| 第5章.小脳 |
要約:慶應義塾大学解剖学教室 川村光毅
・比較解剖学的側面
・小脳の縦帯区分
・小脳皮質
・プルキンエ細胞
・顆粒細胞
・小脳皮質への求心性繊維
・小脳皮質の構造の特徴
・小脳核
・小脳の内での体性部位局在
・小脳の繊維結合。一般的ないくつかの点
・前庭小脳路
・前庭神経核からの二次前庭小脳投射
・脊髄小脳路
・三叉神経核小脳投射
・視蓋小脳路
・小脳前核とそれらの結合
・橋核

数多くの解剖・生理学的研究が小脳の分野でなされたにもかかわらず、小脳の機能とその他の脳の部分との協同を正しく理解するまでには至っていない。Sherringtonは小脳に“固有知覚系の頭部神経節”という名前を献上した。が、後年の研究により、小脳は“固有知覚系のみならず他の機能分野の活動にも関連していることが示された。その繊維結合から判断するに、小脳は脳の(他の)殆どどの部位にも影響を及ぼし得ると思われる。従って、一般に小脳は、筋活動を調節regulateするとしられていると同じ様式で神経系が関与するほとんどすべての機能をも調整、制御coordinates and controlsすると推量して良いであろう(surmise)。小脳は幾多の身体機能を完成に行うという欠くべからざるものであろう。しかしながら、小脳は生命にとっては不可欠のものではない。事実、小脳が先天性に欠除した人でもさしたる欠陥なく日常生活を営みえるのであるから。
この10年間の解剖学と生理学上の小脳に関する成果はめざましく。簡潔でしかも意義ある小脳に関する説明(考察)、結合、機能をすることは今日不可能なこととなった。この章は、故に、新しい所見(証明)のやや詳しい提示をもしなくてはならぬ。この新しい情報の多くが、われわれの小脳構成に関する理解をより深めたとしても、これまでのところ臨床神経学に対する影響結果は大きくないものであった。しかしながら、ここでなされる小脳に関する多少とも詳しい記述(考察)は一定の目的に役立つ。即ち、神経系というものの構成の若干の一般的特徴を例示し、その研究に於いて種々の戦略を例示する。
小脳内の機能的区分を反映するが上に、小脳機能を理解する基礎として価値がある。一対の原始小脳(primordia)で2つの部分に分けられる。そのうち一つは前庭核のmatrix(未分化細胞塊)と密接な関係を有し、もう一つはそのすぐ前方に発達するものである。前者は大抵の脊椎動物で大体きまった形をしておる。Larsellに従い。これを片葉小節葉flocculonodular lobeと普通よばれ、片葉と小節からなる(5-1図)。
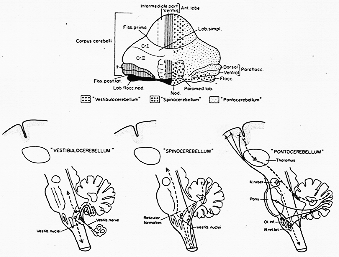
FIG. 5-1 Above, a simplified diagram of the mammalian cerebellum. In the left half the three main subdivisions, which can be recognized on a comparative anatomical basis, are seen. Black; Archicerebellum, flocculonodular lobe. Hatchings: palaeocerebellum, the vermis of the anterior lobe, the pyramis, uvula, and paraflocculus. White: neocerebellum. In the right half the main terminal areas of the vesstibulocerebellar fibers are indicated by heavy dots, the main terminal areas of spinocerebellar pathways by open rings, and the terminal areas of pontine afferents by small dots. The three very simplified diagrams below illustrate that each of the main functional subdivisions, by way of its efferent projections, will influence first and foremost the part of the nervous system from which it receives its main afferent input. See text. From Brodal (1972a).
他の小脳の大部分は片葉小節と後外側裂fissura posterolateralisという一つの裂溝によりへだてられる。この裂溝は系統発生的にも個体発生上も最初に現れるものである。この裂よりも前方に発達する小脳部分は小脳体corpus cerebelliと呼ばれる(図5-1)。片葉小節と対照的に、小脳体は脊椎動物が系統的発生的に高等化するほど大きくなる。しかし、小脳体のすべての部分が等分に大きくなるのではない。最前部は、前葉と呼ばれ、いわゆる第一裂溝fissura primaにより他部と区画されるが、適度の変化(程々の)しか示さない。(第1列は以前にいわれていたように小脳最古の裂溝ではない)。小脳体の最後部は、(虫部)錐体pyramisと(虫部)垂uvulaも又、比較的一定の形を大抵の脊椎動物でしている。大変形が大きくなるのは小脳体の中央部である。その中心部に関してそうであるが、とくにその(小脳体の)外側部が著しい。ここは、哺乳類でのみ明らかに発達しており、サル、類人猿、ヒトではたいへん大きくて小脳の他の部分を完全に被っている。この外側部が大体小脳半球といわれる部分に相当する。前葉の外側部の大きくなる。
小脳の外側部および虫部の中央部は小脳のうちで系統発生上最も新しい部分でしばしばまとめて新小脳neocerebellum(図5-1、左半、白い部分)と呼ばれる。これに対比して、他の部分は時に旧(or古)小脳paleocerebellumとしてまとめれる。しかしながら、Larsell(1934,1937)により提唱された区分によろう。Larsellによれば、片葉小節葉は原始小脳archicerebellum(図5-1、左半、黒)と呼ばれ、旧小脳paleocerebellumは前葉の虫部および錐体、垂、と房片葉(図5-1、左半、斜路)という。
比較解剖学を基礎にした小脳区分は大体、小脳求心繊維結合を基にした区分と相応する(しかしながら、後述をみよ)。
原始小脳-前庭小脳、旧小脳-脊髄小脳、新小脳-橋小脳、の如く言及される。
ヒトの類同は議論のある所であったが、大体明らかとなった。(図5-2)。
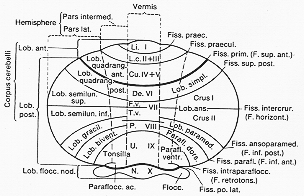
FIG. 5-2 Diagram of the subdivision of the human cerebellum, based on comparative studies of the mammalian cerebellum. In the left half of the diagram the classical names of the various lobules are shown, to be compared with the names used in mammals in general in the right half. On the left is further indicated the principal subdivision of the cerebellum in the flocculondular lobe and corpus cerebelli, the latter again being subdivided into an anterior and posterior lobe. In the vermis the Roman numerals I to X suggested by Larsell (1952, and later) for the transverse foliation are shown. Finally, the longitudinal subdivision into vermis, intermediate zone, and lateral part (cf. text) is indicated. Abbreviations for vermal lobules: Li.: lingula; L.c.: Lobulus centralis; Cu.: culmen; De.: declive; F.c. and T.v.: folium and tuber vermis; P.: pyramis; U.: uvula; N.: nodulus. From Jansen and Brodal (1958).
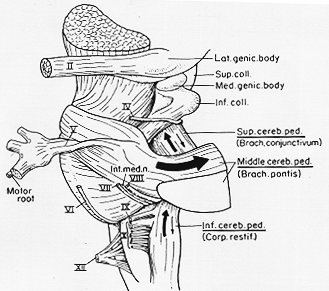
FIG. 5-3 A drawing of the lower part of the human brainstem, seen from the left, to illustrate the gross features of its connections with the cerebellum (removed). Direction and thickness of arrows indicate direction of fibers and (roughly) their relative numbers in the three cerebellar peduncles. See text.
小脳を葉や小葉に古典的に区分するに加えて、最近縦割りパタンの存在が小脳皮質にあることが示された。
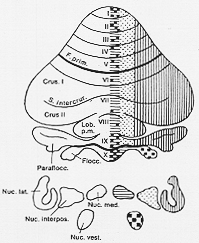
FIG. 5-4 A simplified diagram of the longitudinal zonal subdivision of the mammalian cerebellum based on the patern in the corticonuclear projection. The cortical sites of origin of fibvers to the various in tracerebellar nuclei and the vestibular nuclei are indicated by symbols corresponding to those in the nuclei. From Jansen and Brodal (1958).
Jansen と Brodal(1940, 1942) 3帯. (ネコ、ウサギ、サル)
medial (zone) (vermis)→N. M.
intermediate→N. I.
lateral→N. L. (図5-4)
この見解はChambers と Sprague (1955a,b)の生理、解剖の仕事により支持された。Korneliussen (1967, 1968, 1969)ラット・クジラの小脳皮質と核の個体発生の研究
中間帯を2帯に、内帯を3亜帯に分けた。
中位核が発達しているクジラ類は中間帯が小脳皮質の大部分を占める。
Voogd (1964,1969)詳細にした。細い繊維(raphes, Voogd, 1964)は域帯の境界を作るがどこも等しく明瞭という訳ではない。
皮質と核への求心繊維の終止部位の研究を行い。多くの求心繊維(成分)(contingents)は、皮質とstripを作り分布するのが明らか。例えば脊髄小脳路や楔状束核小脳路cuneocerebellar tract (Voogd, 1969. for review)。
これらの神経路の各々は、一つの帯以上に終止している。このことは、機能的に異なるカテゴリーの繊維を運んでいることを示している。この帯状パタンはとくに下オリーブ核からの終止(域)で詳細にマップされた。(後述、5-18図)。図5-5; A,B (Vermis), C1C2C3(intermediate) D1D2 (lateral).
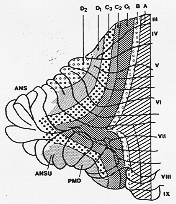
FIG. 5-5 A diagram of the longitudinal zones in the cerebellar cortex of the ferret, according to Voogd. Note that the lateral zones are not arranged parasagitally. See text. From Voogd (1969).
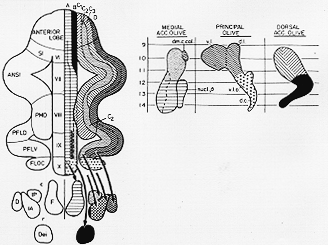
FIG. 5-18 Summarizing diagram of the pattern in the olivocerebellar projection as studied by tracing efferent fibers from the olive. As indicated by different symbols, particular parts of the inferior olivary complex project to different logitudinal zones. These extend more or less continuously through the cerebellum. The diagram further shows the general pattern in the efferent projections (arrows) from the various cerebellar cortical zones to the cerebellar nuclei and the lateral vestibular nucleus (of Deiters). The positions of the olivary subdivisions (right) have been rearranged to facilitate comparisons with the diagrams in Fig. 5-16,17,19, and 20. Some abbreviations: D: dentate nucleus; Dei: lateral vestibular nucleus; F: fastigial nucleus; IA and IP: nucleus interpositus anterior and posterior; d.c.: dorsal cap; dm.c.col.: dorsomedial cell column; d.l. and v.l.: dorsal and ventral lamellae; v.l.o.: ventrolateral outgrowth. From Greoenewegen, Voogd, and Freedman (1979).
生理学的にも(cp. Oscarsson, 1973). 図のようにシンプルでなく。もっと複雑、B帯、C2帯の如く全域に互らぬもあり、又、或る系は上肢、下肢のinformationの如くzoneの特定の部分に終わるものあり、全帯域に分布するものではない。
たくさんの深い溝のためヒト小脳の最前から最後までの距離は1メートルを超える(BraitenbergとAtwood, 1958)多少の領域差があるが、小脳はどこをとっても基本的に同一構造を呈する。分子層、Purkinje細胞層、顆粒層、プルキンエ細胞、フラスコ型、整然と配列。たくさんの樹状突起。(一つの面にひろがる)-小葉の長(タテ)軸に垂直な面。自己の領域territoryを有するが如し、棘がありシナプス結合1つのプルキンエ細胞の棘の数のネコで8万個、ラットで1万8000個、平行繊維と結合。棘は樹状突起の近位部にも起こる。登上繊維は棘とのみ結合する如し。
核と前庭神経核へ軸索、それは反回副側枝を出し、一面上でその細胞の樹状突起につく。一部は細胞体や近位樹状突起やGolgi細胞につく。Golgi細胞は顆粒層内にある。
細胞質乏しいscanty、4-5本の短い樹状突起、色々の方向に放射する(鳥獣の)つめ状のひろがりclaw-like expansion苔状繊維終末結合。軸索は特徴的で分子層まで上行しT型に2分し平行繊維parallel fibersと呼ばれ、常にプルキンエ細胞の樹状突起dendritic treesを貫く形で小葉をタテ方向に走っている。平行繊維の長さは2分れらた部分を合わせて1.5-3mm (Fox と Barnard, 1957)(ネコ)でヒトではやや長い程度。平行繊維はPurkinje細胞の棘と結合(他に星状細胞、籠細胞、Golgi細胞とも)。1個のプルキンエ細胞のdendritic treeを貫く平行繊維の数は、20-40万本(ネコ)。これからして恐らく1本の平行繊維が約450個のプルキンエ細胞と結合するので、各々のプルキンエ細胞は恐ろしい数の顆粒細胞の影響下に入る。
顆粒層内のもう一つの要素は、ゴルジ細胞である。ある面でプルキンエ細胞に似ている。細胞は大型で分岐した樹状突起treeをもち、分子層にまでも広く外に広がっている。しかし異なる所は、樹状突起がすべての方向にひろがる点である(5-6図)。樹状突起は平行繊維とも他の求心繊維(若干の樹状突起は顆粒層に残り、且苔状繊維と結合している)とも結合する。ゴルジ細胞の軸索は豊富に分岐しているが小脳皮質を去ることはない。
分子層は繊維が多く比較的に少数の神経細胞を含んでいる。それらのあるものは星状細胞stellate cellsで数型ある。特殊型としていわゆる籠細胞basket cellがある。これはプルキンエ細胞のすぐ上に位置する。その樹状突起はプルキンエ細胞のそれと同じく小葉の横断面にのびており、登上繊維の側枝を受けている(後述)。籠細胞の特色はその軸索の配列である。この軸索は可成り長い距離を小葉を超えてPurkinje細胞のすぐ上を走直角に下行する側枝を出す。これらの側枝はプルキンエの細胞体をとりまいて、それとシナプス結合をもつ。これらのゴルジ細胞の配列があることにより、小葉間-を超えて配列してる一連のプルキンエ細胞に働きかけることができ、これは、小葉(の縦軸)に沿った一連のプルキンエ細胞を活動する平行繊維と対比している。これらのそして他の特殊な幾何学的配列は小脳皮質の機能を解析する上で興味深いものである。
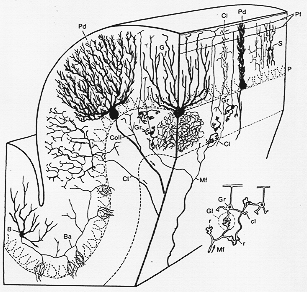
FIG. 5-6 Semidiagrammatic representation of part of a cerebellar folium to show the main elements of the cerebellar cortexx and their topographical relationships and orientation. Note especially th earrangement of the Purkinje cell dendrites (Pd) and basket cell axons (Ba) in the transverse plane of the folium and the longitudinal arrangement of the parallel fibers (Pf). Other abbreviations: B.: basket cell; Cl: climbing fiber; Coll: recurrent collateral pf Purkinje cell; G: Golgi cell; Gr: granule cell; Mf: mossy fiber; P: Purkinje cell; Pd: Purkinje-cell dendrites; Pf: parallel fibers; S: stellarte cell. Slightly altered from Jansen and Brodal (1958). The figure to the lower right illustrates diagrammatically the relation between the rrosettes (r) of the mossy fibers (Mf) with the dendritic "claws" (cl) of the granule cells (Gr) in a glomerulus (Gl). Other elements of the glomerulus are not shown. Adapted from Hamori and Szentagothai (1966a).
登上繊維 細いgranular layer →Purkinje cells dendrite branch(←spineにシナプス結合)に沿ってfollow and wind (along)巻きつく。主たる標的Purkinje細胞に行く他、側枝がとなりにあるプルキンエ細胞、星状細胞、籠細胞、ゴルジ細胞終わる。(Scheibel と Scheibel, 1957; Hamori と Szentagothai,1966a)プルキンエ細胞に強力なシナプス作動をする。登上繊維の大多数は下オリーブ核、他の脳幹の核からもあるかもしれない。1本の登上繊維が1ヶのプルキンエ細胞へという特別な関連があると云われていたが、実際は皮質の直下又は内部で分岐し、2-3-4のPurkinje細胞を支配する(互いにそう離れてない所)。生理上は1本の繊維が分岐し、かなり離れた小葉を支配する(Faber と Murphy, 1969; Armstrong et al. 1971, 1973a; Cooke et al., 1972)。同一帯状内で分岐する。
両側オリーブ核細胞の数は約100万個(ヒト)。プルキンエ細胞の1/15にあたる。ネコでは両側オリーブ数12万~14.5万個でプルキンエは1.2-1.3-1.5 million故これも1/10にあたる。(研究者名と年号略)
苔状繊維はあらゆる点で登上繊維と異なる。比較的太く有髄。皮質内で何回も分岐を繰り返す。1つの繊維が2つ又はそれ以上の小葉を支配する。経路中たくさんの側枝を出しそれらは、終末枝さながら、房状に小さい終末をロゼットとしばしばよばれるものを作って顆粒層内に終わる。これらの終末は顆粒細胞のつめ状の樹状突起といりくみ合ってシナプス結合をする(5-6図)。この部分は小脳の糸球体cerebellar glomerulusと通常呼ばれるものに属している。
(接触要素)
細胞染色標本では神経終末は染まらないのでglomeruliは顆粒細胞の内に抜けた空隙としてみられる(“cerebellar islands”)
EM:1本の苔状繊維がたくさんの異なる顆粒細胞からの樹状突起とシナプス結合を糸球体内にみることがある。前述したように、ゴルジ細胞の軸索も糸球体内に終わっている。このGolgi axonsは顆粒細胞のdendritesとシナプス結合をしている(又、逆にGolgi cell dendritesに苔状繊維の終末がついている)
苔状繊維の顆粒細胞に対する影響は興奮性であるが、Golgi細胞の活動は抑制性である。
オリーブ以外、ほとんどmossy,系により分岐の度合い(程度)が異なる。
regularity規則性と各要素間にみられる幾何学的パタンgeometrical patterns→機能に反映(仮定)。
プルキンエ細胞への抑制経路は興奮経路よりも1シナプス多いので、平行繊維を刺激するとEPSP後1-2 msecしてから1 PSPが現れる。
Purkinje細胞は抑制性(小脳核、前庭核)(Ito と Yoshida, 1966)。しかしすべてのプルキンエ細胞が然りとはすべての研究者が確信している訳ではない。
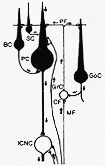
FIG. 5-7 Diagram of the most significant neuronal connections in the cerebellar cortex, according to physiological studies of Eccles and his collaborators. Cells and terminals shown in black are inhibitory. Abbreviations: BC: basket cell; CF: climbing fiber, GoC: Golgi cell; GrC: granule cell; ICNC: intracerebellar nuclei; MF: mossy fiber; PC: Purkinje cell; SC: stellate cell. See text. From Eccles (1966a).
図5-7は、登上繊維と苔状繊維を刺激(activate)して小脳皮質で見られるインパルスによっておこるであろう多くの可能な回路のうちの若干のものをとりあげて示したものにすぎない。明らかに求心インパルスの到達時点が、その結果おきる活動にとって重要である。更に、一定の領域に終わる苔状繊維は多くの源から起こってる来る。異なる種類の生理学的意味をもった情報を選んで来るものであることを思い起こすべきでしょう。条件は確かに非常に複雑であり、しかも1つ又は2~3の要素が別々に研究される実験動物におけるよりも、成体で条件はより複雑である。
しかしながら、これらの新しい知見は小脳皮質の働く機械として興味ある(討)論議を生んだし、機能の主たる様式の模型を作り上げることを促進した。ここでは、種々の要素とこれらの幾何学的配列の性質の観察に基づいて(た)推論(演繹deductions, 普遍命題→特殊命題)が結びつけられた。この主題の説明に関しては読者はEccles (1966a) と Eccles, Ito と Szentagothai (1967)を参照されたい。
最近、Falck と Hillarpの組織蛍光法を用いて製版核からのアドレナリン性繊維縫線核からのセロトニン性繊維が小脳皮質に至ることが示された。これらの繊維と古典的な苔状繊維と登上繊維との関係はなお明らかにされていない。小脳の構造と機能は充分には明らかにされたとは云えない。総説と多くの詳細についてはChan-Palay (1977)のほんの中に見い出される。
アドレナリン性繊維 少、散在性のものプルキンエの樹状突起と棘にシナプス結合(Bloomら、1971)。EM歯状核(外側核)にラット、蛍光繊維2型(CAT1,CAT2)アドレナリン性とセロトニン性に相応と推量する。
室頂核
球状核
栓状核
歯状核
核内に種々の大きさと型あり、細胞構築物にminor regionあり、一様な結合と機能を1つの亜核がものではないことを示す。未解決のFlood と Jansen (1961). Courville と Brodal (1966) Brodal と Courville (1973) NLとNIAの境がF.J.より少し外側より。
小細胞群(SMP) 1. NMの腹側部 subnucleus pervicellularis medialis
(SLP) 2. NLの腹側部 subnucleus pervicellularis lateralis 特別の結合(後述)
アカゲザルの小脳核Courville と Cooper (1970)
小脳核への主たる求心繊維はプルキンエ細胞の軸索
他は、脊髄、下オリーブ核、橋核、小脳前網様核、赤核、他、多くは側枝らしい。
小脳からのほとんどすべての遠心繊維は小脳核の細胞の軸索である。
行先は:前庭神経核、赤核、視床、網様体、下オリーブ核、他
核内の構成(造)とその機能 cp. Vicky (1977)
どの亜核でも基本は似ている。(松下と岩堀、1971a,b; Angaut と Sotelo, 1973; Sotelo と Angaut, 1973)。プルキンエ軸索は核に入ると豊富に分岐するが、各々の繊維は大体1つの円錐域を支配する(Cajal, 109-111)。プルキンエ数と核ニューロン数=約26:1で一本のプルキンエ細胞軸索は35個の(平均)核ニューロンとシナプス(主にaxodendritic)結合をする。(Palkovits, Mezey, Hamori, と Szentagothai 1977).
小脳核の細胞からのaxonsは側枝を小脳核に出す。小細胞に結合(松下と岩堀1971c)。介在ニューロン?
最初にBolk(1906,オランダの解剖学者)が説えた。哺乳類の比較解剖学を基礎にして、身体の筋肉の量(mass)と小脳の一定部位との間に関連(並行)ありとした。(仮説)
eg. 前肢→Bolkのcrus I
後肢→Bolkのcrus II
熱烈に迎えられたが、必ずしも一致せず決定的な結論とはならなかった。
最初の確信持った証明は(自然又は電気)刺激→活動電位。
Adrian (1943), Snider と Stowell (1942, 1944). Cp. 5-9図。
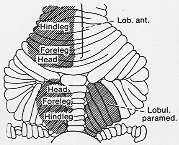
FIG. 5-9 Diagram of the surface of the cerebellar cortex of the monkety, showing (shaded) the areas in which discrete movements of hairs on the left side of the body give rise to action potentials. The somatotopic "representation" is indicated. In the middle part of the vermis and overlapping with the Vermis and overlapping with the "head areas" is the audiovisual area. See text. From Snider (1950).
同側前葉と両側Paramedian lobule. 皮膚受容器、又は、皮膚神経からよく得られる。(体性)虫部中央部(顔面域と一部重複)から聴・視刺激(+) 活動電位
対側の大脳皮質を刺激した後も、同じ領野から(+)。
Adrian (1943; Snider と Eldred, 1948, 1951,1952; Hampson 1949)後に内側・外側の局在も見い出させる。
B帯の外側→後肢(B1)
B帯の内側→前肢(B2)
求心系はすべての受容器から(固有知覚、(一般)体性知覚、前庭、聴覚、視覚など)すべてから来、すべて(殆ど)に行く。直接・間接に影響 結合は広く相互的、複雑、然るにいくつかの一般的な特徴があることに注目すべき也。
第一に、小脳からの遠心繊維数/求心繊維数は著しく劣り1/40である(Heidary と Tomasch, 1969)。つまり入出力関係でみると入力がはるかに優る。繊維の情報伝達力が完全に発揮されていると合理的に仮定してみたとき、小脳の作業効率をみてみる(計算して)、入力シグナルの5%以下しか、作業結果を伝えるのに要していないことになる。
遠心性結合に関して云えば、皮質からのものすなわちプルキンエのaxonsの大多数は小脳核より先に行かぬ(少しは前庭核へ行く)。核からは、しばしば中継部位を介して、小脳メッセージを他の領域に伝える(脊髄、大脳皮質、他)。
求心性繊維については、直接(前庭装置)もの又脊髄から(dorsal と ventral spinal cerebellar tract)又、中継所、relay stations in脳幹(下オリーブetc.)数種のインパルスの集中convergenceの座、多く相互結合 直接
重要なものは:橋核、下オリーブ核、小脳前網様核。他に弱い小脳投射のあるものとして、perihypoglossal nuclei、縫線核、青斑核、赤核など
前庭装置から小脳へインパルスを伝える経路。前庭神経節の細胞の軸索が直接行くものと中継される間接的なものあり。
Dow (1936) Marchi法 一次前庭神経繊維→片葉小節、虫部垂の腹部、および室頂核(NM)。
Dow (1939) ネコ・ラットVIII神経の刺激(電気的)で小脳の同領域に活動電位。
Brodal と Hoivik (1964) 鍍銀変性法で以上の他に腹側旁片葉と歯状核の小細胞部分(SLP)に終止を見たが、NMへの終止は確かめれなかった。
最近、一次前庭神経の小部分(minor portion)が全虫部域に終わることが示唆(HRPでKotchabhakdi and Walberg, 1978a)。
電気的又は自然刺激を前庭装置に与えたのち、全虫部域と中間帯部の領野にpotentialsが記録されている(Prechtら、1977, Ferin, Gregorian と Strata, 1971)。しかしながら、これらの反応のうちには中継核を介するものも含まれるかもしれぬ。
Dow(1936)二次繊維の分布は一次繊維の小脳分布と相応する。苔状繊維と思う(Carreaら、1947)。グッテン改良法で起始部は前庭神経核の内側核と下核の一定部位及びGroup x of (Brodal と ポアンペア1957)に限られている(Brodal と Torvik, 1957)HRP法でも以上の領域および前庭核群の他の部分からも少し投射があることが確かめられた(Kotchabhakdi と Walberg, 1978b)。彼らは、片葉小節葉以外、たとえば前葉及び後葉の虫部にHRPを注入した例でも前庭核にいくつかの陽性細胞を見い出した(Precht, Volkind とBlanks, 1977もみよ)。 一次と二次の前庭小脳投射の他にも、多少の関節ルート(脳幹経由e.g.二次繊維をうけて小脳に投射する外側網様核)があるようだ(Prechtら、1977が示唆)。
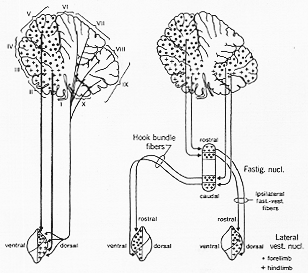
FIG. 5-30 Diagram illustrating major features in the projection from the cerebellar cortex onto the nucleus of Deiters (to the left) and (to the right) in the projections from the cerebellar cortex onto the fastigial nucleus and from this to the lateral vestibular nuclei. Note that the direct cerebellovestibular fibers and the projection from the rostral part of the fastigial nucleus end in the dorsal half of the ipsilateral lateral vestibular nucleus, while th efibers from the caudal part of the fastigial nucleus via the hook bundle supply the ventral half of the contralateral lateral vestibular nucleus. Within each of these projections there is a somatotopic localization. (See text). Slightly altered from Brodal, Pompeiano, and Walberg (1962).
古典的な経路のDSCTとVSCTの他にRSCTと(external)外側(後柱を上行する一次知覚繊維を受ける)楔状束核を経由するもの(E)CCT(一般にcuneocerebellar tractと呼ばれている)がある。→前肢から
又最近中心頸核central cervical nucleusからの経路も発見された(後述)。
背側脊髄小脳路DSCTは、長いことクラーク柱(背核)から起こるとされていた。Th1-L2(ヒト)[ネコでは~L3-L4まで拡がる]RexedのVII層(後角の基部)やや太い有髄繊維同側性、(外索)外側皮脊髄路の背外域を上行する(Fig.2-3B. cf. p.63)下小脳脚に入り、小脳の”spinal region”に終わる(後述)。
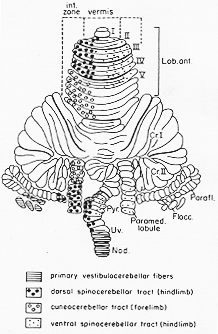
FIG. 5-10 Diagram of the rcerebellar surface of the cat, showing the distribution of the fibers of the dorsal and ventral spinocerebellar tracty and of the fibers from the external cuneate nucleus as determined by Grant (1962a, 1962b). Note somatotopic pattern. The terminal areas of primary vestibular fibers as determined by Brodal and Hoivik (1964) are aoso shown.
Marchi法で、DSCTの終止領は主に、同側性に前葉(虫部と中間部)と後部虫部主に錐体pyramis paramedian lobule (正中旁小葉)にも終わるとした鍍銀法でGrant (1962a) がDSCTは前葉と正中旁小葉の後肢域にのみ終わることを示した。(5-10図)。錐体(VIII小葉)の尾部にもいくらか終わる。小脳に到達する前に内・外に配列した一連の束に分離し、縦帯に終わる。Voogd (1969)は諸動物で6つの束を前葉に区別したが、大多数の繊維はC帯を給する。生理学的にも前葉に終わるDSCTの繊維は主に中間帯にintermediate partに終わることが確かめられた苔状繊維(解剖学的にも又responseタイプも然り)。(Oscarsson, 1973)、DSCTは当初の間、固有受容器からのインパルスを伝えるとされていた(Grudfest と Campbell, 1942; 他)が、単一繊維からの記録からLundbergと共同研究者は(Lundberg と Oscarsson, 1960をみよ)。いくつかの(数種の)機能的な成分を区分し得た。
筋紡錘、腱器官、無毛の掌手からの圧受容器、有毛皮膚部の触・圧受容器からの情報を伝える。関節受容器から単シナプス的に興奮するニューロンがクラーク柱にも発見(Lindstrom と Takata, 1972)。多くは小さい受容野をもつ。1cm2のみ、又、単一筋からactivateされる。DSCTは明らかに下部体幹と下肢からの様式及び室間-特異的な情報を運ぶ。modality-and space-specific external, lateral, or accessory頸髄でのクラーク柱に相当するものは外側(副)楔状核である。(モナコフ核ともよばれる)CCTこの核は薄束核や主楔状束核と対比して、相当数の大型細胞を有する。同側性、下小脳脚、ほとんどのcellsが遠心ニューロンらしい。C1~Th4-5の後根繊維が求心繊維。cuneocerebellar tract (副楔状束核小脳路) 頸部、前肢、上部体幹。終止域は、前葉中間部の後方域(主にV小葉)と正中旁小葉の前方部の他に少し後部虫部に。Cooke, Larson, Oscarsson と Sjolund (1971a)によれば、ECNの細胞は殆ど2分し小脳のこの二つの領域に体尾在投射する。←ECNの一部は視床核に投射する(2章)。←頸髄と上部胸髄の後根繊維。(空間特異性が高い)
CCT1つ以上の成分より成る(Oscarsson, 1973)副楔状核のみでなく楔状核からも恐らく薄束核からも小脳へVSCT,RSCTは脊髄から小脳へ直接投射であるがDSCTやCCTと多くの点で異なる。VSCT(昔からGower路)主として交叉性に側索を上行、DSCTの腹側を上行。下小脳脚に入らず延髄橋を上行し、三叉神経根繊維の上を曲がって背外側にまがり上小脳脚から小脳に入る。Marion Smith (1957)によれば、ヒトでは多くの繊維がDSCTにjoinする。cp. 5-10図。一部が小脳内で交叉する(正中部で)。後肢、下部体幹からのみのインパルスを伝える(Oscarsson, 1973をみよ)。Cooper と Sherringtonはサルで”spinal border cells”から起こると示唆した。(1940)前角の背外側に主に表層性に位置する幾分大型の細胞。後にネコでも、Sprague(1953), Matsushita と Hosoya (1979) Hubbard と Oscarsson (1962) V-VII(主にVII)L3-6(ネコ)
VSCTに相当する前肢よりのインパレスを伝えるものにRSCT(OscarssonとUddenberg, 1964)機能的に多くの点でVSCTに相当する起始細胞の受容野は同側性で前肢に関連している。同側性(非交叉性)に上行し、一部上小脳脚に、一部下小脳脚に入るようだ。松下と細谷、VII(C4-8)。
VSCT, RSCTは共に屈曲反射求心繊維により多シナプス性に強く活動化activateされ、せれ、脊髄レベルで高度に統合された情報を運ぶ、屈曲反射求心繊維のRSCT細胞へのactionは主として興奮性であり、一方VSCT細胞へは抑制が優位である。この2つの神経路は多くの点でDSCT, CCTを明らかに異なる。これら4つの神経路は苔状繊維として終わるようだが(文献略)。DSCTとCCTの単一繊維により介されるインパルスはVSCT, RSCTからのものよりもはるかに狭く小脳領域に分布しており、このことはこのことは後者の苔状繊維はより豊富に分岐していることを示すものだという生理学的に証拠がある。
一次性V-fiberが小脳へ行くことは疑わしいが二次繊維の小脳投射は確率された。Carpenter と Hanna (1961) interpolarisとoralis(三叉神経脊髄路核の)から、同側性(主に)V小葉後部とVI小葉中脳路核顔面からの固有感覚インパルス、有るか?
昔の仕事、(OgawaとMitomo, 1938)正常標本、あったとしても量は少ない。
小脳前核は多くの源から求心繊維を受ける。一部小脳からも
大脳皮質から小脳への最も重要な中継核、Ndl.大型細胞、Nv.小細胞系統発生的に大きくなりヒトでは片側に2000万個のニューロンあり(Tomasch, 1969)。背側にnucleus reticularis tegmenti pontis of Bechterewあり。構造は多くの点でreticular formationに似る。すべての細胞が小脳へ軸索を送るようだ。多くの繊維は比較的細い、苔状繊維中小脳脚、主として交叉性、昔の解剖学研究(?)(Jansen と Brodal, 1958; Larsell と Jansen, 1972をみよ)小脳半球や旁片葉に豊富に繊維を送るが虫部へは比較的少ない。
半球へは主に対側性だが虫部には両側性。片葉投射も少しある。小節(nodulus)は橋核投射をうける唯一の小脳部分と思われる。小脳核もうける。いくつかの橋核部分から一定の小脳皮質域へ、柱、縦状、ラット(Burne, Eriksson, Saint-Cyr, と Woodward 1978)の橋核小脳投射も似たパタン。
サルの構成も同じパタン(P. Brodal, 1979)。起始ニューロンの柱状パタンはネコに比してハッキリしない。一部fuse。ネコと少しの異の小脳終止領の重複はネコに比して少ない。
橋核小脳路は非常に正確に構成されている。converge - divergence特定小脳小葉へ、小橋核域から。
皮質橋核投射にも
同様の原理原則principleある如し。橋核へは上丘、下丘、小脳核、他から来るが、求心繊維の最大はcortexから、体性局在パタン、end within circumscribed, approximately longitudinally oriented columnar or more lamellar-shaped pontine areas.皮質の“運動性”、“知覚性”橋核終止域はネコよりもサルで分離している。よりハッキリした機能分化を示している(反映している)。サル(P. Brodal.)皮質-橋核路(1978b)
強い投射。area4, 3,1,2,5,視覚野(peripheral visual field)
弱い投射。central regionの前方と後方
皮質脊髄路の側枝によって橋ニューロンがactivateされると生理学者達に考えられていたが、之は程度が限られている。ヒトで橋縦束(peduncle): 錐体=20:1である。側枝支配の多い橋部とそうでない所があるのであろう。cp. Allenkom, Oshiwa と Toyama (1975)の成績は代表例といえぬ。Jones と Wise (1977)
一般に1つの部位に繊維を送る細胞は他の部位に側枝を送るとは思われぬ。皮質橋繊維は橋ニューロンの樹状突起のみにシナプス結合(Hollander, Brodal と Walberg, 1969)介在ニューロンの存在も? Golgi (Mihailoff と King 1975; Cooper と Fox, 1976)とEM(Cooper と Beal, 1978)研究で主張。corticopontine fiber単シナプス、fast fiberとslow fiberあり(Allen, Korn と Ooshima, 1975)。
生理でシナプス関係、集合(求心インプット)を云々するとき部位を明示する要あり、橋核は広大なるニューロンの集合体であり、構成が複雑で分化している故に、
Allen と Tsukahara (1974) review.
小脳半球とNL. 運動のプレプログラミング
中間帯とNI updating of ongoing movements進行中の運動を更新する。
運動の発動と遂行における運動領と頭頂連合による相異なる役割を考えてみるとき、(4章をみよ)これら2つの皮質から橋核への投射とそこから更に小脳への投射は興味がある。
橋核ニューロン:皮質橋核繊維=1:1.6 (Tomasch 1969) cortex以外にも橋核求心繊維あり、最も重要な可能性は小脳核から、上丘から、下丘からVGL. →paramedian nucleus (Graybiel, 1974). Pretectumから(Itoh, 1977). 小脳核から上行性上小脳脚からNLとNIAから起こる。交叉性の下行枝により。橋核内に3つの縦柱に、NIPないようだ(Angautをみよ、1970)。NM(hook束を離れ)から若干がオホムス、とサルでみられている。脊髄から少し(Walberg と Brodal 1953b; Kerr, 1966; Ruegg, Eldred と Wiesendanger, 1978).弓状核arcuate nucleus.(ヒトでのみよく発達)とpontobullar bodyもcortexからafferent, cerebellumへ送る。
最終更新日:2002/09/13
