前中心回のBetz以外であることは自明でしかも、4野以外からも出る(後述)。
| 錐体路の経路(路) |
皮質-内包(後肢の比較的せまい(小)域)(Fig. 4-9, p.212参照)内に視床、外にレンズ核、皮質に橋路、皮質―赤核路、淡蒼球-視床路内包内に含む
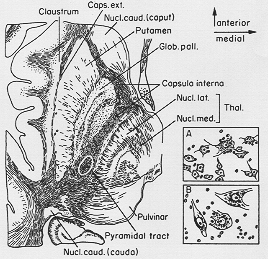
FIG.4-9 Drawing of horizontal section through the human brain, showing the corpus striatum (myelin-sheath staining). Redrawn from Ranson (1943). Circle in dicates the approximate position of the corticospinal fibers in the internal capsule. Below to the right, representative cells from the striatum (A) and the pallidum (B). Redrawn from Foix and Nicolesco (1925).
cp. Foersterの図は多くの教科書に転記されているが、内包の後肢全体域と膝部にこの繊維路あり(ヒトでは誤り)。
大脳脚の正中部2/3を錐体路が占めると良く言われるが、之は道理にかなっていない。というのはヒトで一側大脳脚にある約2000万(20million)本の繊維(Tomasch, 1969)のうち皮質脊髄繊維は約100万本(1million)であるから。大脳脚の繊維部分のパタンはFig4-2を参照。これはヒト、サル、ラットの所見を参考にして作ったものだが、恐らくこんなにはっきりとは区別されていない。橋核ではpeduncleからの繊維が数束に分かれ橋核細胞の大きな細胞の周囲の中に入っていく。(後述、小脳の章)。橋下部でponsに分布しない繊維が集合して延髄の錐体を作る。
脳幹経過中に脳神経の運動(体性)核を支配すべく繊維を与える。これらの繊維のあるものは正中部を超えて交叉するが、残るものもある。このことは実際的に意味で言及の要がある。このことは、脳神経核の(運動性)あるものは両側性大脳支配なるこを意味する。この結論は臨床観察より。
完全に交叉性のものは、舌下神経核
(個々の異変のあることを許して一般的に云えば)副神経核、顔面神経核の一部(下部顔面に関係を支配する部分)
ある個体では、三叉神経運動核も然り。
大多数の繊維が錐体交叉lateral corticospinal tracts(後角の外側域)(2-9図、1-2図をみよ)ある研究者は、Foersterに反対して、外側皮質脊髄路には体部位局在の配列はみられぬという。
非交叉性のものは、(小部分で%は異変あり)。ventral cortex spinal tracts(前正中裂のそば)前索内、この腹側皮質脊髄路は恐らく胸髄以下に至るものは稀。
加え、しかしながら、若干の非交叉性の繊維は外側皮質脊髄路に加わる。
カハールによりずっと指摘されている所であるが、全部ではないか、数多くの皮質脊髄繊維は経定中に豊富な側枝collateralsを出す。→線条体、視床核、赤核、橋核、網様体これは生理の研究でも知られている所である。皮質からの直接繊維もある。脊髄内でもたくさんの側枝を与える。生理の(antidromic)仕事では頸髄に至る皮質脊髄繊維の30%がspinal cord下位レベルにも側枝を与えている(Shinoda, Arnold, Asanuma, 1976).
古典的“運動野”である4野のみが皮質脊髄路の源泉ではない。ヒト、サルで6野にも+が、6野よりも前方の領域からも+であるか決定的結論は出されていない。
ネコではgyrus proreus (primate 8-12野に相当)から+(Nauta法、Nyberg-Hansen 1969b)3.1.2野(一次感覚野)はすべての動物で錐体路の重要な源である。更に頭頂葉(5,野)も脊髄に繊維を送る。少なくともネコでは第2次(第2)体性感覚野からも+、又ネコで内側面皮質の補足運動野も(+)。側頭葉、喉頭葉からはWalberg と Brodal (Glees法)で少しあるがNauta法(Nyberg-Hansen, 1969b)でみられない。
サルでも同じ様
HRPで・・・
5、7野からはサルの方がネコよりcorticospinal fibers多いが、一方では前運動野premotor areaからは送らしい。
HRP(サル)
Betzを含む大型細胞もより小さい錐体細胞もorigin (Coulter, Ewing, Carter (1976, 他)すべての第1知覚運動皮質から。
Jones と Wise (1977) 数ヶの房状に集まって配列した細胞がgapに隔てられる傾向。V層の深部のみ。
錐体路繊維の径は大きなばらつきがありしたがって伝導速度も然り、大多数は細いものLassek (1942b) によれば(ヒトの錐体)
90%・・・直径1~4 μm
1.73%≧直径11~22 μm
60%がヒトでは有髄(Lassek, 1942b)
94% (DeMyer, 1959)
大経有髄繊維の数は4野のBetz細胞の推定数と大体一致する。注目すべきは4野を破壊すると殆どすべての大径繊維が消失する(Haggqvist, 1937)と軸索の長さとの間に関連があるという一般の推定(assumption)を支持する。
ばらつきが大きくネコで7-70m/秒。この内14 m/secと42 m/secとに2つのピークがある。これをもとに生理学上時々”fast”(中間帯よりventralを支配)および”slow”(もっとdorsalを支配らしい機能的には、感覚のインパルスの中枢伝導と脊髄反射に影響を与える。)皮質脊髄繊維の区別がある。
皮質脊髄繊維の終止
ネコ(鍍銀法)あったっとしても極く僅かしか大型運動ニューロンに接触しない。“中間帯”と後角の基部終わる。後日確認もされ、生理学上もcorticospinal fibersインパルスは介在ニューロンより運動ニューロンをactivateする(Lloyd, 1941)。詳しくは、IV-VII層(主にV-VI)of Rexed (1952, 1954)に終わる[Nyberg-Hansen と Brodal 1963]。IXまでは1本も繊維を追跡し得ず。cp. Scheibel: 1966b (4-4図)
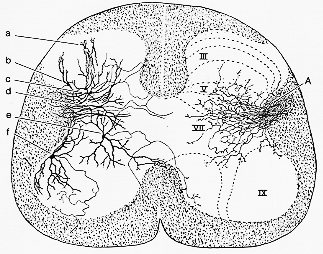
FIG. 4-4 Synaptic presentation of findings made in Golgi sections of he cat's spinal cord. Rexed's kaminae are indicated. On the right side are seen corticospinal fibers and collaterals (A) entering the gray matter in a fan-shaped pattern, mainly within laminae V-VII. On the left side various types of cells in the gray matter are seen. These may be contacted by terminals of corticospinal fibers. Some cells (f) send their axon into lamina IX, harboring motoneurons. Note that some (e) send their axon across the midline. Interneurons. c, d, and e are situated within the terminal area of corticospinal fibers. From Scheibel and Scheibel (1966b).
中心前回(運)から太”fast” 主にVI, VII
中心後回(知)から細”fast” 主にIV, V (VI少) ネコ、サルでも生理学的にも
Coulter と Jones (1977) サル、ARG. 3a, 4の一部→運動細胞群のある領野に終わる。
3.1.2間でも異なる。
重複終止域があるが、区別がある。
運動補助野からの終止域は第一(次)知覚運動野からと同じ(Nyberg-Hansen, 1969a)
ネコでは”central region”からの皮質脊髄繊維は電気生理学的にも決着づいたように(Lloyd, 1941; Hern, Philips と Porter, 1962;他)介在ニューロンを介してのみmotoneuronを活動化させるが、サルでは若干のものが、直接motoneuronに終わる(Hoff と Hoff, 1934; Kuypers, 1960; Liu と Chambers 1964)生理学的にも(polysynaptic)の他に単シナプス性の証拠がある。ヒトではよく知られていないが、若干が終わる。
Kuypers と Brinkman (1970) precentral cortex中心前回からのcorticospinal fiber詳しい。
体性及び内臓性の遠心性脳神経核は、ネコ→その付近に終わり核内には終わらぬがサルでは核内に終わるものもあり(Kuypers, 1960)
一般に皮質脊髄繊維は、屈筋支配の運動ニューロンに対しては促進性作用(facilitatory action)をするが、伸筋の運動ニューロンに対しては通常抑制性である(文献略)。更にγ-ニューロンに対する作用も同様に思われる。
ネコ V-VII(4-4図をみよ)にある介在ニューロン→α,γ-neuron.
ある種の体性脊髄反射の反射弓に組み込まれている。介在ニューロンはcorticospinal fiberのインパルスで促進される如し(Lundberg, Norsell と Voorhoeve, 1962)。運動ニューロンへの皮質脊髄繊維の興奮性効果はdisynapticallyに起こる。そしてこの際の介在ニューロンは一次spindle afferents (Ia)から、下行繊維系からも共に影響されるが如し(Illert, Lundberg, と Tanaka, 1975)。これらの介在ニューロンは重要な統合機能を持っているように思われる。
サル 単シナプス結合はとくに関心をよぶ。Phillips と Porter (1977)の本。
corticospinal fibers 単シナプス性に運動ニューロンを興奮させる。
一方抑制は介在ニューロンと介して=シナプス(disynaptically)性に起こるようだ。
介在ニューロン(一部)
皮質脊髄路といくつかの共通した特徴がある。赤核は新しい脳で黒質の赤帯のように幾分赤みがっかてみえる。中脳の上級レベルにあり殆ど球形。鉄を多く含み求心性および遠心性の繊維で構成されている一種のcapsule包みで被われている。動眼神経の多くの繊維が横切る(4-5図)。繊維結合からも、細胞構成からも赤核は1つのentityではない。習慣上、尾部の大細胞性部分(大型多極性細胞を含む)と吻側部の小細胞(性)部分(種々の小型細胞を有する)に区別される。
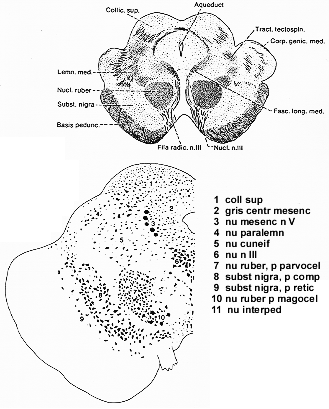
FIG. 4-5 Drawing above of a transverse section through the mesencephalon in man. The red nucleus is traversed by root fibers of the oculomotor nerve. Left, a diagram (from Nieuwenhuys, Voogd, and van Huijzen, 1978) of the main cellular groups at a level of the brainstem corresponding approximately to the level shown above.
この区分はヒトのものから出ずるが如し、ヒトでは大細胞は最も尾方部のみにある。そこで小細胞を混ず、系統発生的には大細胞の比較的な占める割合は減少する。動物の赤核には“小細胞性”“大細胞性”という名称が適用されているのは不幸なことである。何故ならたとえばネコでは、大型、小型の細胞の出現に関して云えば、全く徐々に漸次移行して(caudalからrostralへと)いる。サルでは大型細胞は尾部1/3部のみ-小型も混じっている-(Kuypers と Lawrence, 1967; Miller and Strominger, 1973)。
赤核にはafferent efferent多いがいくつか考えてみよう。まづ、赤核脊髄路(しばしばMonakow’s bundle, モナコフ束といわれる)。ヒトの知識は少ないので動物所見とする。
鍍銀法でネコ、サル、ラット、オポサム腰仙髄レベルまで至る。Edwards (1972) ARG法、ネコ、ほとんどすべてが交叉性、赤核を出て直後に交叉する。脊髄では赤核脊髄路は外側皮質脊髄路のすぐ腹側でやや外側にある(4-5図)。
大・小細胞共にorigin(ネコ、サル)。繊維の太さも種々、伝導速度も31-120 m/sec. HRPでもこの赤核脊髄路はサルで赤核全長に起始あることを示した(Kneisley, Biber と LaVail, 1978)。ネコも然り。局在あり。
脊髄での終止域V,VI,VII層、IX層には(-)、大体分銅皮質域からの皮質脊髄繊維終止域と一致する。axodendritic contact.
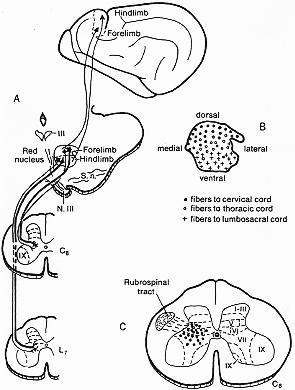
FIG. 4-6 Diagram showing the principal features in the cortico-rubro-spinal pathway as determined experimentally in the cat. A: The corticorubral fibers take or igin in the anterior sigmoid gyrus, the "motor cortex" (above), and end in a somatotopical pattern in the red nucleus. The somatotopic arrangement is upheld in the rubropinal projectionk, coming from small as well as large cells in the red nucleus. B: The somatotopic pattern in the red nucleus as seen in a transverse section at middle levels of the nucleus. C: The sites of termination of rubrospinal fibers are restricted to Rexed's laminae V to VII. Based on findings of Pompeiano and Brodal (1957c), Rinvik and Walberg (1963), and Nyberg-Hansen and Brodal (1964).
電気生理
赤核刺激は対側の屈曲の運動ニューロンをactivateする(細胞内にEPSP記録される)。一方、対側の伸筋支配運動ニューロンIPSP(抑制)、(Sasaki, et al. 1960)介在ニューロンを介す(Hongo, Jankowska, Lundberg, 1965)。Hongoらは、VIとVII層の介在ニューロンに終わり,このニューロンは赤核脊髄路によって促進される。
Granit と Holmgren (1955)が最初に示唆したように、この路はαニューロンばかりでなくγ―ニューロンにも影響を与える。Appelberg と Kosary (1963)によれば、屈筋の静的(static)γ―運動ニューロン(3章をみよ)は促進され、伸筋のそれは抑制される。
(マトメ)赤核脊髄路
機能、サル、ネコ恐らく同じ、体性局在的に構成。
α―運動ニューロンと静的γニューロン(屈筋の)を興奮させる。
赤核の生理に関する総説としてMassion, 1976を参照
赤核からの遠心性繊維(支配)-脊髄以外に-
外側網様核、下オリーブ核、外側楔状核-これらは小脳へ繊維を送る-
若干の赤核からの遠心繊維は直接小脳へ行く。(NIAへのみ、体性部位局在あり)故に相互結合(reciprocal)尾方2/3のみらしい。
cp. NIA→赤核 主として細胞体と近位樹状突起に終わる(ネコ、ウサギ)
NIA刺激、単シナプス性にEPSPたくさんの軸索が単一赤核ニューロンにconverge
他に顔面神経核、(sensory)三叉神経核、薄側核、楔状束核、前庭神経核群の一部に終わる。赤核遠心繊維中、脊髄と小脳へ行く以外のものの起始部は前方部約1/3に殆ど限られるようである。
赤核視床の上行路があるのか否かは、小脳・視床路が赤核内に通るので決定が困難だったが、ないということになった。Hopkinsと Lawrence (1975)上小脳脚を両側破壊したサルを1年生存させた後、赤核傷害で視床に変性をみなかった。
Edwards (1972) ARGで(-)
Anderson (1971)生理学的に証明し得ず。
赤核への求心性結合
赤核の機能的役割の分布
赤核の多くの細胞が2分岐(dichotomizing)軸索をもっており(cajal)、それが2つの異なる標的を有しているだろうと云うことは興味を引く。赤核の逆行変化から(Brodal と Gogstad, 1954) 赤核の脊髄に行く繊維と小脳に行く繊維は広範囲に(to a large extent)同じ赤核細胞から出ると結論された。生理学的にもAnderson (1971)がこれを確認し、更に若干の細胞は脊髄と延髄の核(恐らく下オリーブ核)を同じく支配するものがあることが示された。後者の繊維は非交叉で他の赤核遠心繊維と対照的であり、後に形態的にも結論が下された(5章をみよ)。機能的観点から小脳求心繊維と比較して大脳求心繊維は数がはるかに少ないことは興味ある。
生理学的研究からも、これに一致して赤核への小脳性インプットの方がより強力なことが示された。以上の論考より、赤核は多くの源泉から影響をうけ、特に脊髄を小脳に対して働きうると云えよう。運動のメカニズムに関連している。赤核と小脳中間帯との共同作業collaborationの機能また、そのフィードバック系、赤核は間接的皮質脊髄路の一中継点也。その構成は直接の皮質脊髄路の“中心前回”成分と著しい類似がみられる。両繊維系は大体同じ皮質から起こる。径がまばらなものを包む。脊髄全長に終わる。局在(体性)パタンあり。終止域が大変よく似ている(図4-3)。両繊維系と屈筋αニューロンとγ運動ニューロンを優勢的に促進させ。同じ介在ニューロンを単シナプス性に興奮させる。
| 前庭脊髄路 |
前庭核を出て繊維は腹方向に走り、後方にまがり、同側の前側索(脊髄部で)を下行する。この古典的な前庭脊髄路の他に、内側縦束の中を下行するより繊維数の少ない構成のもう一つの道がある。脊髄内では、これらの繊維は正中付近で前索中に、いわゆるsulcomarginal fasciculus中に両側性にみられる。Nyberg-Hansen (1966a):これを内側前庭脊髄路、とし、前者(古典的なもの)を外側前庭脊髄路とする。ネコで研究。
外側前庭脊髄路
起死核は外側核(ダイテルスDeiters)で終止は腰仙髄レベルまで巨大細胞のみならず小型の細胞もPompeiano と Brodal, 1957a (mod. Gudden法)ネコ太い繊維も細い繊維もPeterson と Coulter, 1977(HPR法)ネコ、核に体性部位局在パタンがみられる(4-7図)。ネコサル殆ど同じ
頸・前肢域 外側核の背尾部
体幹域
後肢域 前腹部
この終止域はcorticospinal, rubrospinalと異なる。VIII層とVII層の腹方、中心部。とくに大きい樹状突起(large dendrites)に、IX層にまで至るものはごく僅かVIII層にある細胞の多くは軸索を正中をこえて対側に送るので、このことから、外側前庭核を刺激したときみられる両側性効果を説明できよう。
Deiters核と外側前庭脊髄路は同側四肢の伸筋(展)緊張extensor tonusを増大させる。(Pompeiano, 1972aの総説をみよ)。Deiters核の前方で除脳とたときの緊張の増大(除脳強直decerebrate rigidity, 後で論ずる)。脊髄に対するDeiters核の対立するもののなくなった状態の行為actionに大部分よっている。そしてこの状態は核が破壊されると消失してしまう。これに一致するものとして、微小電極法でネコ外側核を刺激→伸筋支配の運動ニューロンのみにEPSP (Lund と Pompeiano, 1965). 屈筋支配の運動ニューロンのみにIPSP
興奮性効果は単シナプス性と結論されるが、IX層まで至る繊維が極少故にIX層のmotoneuronのVIII~VIIまで拡がった樹状突起末端に多くの数の結合があるのであろう。
皮質脊髄路、赤核脊髄路と同様に、外側前庭脊髄路もα-motoneuronのみでなくγneuronにもactする。γ-neuronへの行動は興奮性という。
後日の研究で、より複雑なことが知られた。Deiters核への求心路として大脳皮質はないらしい、豊富な投射を小脳の脊髄部皮質皮質、“前庭小脳”および室頂核から繊維を受ける。核内の特定の終止域がある(7-10図、p.474)。ここでは小脳からの求心路が外側核の機能に重要な役割を演ずることを強調して次章にゆづる。
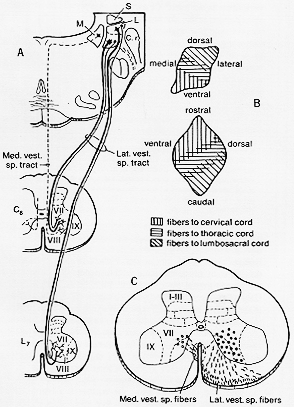
FIG. 4-7 Diagram showing the principal fetures in the vestibulospinal pathways as determined experimentally in the cat. A: The lateral vestibulospinal tract comes only from the lateral vestibular nucleus of Deiters (L) but from small as well as large cells. It is somatotopically arranged. The medial vestibulospinal tract is derived from rthe medial vestibular nucleus (M) and does not reach the lumbar cord. B: The somatotopic pattern in a sagittal reconstruction of the nucleus (below). C: The vestibulospinal tracts terminate in Rexed's laminae VII and VIII. Based on findings of Pompeiano and Brodal (1957a), Nyberg-Hansen and Mascitti (1964) and Nyberg-Hansen (1964).
他の内側前庭脊髄路 modestで外側路に比してよく知られていない。(4-7図)
内側核のみから起こる。終止域は外側路の終止野とほぼ同じ内側路と路は、前庭インパルスを頸部と前肢に主として伝え、外側核と路は全身に影響を与える。多少興味あるのは内側および外側核が前庭装置の異なる部位からおこる第1次前庭神経により支配されているらしいこと(7章をみよ)。
内側核も小脳から繊維をうけるが、大脳皮質からはうけない(中脳からの求心路がある)。
WilsonとYoshida(1969b):内側からの下行枝頸髄上部の運動ニューロンに単シナップス結合をし(しかし(前)四肢の運動ニューロンではない)抑制性である。
| 網様体脊髄路 |
網様体に関する章(7章)で述べられるが、これは散在性の細胞や繊維の集合ではない。いくつかの多少とも区分された核に分かれ、求心繊維、遠心繊維が一部異なり、恐らく、従って、機能上も異なるものであろう。網様体脊髄路に関して云うと、起始部が異なる2つのグループに分けられよう。Torvik と Brodal (1967)modifiedグッデン法。
大小のcells、橋、延髄のすべてのレベルに分布。しかし明らかに2つのmaximal areas (起始)がある。一つは橋に、一つは延髄に、巨大細胞が現れるreticular formationの2/3にある。p.402のFig.6-4をみよ。Ponsからは同側性下行、Medullaからは両側性中脳reticular formationからはない(Nyberg-Hansen,1965a, Edwards 1975)この投射には体性局在のパタンはみられない。異なるレベルに分枝を与えてRMに終わる。(生理の仕事、Petersonら、1975)。異なるレベルの活動のcoordination調整(整合、連絡)に重要であるというなるべし、
Pons reticular formationからの終止域(○より腹側に終わる)主にVIII
Medullary reticular formationからの終止域主にVII
α、γ共に影響を与える。(恐らく介在ニューロンを介して)他の下行路同様
分岐している。(中枢性感覚伝導、脊髄反射)にも影響を与える。
| 脊髄への他の下行路 |
cortex, ruber, vestibular, reticularの他脊髄は他の灰白質部から直接の下行路をうける。
中心灰白質の腹側で交叉、medial longitudinal fasciculusのすぐ腹側をreticular formation中を下行する。上丘のみ、大多数が上部頸髄に終わる。下行頸髄にまで至るものはごく少ない。(Nyberg-Hansen, 1964b)。主にVIIとVIに終わる。上丘を電気刺激すると対側へ目と、頭を向ける。この効果は少なくとも一部は視蓋脊髄路を介する。Andersonら(1971)上行刺激
対側頸部運動ニューロンに短潜時のEPSP
同側頸部運動ニューロンにEPSPとIPSPのcombinationを観察した。
| 間質核(カハール)脊髄路 |
カハール核はrostral中脳に中心灰白質のすぐ腹側にmedial longitudinal fasciculus内縦束にへだてられて在る。脊髄への下行は内縦束内を通り主として同側性、仙髄レベルまで主にVIII層、IXは(-)。終止域は前庭脊髄繊維に似る。求心繊維は多くから来るがとりわけ前庭神経核(7章をみよ)大脳皮質からもあると? 頭身体の縦軸の周りを回転すること(電気刺激でみられる)。又、視覚性および前庭インパルスに反応して頸と身体(体幹)の筋肉に効果を伝える重要な中継核か?(Hyde と Eason, 1959をみよ)。自律系の効果も。
| 弧束核脊髄路 |
弱い投射あり。Loewy と Burton (1978)HRPとARGネコ。
solitariospinal→C4-6の横隔神経の運動神経核域Thalamus前角、intermediolateral column側柱へ。同側優位。
その主たる機能は内臓性感覚を上位レベルに伝えるにある(7章)とはいうものの、また脊髄(性)のメカニズム(体性及び内臓性)にも明らかに影響する。Vagusやglossopharyngealから入ってくる内臓性インパルスに反対して脊髄をactivateする上にこの繊維は恐らく重要也、大脳皮質から求心性繊維をうける。(精神的インパルスが興奮である又は抑制であれ、内臓性反射に影響するというよく知られた事実を説明しようとするときrelevant也。小脳から脊髄への投射は疑われてきたがfastigiospinal路は認められた。弱い、頸髄支配、縫線核から脊髄への投射(主にnucleus raphe magnusから、6章をみよ-raphe)。RM後側索を下行する。serotonergic.
青班核からの下行路noradrenergic同側前索、腰髄まで下オリーブ核からRMへの投射は恐らく存在せず(多くのテキストにあるが)ヒトHelweg’s三稜(角)路
3角帯が前索にあるが、(上部頸髄部)olivospinalというが全く推測の域を出ない。
| 基底核といくつかの関連した核群 |
上述の如く、たくさんの下行線維束が脊髄に至り影響を与える。大脳皮質は直接的に又間接的に(赤核、網様体の一部、上丘、弧束核、縫線核の一部)影響を与える。更に大脳から小脳、それから前庭神経核や網様体との結合を介した遠回しの回路もある。
他の神経路もある。つまり大脳基底核basal gangliaを通るものを介して。この15年間基底核とその結合はよく調べられ関心をもたれて来。大きい特徴のある脳の部分故、少し考察を加えたい。以前から大脳から脊髄へ導く経過にある重要な中継点と思われてきたが、(つまり、一義的に“運動性”機能をもつと)。最初の研究結果はこの考え方の改定を必要とされている。
時代の経過と共に”basal ganglia”は(種々の)異なった(言外に含んだ)意味合いをもっていた。昔の解剖学者は、脳内の視床も包んだすべての大きな核に対する。一般的名称として使用した。脳の発達がよく理解されてからは、視床が除外され、たとえば扁桃体が加えられた。今日まですべての研究者は尾状核とレンズ核(被殻と淡蒼球)を主たる部分(main mass)と考えているものの、どの構造物を包含すべきかという一般に承認された定義をもっていない。前障は通常含められるが、一方、扁桃体とその大ざっぱに云って(大きく)異なる結合と機能の故にしばしば除かれる。通例として、basal gangliaとの関連で視床下核subthalamic nucleusと黒質が考察される。これは以下のような意味合いにおいてなされる。線条体(striate body or corpus striatum)という術語は基底核と殆ど同義語としてしばしば使用されており、前障、尾状核、被殻、および淡蒼球を包含する。この名前は、多数の有髄線維束が細胞集団を貫いており“線条”の外観を与える。髄鞘染色標本のappearanceに言い及んだものである。
| 線条体 |
半球の髄鞘層(白質層)中、繊維条strandによりいくつかに区分、(4-9,4-10図)。
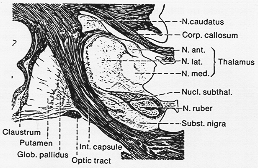
FIG. 4-10 Drawing of a myelin-sheath-stained frontal section through the human brain, showing the basal ganglia.
前障 島の下(最外側部)。thin sheet薄い板の灰白質
最外包capsula extremaにより皮質と隔てられ被殻には外包capsula extermaで、前障は島域の皮質由来の如くでだから発生上厳密に線条体には属さず。
固有の線条体(尾状核-レンズ核)は論ずべき特徴あり、系統発生学的にも微細構造上も、尾状核と被殻は似ており、共に、しかし、淡蒼球とは異なる。(globus pallidus, palladium)
淡蒼球はレンズ核の内側の部分でpale color(尾状核、被殻とくばべ)でこの名あり。
外節(anterior lateral)
内節(posterior medial) primitive neural tube
尾状核や被殻に比し系統発生的に古く(間脳内に生じる)原始神経管の基板から発生する。
尾状核と被殻は、これに反して(他方)、系統発生上、遅れて、終脳(telencephalon)から発生し、大脳皮質の発達に相応して大きくなる。下等な動物ではcaudatusとPutamenはは内包によってハッキリと分離されていない。
ヒトでは内包が発達し、皮質遠心性(e.g.錐体路)と皮質求心性繊維に富む。caudatusとPutamen間に橋渡しの条(strands)淡蒼球はいわゆるpaleostriatum, 尾状核と被殻はneostriatum (単にstriatum)(扁桃体はarchistriatum)
細胞構造をみても
淡蒼球は大型の主として紡錘型細胞がむしろまばらにあり、striatum (尾状核と被殻)は、密に小型多性細胞がみられ、その内に大型の多極性細胞あり、しかしくわしくしらべてみたところ、cytology is far more complex.
淡蒼球は、尾方に黒質の網様部pars reticulateの前部と連結している(下述)。
この2つはneuropilの微細構成が非常によく似ている。おそらく両者ともstriatumからたくさんの繊維をうけているという事実に関連がある。(kemp, 1970; Fox and Rafols, 1976)淡蒼球(pallidum)は、丁度黒質や赤核と同じように、組織化学的に同定されうる。大量の鉄を含んでいる。血管床も(線条体の如く密でなく)異なる。この構造上のstriatumとpallidumとの差はこれらの核の疾患の時に見れる症候の差で、とりわけ、示される機能的相違に対応する。
| 視床下核(Luys体) |
間脳のbasal part(中脳との移行部)に位置し、卵形ovoidの核、尾方に腹方に、黒質とつづく。
ヒトでよく発達している。内眼で暗黒く弓状の帯、大脳脚内に両側にみえる。細胞内密に詰まった色素顆粒(メラニン)、このdark zone(ゆわゆる緻密部pars compacta)の腹側に特、黒質はより散在性の細胞よりなり、それが一部腹側の方に向かって大脳脚中に侵入しているところの一つの境界帯式を作る。黒質のこの部分は(いわゆる網様部part reticulata)内眼でも黒い帯の腹側部に区別し得る。淡蒼球や赤核と同様黒質は鉄を一定、とくにglia細胞内に含んでいる。緻密部の多数の細胞がその樹状突起を腹方に網様部に向かって送っていることは特別に興味あることである。電子顕微鏡の研究で、このdendritesは神経終末に密に包まれている。その結果、緻密部neuropilが微細構造に淡蒼球とすごく似た形を示す。この類似点の他にこの2つの核は病理学的状態で相応して変化をしばしば示す。
| 基底核と関連核の繊維結合 |
われわれの知識はいまだにむしろ不完全である(この主題に関して)。
確証は、顕著な黒質:線条体ドーパミン投射と縫線背核dorsal nucleus of the rapheからのセロトニン結合
ARG, HRP法正確で詳細な構成
以前考えられた以上に結合があり、核間の結合は以前信じられていたよりも(より)特異的specific一般に種々の亜核は豊富に相互に結合されている。
大きい求心結合は大脳皮質から尾状核と被殻への繊維の如し、淡蒼球内節と黒質のみが固有大脳基底核以外の部位に相当程度(量)の投射をするように思われる。主な結合のいくつかをFig4-11に示した。
大脳皮質からの投射:すべての皮質からといってよい。尾状核と被殻へ局在性あり、重複あるが(前後内外も)側頭葉が霊長類で発達しるために動物差がみられる(局在性に)。しかしARG以後、4野→両側性に被殻へ3.1.2野→厳密に同側性に被殻へ9野は+らしいが尾状核投射は無視できるほど少ない。
以上のARGの仕事からみて、尾状核と被殻を一つの機能上単位と見倣し得ず。又、Yeterian と Van Hoesen (1978) Brain Researchサル 連合野→尾状核内で1カ所以上に。皮質間相互結合のあるつの部位から尾状核の同一部位に終わる。尾状核を“運動”か“知覚”かのみの機能を持つと考えるとに注意を喚起した(Teuber 1976)。単一独立の皮質、線状体ニューロンか又は皮質脊髄路などのaxonか、解決していないが独立らしい。V層、(Wise と Jones, 1976, Jones, Coulter, Burton, と Porter 1977)。
黒質からの投射:
大きな源泉也。大部分が緻密部の細胞が起始。(pars compacta)。少数が網様部からとventral と ventrolateral被蓋域にある黒質近傍細胞群から起こる。この繊維は局在的構成黒質の背内縁部に沿って上行し、視床下部へ、そして内包の内側部に入り、それから背吻側方向に走り、尾状核の頭部と被殻の前部に至る。線状体のもっと後部に行く繊維はForel野で主たる神経束から離れ外側を走り、視床下核の背側を通り、内包の後部と淡蒼球を貫いて走る。ドーパミンを含む。線状体ドーパミンの主たる源である。この繊維系の変性はパーキンソン病の病因上重要な面と考えられている。ドーパミン合成の前駆物質(l-Dopa)-l-dihydroxyphenylamine治療。しかしながら近年CNS中のドーパミン病以外の種々の疾患について関連している。
分裂病治療に用いる薬物は脳内においてドーパミンantagonist(拮抗薬)である(総説としてMatthysee, 1973; Snyder, Banerjee, Yamamura と Geenberg, 1974; Iversen, 1975をみよ)という知覚はとくに興味あるものである。dopaminergic路は脳内にもある故、ドーパミン受容体のブロック阻止(後)に観察される抗精神薬効果にとってどのドーパミン経路がcrucialカギなのか、明らかになるにほど遠い。
視床からの投射:
第3番目の重要な線条体求心繊維の成分は視床からのもの也。髄板内核群から、局在性あり。
centralis lateralis CL
centralis medialis CM
paracentralis →線条体の前部へ
centrum medianum CM
Parafascicularis PF →線条体の後部へ
cp. 新見教科書。 中心内側核Nucleus medialis centralis (中心正中核Nucleus centromedianus)視床線条体路同様patchyの終止。
CM→尾状核、被殻(ネコ) Royce (1978a)
CM→被殻のみ(サル) Kalil (1978) 種による差かどうか?
縫線核からの投射
第4番に量的に重要な源はraphe nucleiと思われる。
Dorsal nucleus of the rapheこの核を破壊すると線条体セロトニン量↓と合成酵素のtryptophan hydroxylase↓。脳内で最も高いセロトニン濃度が線条体にみられるが、主にstriatumの腹尾域であるようだ。この系は抑制系。しかしこの系の(線条体へのセロトニン入力の機能的意義については実
線条体を均一なる構造物と考えてはならぬ。このように線条体は少なくとも4つの主な求心繊維成分を受けるが、よく記載されている尾状核と被殻への遠心性投射は淡蒼球へ行くものと黒質へ行くものとである。
線条体遠心繊維は非常に細く(平均0.6μm)乏髄poorly myelinatedである。小束としてWilsonの”pencil”として集まり淡蒼球に集まりそれからこの核内を又沿っていく。尾方へは大脳脚をEdingerの”comb system”の腹側成分として横切り、それから、黒質に至る。GolgiおよびEMの研究で、線条体淡蒼球繊維は、黒質へ行く繊維(軸索)の側枝であることが示された(Fox and Rafols, と Cowan 1975)。線条体遠心路には正確な部位局在がある。(サル、ラット、ネコも同類)。
尾状核→淡蒼球 (内・外の両節)の背側
被殻の後部→最も外側
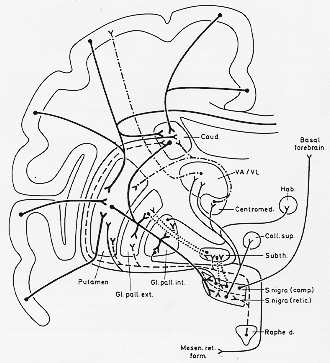
FIG. 4-11 Diagram of some of the main connections of the basal ganglia. For details see text.
黒質はもう一方の線条体からの終止野であるこの10年前黒質求心系の多くの仕事があり図4-11,4-12にまとめた。内包の後肢と大脳脚中を通過中に(striatumの)腹内部と最吻側から出た繊維は、より尾方;背側域から出た繊維により次第に背側方におしやられる線条体黒質繊維は緻密部と網様部へ共に分布する。明瞭な内・外の部位局在をもつ、尾状核頭部の腹内側→黒質の最も内側、被殻の後部→最も多側。
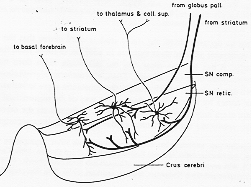
FIG. 4-12 Diagram illustrating the principal features in the organization of the substantia nigra. Note that the dendrites of many cells in the pars compacta (SN comp.) extend into the pars reticulata (SN retic.). For detalis see text and Fig. 4-11.
変性鍍銀法では通過繊維のため決定がむずかしかったが、ARG. HRPにより存在が確定した。更にネコでGrofova 1975,1979)はentopeduncular核(脳脚内核)(淡蒼球内節に相当)からも緻密部に(網様部にはない)弱い投射を証明した。
| 他の黒質求心繊維の起始核として |
縫線核背核,扁桃体の中心核、分界条(stria terminalis)のbed nucleus他、側坐核(nucleus accumbens septi)からBunney と Aghajanian (1976a), Swanson と Cowan (1975)-ARG. しかし他の研究者(HRPで)negative.
この所見の違いの説明は恐らく、Tsaiの腹側被蓋野(VTA)および黒質緻密質内側部にあるドーパミン細胞を区別することがむずかしいことであろう。他方、しかしながら、VTAから側坐核への投射は確立されるが如し(Bjorklund と Lindvall, 1978, for reference)
皮質黒質路、一つの論争点であるが
変性法(Rinvik, 1966), EM(Rinvik, と Walberg, 1966)は疑問としたがHRP法で何人かの研究者は、前頭前野(少数の散在性に)→黒質
ARG(Kunzle, 1978) 6.9野→緻密部(サル)。patchy分布。
第3番目の、最近まで予期されなかった源は、恐らく(+)のちに確証。
視床下核 subthalamic nucleus
最近Nauta と Cole (1978)ARGで多量にあることをサル、ネコで認めた。上述の多くの研究から黒質は新線条体からの投射に加えていくかの求心繊維系があることが明瞭たるべし。同様な黒質はその繊維を線条体にばかりおくっているのではないことも明白となった。すなわち、顕著な黒質視床投射が証明された。このことは黒質が視床を介して大脳運動皮質に影響を与えることを可能ならしめる。しかしながら、黒質線条体により(介して)、黒質-線条体-淡蒼球-視床-皮質のルートを介して同様の影響を大脳皮質に対して与えることができよう。専らでにとしても、主として黒質網様部の細胞から黒質・視床投射がおこるようだ。終止はVMとVLの内側部の一部。VLからは(2章をみよ)。大脳運動領へ投射がしられている。
両側性(同側性優位)恐らく網様部のみから出て、上丘の中間~深層にYork と Faber (1977)の生理学的仕事によれば視覚性定位行動に関して重要であろう。Bentivoglio, van der Kooy と Kuypers (1979)逆行性蛍光二重標識法によれば網様部の大多数が上丘か又は視床に投射する。しかし、可成りの細胞が両方に繊維を送る(dichotomizing axons) 。同研究で彼らは黒質・線条体繊維には分岐軸索はみられなかった。
以上に加えるにHRP法で最近で弱い投射が次に見られた。
黒質→下部脳幹reticular formation (Rinvik, Grofova と Ottersen, 1976)
→中心灰白質(Hopkins と Niessen, 1976; Grofova, Ottersen と Rinvik, 1978)
特に興味あるのは黒質の緻密部から線条体へ行くものばかりでなく。他にolfactory tubercle (嗅結部)、扁桃体のようなbasal forebrain structuresへ更に皮質前頭葉にさえ行くものがあるという研究である。これら脳底前脳部(basal forebrain structure)への黒質遠心性投射はドーパミン性である。これらが黒質線条体繊維(ドーパミン性の)の分岐かどうか定められていない。Fallonら(1978)は新線条体へとallocortexへ投射するnigra緻密部でドーパミンを含む細胞が分離集団を作っていることを示した。
脳内のいわゆるドーパミン経路が多く出版させた。この投射についての大きな関心はとくに分裂病治療に使用する薬物は脳内でドーパミン拮抗薬dopamine antagonistとして動く(作用するact)という観察に由来する。黒質線条体投射はCNS内で最大の-少なくも一番よく調べられている-ドーパミン経路であろうが、この数年のいくつかのlinesが明らかに示すところで、中脳におけるドーパミン含有細胞は黒質の緻密部にのみ局在しているのではない。この部から連続して内側にあるTsaiのventral tegmental areaへnigraからつづいており、背側へは中脳reticular formationの腹側部にドーパミンを多量に含んだ細胞が散在している。(最近の総説としてBjorklund と Lindvall, 1978をみよ)。このニューロン帯(band)の内の細胞ははっきりしたdistinctな遠心性および求心性の繊維結合を持っている。この事実は心に銘記すべきである。脳機能中でのドーパミンの役割に関して一般化することに対する警告(Warningとして役立つ。)である。種々の求心遠心結合により、一目瞭然、黒質は様々の高次脳機能に多方向の活躍をする。ドーパミン以外の伝達物質候補に注目。
striofugal…GABA
(nigral)…substance P 異なる線条体ニューロンから?
striatofugal fibers →GABA→
共に(単シナプス性に黒質視床細胞黒質上丘細胞にがドーパミン性黒質線条体細胞に単シナプス性につくかは今はお、論じれている。(deleted)議論のあるところ)
pallidonigral fibers→GABA→
ドーパミン性黒質線条体ニューロンは直接尾状核の黒質への投射のニューロンに接触するのではなく(Kitai et al.)線条体の介在ニューロンにつく(McGreerら、1975)。
黒質視蓋ニューロンは恐らくGABAergic
視床下核の繊維結合(4-11図をみよ)はとくにむづかしかった。
非常に最近まで確証さえていたものは淡蒼球外節から視床下核へあるだろうということ(Ransonら1941)(他に)。この淡蒼球、視床下核、投射には部位局在がみられ、GABA性らしい。この他の視床下核求心性投射として、originがHRPで(ネコ、サルで)。
縫線核背核 dorsal raphe nucleus
青斑核
黒質緻密部 (これは相互性結合!!)
(大脳)脚橋核pedunculopontine nucleus
上述の如く淡蒼球は内節と外節とから成っている。
ARG(サル、ネコ)。Nauta と Cole (1978)は視床下核は以前一般に考えれていたように内節へだけでなく。内・外両節へ投射することを示した。淡蒼球の内節は、黒質の網様部と共に、基底核固有部以外に繊維を送る基底核(群)の唯一の部分である。これらの淡蒼球より出る遠心性繊維は2つのはっきりした束に分かれている。
一つはansa lenticularis レンズ核(係蹄)
他はfasciculus lenticularis レンズ核(索or束) 内包に対し(関し)異なるコースをとる。
レンズ核蹄は主に淡蒼球内節の外側部から起こる。この繊維は淡蒼球の腹側縁に沿って走り、内包の後肢のまわりをかすめて通り、ForelのH野に入る。レンズ核束の方は、内節の背内側より起こり、内包内を数ヶの小さい束を作って通り抜ける。レンズ核蹄は及び束を作って通り抜ける。レンズ核係蹄及び束は、Forel H野の中で合体し、外側及び前方方向にfasciculus thalamus視床(核)束を通る。fasciculus thalamicus中の繊維の大部分はVLo, VLmの外側部、VApcに終わる。又CM核にも終わる他にpedunculopontine核(脳幹、上小脳脚繊維に一部埋もれている)。
前障 claustrum
ハッキリした機能は知られていないようだ。全大脳皮質から局在パタンを示して前障に投射があるという点で皮質線条体投射によく似ている。相互結合がある。前障→皮質で運動領へ行くものが両側性(同側優位)なるは興味あり、他の前障遠心系は充分には知られていない。
視床下核と黒質を含んだ大脳基底核の繊維結合は完全にほど遠い。しかい明瞭なるべきことは、これらの脳の部分は、脊髄繊維を与える諸核に対して限られた範囲の影響しか与えないということである。実際、たった一つの重要な候補としては黒質視蓋路のみである。基底核は、何をさておいても第一に、大脳と視床核との間のcollaborationに関与しているように思われる。このことは次の事実から云えよう。即ち、基底核はよく構成された皮質投射をうけており大量の遠心繊維を視床核に送る。他方、最近の研究で基底核の繊維結合は思ったよりももっともっとこみいっていることが判った。臨床との結びつきはgapがある。
最終更新日:2002/09/13

{kind=link}