| ��U�́D�]���ԗl��(Reticular formation) |
�v��F�c��`�m��w��U�w�����@�쑺���B
�E�ԗl�̂Ə�s�������n
�E�ԗl�̂̉�U
�E�ԗl�̂̑@�ی���
�E��s������
�E���S���@��(reticular formation��)
�E�ԗl�̂̍\��
�E�D���j
�E���s���̉��S���@��(�D���j�����)
�E���j
�E���S������
�E���S�D����
�E�ԗl�̂̋@�\�I����

�@�]���ԗl�̂͌ÓT�I�_�o��U�w�҂ɂ��CNS�̒��̂P�̕������ꂽ�����Ƃ��ĔF�߂�ꂽ�B���I�̕ς��ڂ�reticular formation�̌�������Ƃ��Ēm���A���̋@�\�Ɋւ��鉼�����ł��o���ꂽ(�E���ɂ�)�B�������Ȃ���A�`�Ԋw�҂��A�����w�҂��A�Տ��Ƃ������I�O���̊Ԃ�������ڂ��Ȃ������B����ÓT�ƂȂ���Moruzzi��Magoun�ɂ��g�]���ԗl�̂Ɣ]�g�̊����h��1949�N�ɏo�ł���ĈȌ�A���̏���͑S����ς����B���̎傽�闝�R�́A�ԗl�̂��A����ꂪ�ӎ��ƌĂԂȂǂ��߂��@�\�̈ێ��Ɋ֗^���Ă���@���v��������ł���B����ɂÂ��āA�����w�I������Moruzzi��Magoun�̂��̍ŏ��̊ώ@���m�F���X�ɂ��������߂�(Rossi �� Zanchetti, 1957�̑������݂�)
�@Chloralosane�Ŗ������ꂽ�l�R�Ŕ]����d�C�I�ɍ��p�x�̎h����^����ƁA�]�g��̍��k�����g(���F�����p�^��)�̓��������˂���U�����g����(���F�o���p�^��)�ɒu���ւ�����B����̓q�g�]�g�ł݂��郊���b�N�X�����ʂނ�����(drowsy)��Ԃ��璆�ʂ��ނ����o���������(alert)�ւ̈ڍs�Ɍ����鑊������ω��ɑ�ς悭���Ă���B�����̕ω��͈�ʂɒE������(desynchronization)����(activation)���͊o������(arousal reaction)�ƌĂ��B�����͑O�玿�ɍL�͂ɋN����B�g�L��������(����)�n�h(diffuse thalamic system, H.H. Jasper)�����ƍl�����Ă���(��Q�͂Ō��y���������̏͂ł���ōl�@����)�B�����̕ω��́A�����̉���reticular formation,������ђ��]�̔�W����ɔw�����������A����ю������j(subthalamus)�̎h���ŋN����B���l�̕ω����Ґ��_�o�A�O���_�o�A�����_�o(?, splanchnic), �V���_�o�A���_�o�A���̔]�_�o�̎��R���͓d�C�h���ɂ���Ă���N�����B(�����ɂ��Ă�Rossi �� Zanchetti, 1957)�B�����̌o�H�ɂ��reticular formation�ɂ͂��Ă���h���́Apotentials���L�^�����B��]��d�C�I�ɖ��̓X�g���q�j���ɂ��h������ƁA���l�ɁAreticular formation��potentials��^����B���A���A�����̒��́g���ʗ��f�]encephale isole�h�l�R(�Ґ������̔]�������番�����ꂽ�l�R�F�����E�Ґ��Ԑؒf�l�R�|�g���hBremer, 1937)�ɂ�����EEG arousal�]�g��̊o���p�^���������N�������Ƃ�����B�����j�h����reticular formation����Ĕ玿�d�ʂɉe�����y�ڂ��B�]���Łg���ꊴ�o�H�h��ؒf������(�����ётƐҐ������H��)�A�_�o��reticular formation�̎h���Ŕ]�g�̕������Ȃ���������B�������Ȃ���A�]���̒��S����j�Ă��܂��ƁA�s����Ö���ԂƂȂ�A�玿�d�ʂ͓�����synchrony�ƂȂ�B�����āA���S��������̎��͗L���ł������h���łɂ���Ă��������͓����Ȃ��Ȃ�B���l�̌��ʂŃT���ł�(French, von Amerongen �� Magoun, 1952)�B�����y�ё��̊ώ@����A�g��s���ԗl�n�h(ascending reticular system�h���]���ɂ���ƌ��_���ꂽ�B���̊T�O�͍X�Ɍ��������elaborate���ꂽ(���߂��܂������ꂽ)�B���̌n��tonically active(������or�ْ����Ɋ���������)�������x���͋��S�C���p���X�̗ʁAhumoral�̕����A�A�h���i�����ACO2�Ɉˑ����A�n�̊����͔]�g�ɔ��f���A���S�Ȋo���ƏW�����炤�Ƃ��Ɓ`�����Ɏ��邳�܂��Ȏ�̈ӎ����x�������肷��ƍl����ꂽ�B���̌n�́g����h�g���o�n�h�ƑΏƓI�ɁA�S���L���ɍ\������Ă���ƍl���ꂽ�B��s�������n�̍\����̊�́A�]����reticular formation�g����ꐫ�h�����j(�O�҂����҂ւ̏�s���������܂�)���X�Ɏ�������玿�ւ̓��˂ł���B���̏�s���`�B�̐����w�I�ώ@�ɂ���A�̒Z���������������זEshort-axoned cells���琬��Ƃ��ꂽ�B�X�ɁA���̌n�̕����͑��`�I�ɓ��ꊴ�o�H����́g���}�ɂ��Ђ낪��h(collateral spread)�ɂ��N����ƌ��_���ꂽ�B
�@�g�����n�Aactivating system�h���]���ɑ��݂��邱�Ƃ͊m������A���̔������]�̏d�v�ȋ@�\��啝�ɗ��������߂����A�Ȍ�̂��̌n�̉�U�w�I��ՂɊւ���C�����N�����ė����BMoruzzi��Magoun(1942)�͂��̓_�A�ނ�̌��_�́A��U�w�m�������Ȃ����̂ɔނ�̌��_�͈ꎞ�Itentative��Ɩ����ɏq�ׂĂ���B��N���̌������̒f��(���Ƃ��)�|�ۗ��|���������ꂽ�B���҂炪�hreticular�ԗl�́h�Ƃ������Ƃ𖽖�(ascending reticular system��)�Ɋ܂߂����Ƃ�(�s�K)�܂��������B����͌`�Ԋw��̌��t�ł��萶���w�I�m�����A�`�Ԋw�ɒ�߂�ꂽ�ԗl�̂��������ɍL���Ă��܂������Ƃ͖����ł���B�����̈ꕔ�͕����̃��J�j�Y���Ɋ֗^���A�̂�anatomist�ɂ��ԗl�j�Ɩ��Â���ꂽ�Ƃ��Ă��A�V��reticular formation�Ƃ͐����ɂ͊܂߂邱�Ƃ͏o���ʁB��������ɂ������Ė������������Ă����ׂɑ傫�ȍ������N�������B�����n�̓��`��Ƃ��Ėԗl�n�Ƃ������Ƃ������Ηp�����Ă��邪�A����͑����Ԃ������ɂ��Ƃ�����邾���ł���B�hactivating system�h�͋@�\�I�T�O�ł���A�hreticular formation�h�͌`�Ԋw�I�T�O�ł���B���Ƃ͖����ɂ��ׂ��ł���B���̗��R�ŁA���̋��ȏ���Moruzzi��Magoun�̊T�O���]�X����ۂɂ́hreticular�h�Ƃ������t�͗p���Ȃ��B�g��s��ascending�h�Ƃ������t���c���Ă������Ƃ��܂���(���������)�Ȃ��Ȃ�A��s�������Ɋ֘A����̈�͑������銈����Ґ��ɂ��ׂ�����ł���B�g�����n�h�ɂ��Č��Ƃ��g�]���́h�Ƃ������t���Ȃ��������悢�B�]���Ƃ����p��͈قȂ��Ďg�p����邩��ł���B��ʂ̎g�p�Ɏ^�����Ă����ł́A����I�Ӗ��Ɏ��A�����A���A���]���܂�ł����ł͎g�p���悤�B�ԗl�̂̍\���������ɂ��Đ���������ɁA�����́A���̋@�\�ƕ����n�̎��ɗ����߂낤�B
�@�]�̂��镔�ʂ�������ԗl�\����悵���̂��߂Ɂg�ԗl�́h�Ƃ����邱�Ƃ��������Ƃ��Ă��A�Ⴆ�ΐҐ��̔@���A�����ł͔]���̖ԗl�̂���舵���B��ɏq�ׂ����E����(��`)�ɏ]���āAMedulla, pons��mesencephalon���������ƂɂȂ�B�n�������I�ɔ]�̌Â������A�̂̉�U�w�҂ɂ�薼�t����ꂽ�B���̓����́A�\����A���ׂĂ̕����ɑ��邽������̑@�یQ�ɂ�蕪������Ă����X�̌^�Ƒ傫���̍זE�̎U�ݐ��̏W�c(diffuse aggregations of cells)����ł��Ă���B���E�̂͂����肵���זE�Q�A�Ԋj�A��I���[�u�j�A�]�_�o�j�́A�܂܂�Ȃ��B���Ȃ�֊s�̂Ƃꂽ�jNrl(nucleus of the lateral funiculus�Ƃ�����)�ANrt(Bechterew)�͍\����reticular formation�ɑ����邪�A��ʂ�reticular formation�Ƃ������Ƃ��ɂ͏������B�����̊j��(paramedian reticular nucleus�Ƌ���)reticular formation�̎c��ƈقȂ�A���]�ɓ��˂���̂Ő����ȗ��R�ƂȂ낤�B����͂T�͂ŏq�ׂ����A�������đ��̋@�\�I�ɂ����ƂȂ�B
�@�D���jraphe�A�ԗl�\���ɂ��ւ�炸���ʖ��́A�̂ɊܗL�����B
�@�ȉ�reticular formation��Nrt, Nrl, Nrm���������]�|���|�����̖ԗl�\����悷���Ƃ����G�c�ɔ]���̒��S���A�����ɒ��_�o��(medial longitudinal fasciculus, medial lemniscus, spinothalamic tract)�Ɛڂ��B
�@Meessen �� Olszewski (1949) �E�T�M (�זE�Q�ɕ�����)
�@Olszewski �� Baxter (1954) �q�g
�@Brodal (1959) �l�R
�@Valverde (1962) ���b�g
�@Petrovicky (1966) �������b�g
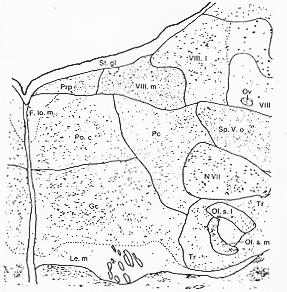
FIG. 6-1 Semischematic representation of part of a Nissl-stained section through the human brainstem at a low pontine level. The facial nucleus (N. VII), the rostral part of the spinal trigeminal nucleus nucleus (Sp. V.o), parts of the vestibular nuclei (VIII.l and VIII.m), the nucleus praepositus (Prp), the medial longitudinal fasciculus (F.lo.m), and the medial lemniscus (Le.m) surround the reticular formation. Within this three subdivisions can be separated at this level, the nuclei gigantocellularis (Gc), pontis caudalis (Po.c), and parvicellularis (Pc). From Olszewski and Baxter (1954).
�}�U�|�P�@�q�greticular formation Olszewski �� Baxter (1954)
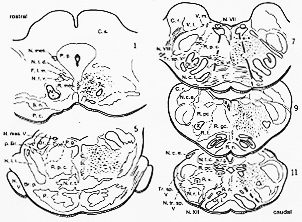
FIG. 6-2 A cytoarchitectonic map of the reticular formation of the cat. In a series of transverse sections are plotted the various cell groups and their composition of small, medium-sized, and large cells. some abbreviations: Coe.: nucleus subcoeruleus; F.l.m.: medial longitudinal fasciculus; N.c.e.: exxternal cuneate nucleus; N.f.c.: nucleus cuneatus; N.f.g.: nucleus gracilis; N.r.: red nucleus; N.r.t.: nucleus reticularis tegmenti pontis; N.tr.sp.V: spinal nucleus of trigeminal nerve; P.: pontine nuclei; P.g.: periaqueductal gray; R.gc.: nucleus reticularis gigantocellularis; R.I.: lateral reticular nucleus (nucleus of lateral funiculus); R.mes.: reticular formation of the mesencephalon; R.n.: nuclei of the raphe; R.pc.: nucleus reticularis parvicellularis; R.v.: nucleus reticularis ventralis. From Brodal (1957).
�}�U�|�Q�@�l�Rreticular formation Brodal (1975)
��זE��reticular formation����(�����2/3)�ɂ���B���A���ƍ����āA
�O��1/3�͏��זE�̂݁B
�܂���זE�͂��郌�x���ɓ��ɂ݂���(�}�U�|�S���݂�)
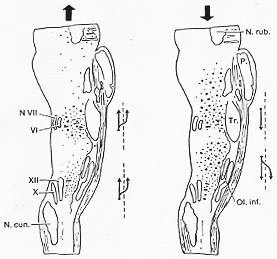
FIG. 6-4 The distribution of cells of the reticular formation sending long axons to the spinal cord (right) and of cells having long axons ascending beyond the mesencephalon (left) is mapped in parasagittal sections of the cat's brainstem. Large dots indicate giant cells. Note that, although both types of fibers take origin from paracctically the entire longitudinal extent of the reticular formation, they both have their maximal regions of origin. In spite of some overlapping, these maximal regions are different. The arrows to the right of each drawing indicate that the pontine reticulospinal fibers descend homolaterally, while all other contingents are crossed as well as uncrossed. From Brodal (1956).
R.gc. gigantocellularis (����)
R.p.c. pontis caudalis�@��R.p.o.�b���ڍs(����זE������)
Golgi����
�@Scheibel �� Scheibel (1958)
�@Leontovich �� Zhukova (1963) �g���ׂ�axons�͑O������ɑ�������@���h
�@�T�^�I��Golgi II�זE�Ȃ��@(�Z�����A���}�A��݃j���[�������͘A���זE�̌��^prototype)
�@���b�g��Valverde (1961b)�͊O���̏��זE���̈�ɋɂ߂ď�����Golgi II�^���݂�B
�@���ׂĂ̒�������肢�����̑��}���o���A���q�p�^����l�Ȃ炴��@���A����ˋN�͒ʏ풷�����ˏ�ɂЂ낪��B������isodendritic�^(Ramon-moliner �� Nauta, 1966; ��)�B�قƂ�ǂ��A�]���̒����ɑ��Đ����ʂ�dendrite���Ђ낪�邱�Ƃ͓����I�ł���B�]���������ɐ_�o�זE���́|raphe nuclei (�D���j)
�@�敪�|Taber, Brodal �� Walberg (1960)
�@�זE�̌^�y�ё��̂������̓_�ŖD���j�͖ԗl�̂Ɏ���B
�@PPRF(paramedian pontine reticular formation)
�@reticular formation�̔w���ɂ������̎U�ݐ��ɋ敪�����̈�Ő�����horizontal gaze�̃R���g���[���Ɋ֘A(�V�͂�g.�Q��)
�gthe reticular core of the brainstem�h
�@reticular formation�Ǝ�̑��̍זE�Q���܂Ƃ߂Ė����Ƃ��Ď�̌����҂͂��̖��̂̎g�p���D��(���_�ɂ��Ă�Scheibel 1980)���݂�B
�@���S���@�ہF�Ґ��A�����A����є]�����y�т�蕫���̑��̏��j�ցB
�@reticulospinal tracts (�S��. p.205).
�@�}6-4. ��ɂQ�̈�(pons(����ɓ�����) �� medulla(��������)), ����2/3 of reticular formation���Ƒ�זE�B�Ґ��̉^���j���[�����Ƃ̓V�i�v�X������������VI-VIII�ɏI���B
�@���s���Ɠ������I��I�N�n����(�Q��)������B��2/3����P�͉����A���͋������|�����㕔(�Q�̋���זE�͏�s����(���s�@�ۂ̂ݏo��)�Ґ��ւ̉e���̂�)�B������(�}6-4)(Brodal �� Rossi, 1955)(��Golgi �� Nauta�@��)��q�̈�̏��Ȃ��Ƃ�1/3�̍זE�͒��]���O����axon���o���@���B�����A���]reticular formation������s�@�ۂ���(Nauta �� Kuypers, 1958)�B����̏�s�@�ۂ��A��́g�����j�h�ɏI���(cp. 6-12�}�A430��)��������s�@�ۂ̏I�~�Ɋւ��Ă�open question��B���]reticular formation�����������Apreoptic area, �������u�j�A�X�ɔ���j����Y�j�A���̒��]����̑@�ۂ͎������ђʂ������̕�����ʂ�B���̋�������_�͌�q����B�ȏ�̏����͓t��@�ő�̊m���߂�ꂽ(Lynch et al., 1973)(���ABowsher, 1975)��reticular formation�ȉ��̏��͑@�ۂł͑@�ۂ͎������j���ĕ����֍s���Ȃ��B������������̏��ł́A��̑@�ۂ�e.g.����j�A�O���t�ɂ������ǐՂ�����B
reticular formation��s�@�ۂ̏��Ȃ�����̂��̂̓R�����쓮��(Shute �� Lewis, 1967)���A�Q�̖L�x�ȃm���A�h���i�������̏�s���˂���(�]��reticular formation����)(Fuxe,��A1970)���̃R���������m���A�h���i�������͏�s�����n�̈ꕔ�ƍl������B
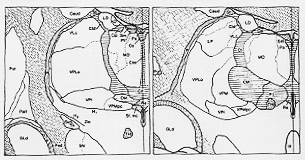
Fig. 6-12 Drawings of transverse sections through the thalamus of the macaque monkey, showing the arrangement and subdivisions of the "nonspefic" thalamic nuclei. The reticular nucleus is indicated by dots, thie intralaminar and midline nuclei by horizontal hatchings. The section to the left is placed most rostrally. Some abbreviations: Ci: internal capsule; Cif: nucleus centralis lateralis; CM: centromedian nucleus; Cs: nucleus centralis superior; Csl: nucleus centralis lateralis; CM: centromedian nucleus; Cs: nucleus centralis superior; Csl: nucleus centralis superior laateralis; GLd: nucleus geniculatus dorsalis; LP: nucleus lateralis posterior: MD: dorsomedial nucleus; Pa: nucleus paraventricularis; Pf: nucleus parafascicularis; R: nucleus reticularis; Re: nucleus reuniens; Sf.mc: nucleus subfascicularis pars magnocellularis; Sm: stria medullaris; SN:: substantia nigra; St: stria terminalis; THI: tractus habenulointerpeduncularis; VLc: nucleus ventralis lateralis, pars caudalis; VPl: nucleus ventralis inferior; VPLo: nucleus ventralis posterior lateralis, pars oralis; VPM: nucleus ventralis posterior medialis; VPMpc: nucleus ventralis posterior medialis, pars parvocellularis. From Olszewski (1952).
������s�A�y�щ��s���˂̉�U�w�I����
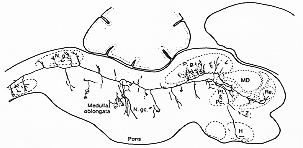
FIG. 6-5 Drawing of a sagittal Golgi section from the brainstem of a 2-day-old rat, showing a single large cell in the magnocellular nucleus. It emits an axon which bifurcates into an ascending and a descending branch. The latter gives off collaterals to the adjacent reticular formation (N.gc., nucleus reticularis gigantocellularis), to the nucleus gracilis (N.g.), and to the ventral horn in the spinal cord. The ascending branch gives off collaterals to the reticular formation and the periaqueductal gray (P.g.); it then appears to supply several thalamic nuclei (Pf. & Pc.; parafascicularis and paracentralis; Re: reuniens, and others), the hypothalamus (H), and the soccalled zona incerta (Z). MD: dorsomedial thalamic nucleus. From Scheibel and Scheibel (1958).
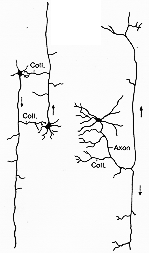
FIG. 6-6 Diagram illustrating two anatomical features which make possible a close correlation of caudally and rostrally directed influences of the reticular formation. To the right a simplified drawing of a typical cell in the medial reticular formation. Its axon gives off a long ascending and a long descending branch provided with collaterals (cf. Fig.6-3). To the left a diagram showing how cells gibeing off ascending and descending axonsm, respectively, may influence each other by way of collaterals. From Brodal (1964).
�@���s�A��s�C���p���X�݂��ɉe��������ڂ�(�}�U�|�U)(��)�B���ɖ��ڂ�integration������B
�@���}���A�]�_�o�j(���ׂĂ�)�Ɂ|�m�Ɖ^�Ɓ|�@����j�ɂ����}�s���@���Breticular formation ���@�O��_�o�j�ւ�(Hoddevik, Brodal �� Walberg, 1975).��Ƃ���R,gc.���]reticular formation����Ȃ��B
���S���@�� (reticular formation��)
�@�Ґ����瑽���A����͒��ڂ̐Ґ��ԗl�̑@�ۓ��͂őO������Ґ������H�ɍ����ď�s����B�����ёт̑��}�͏ؖ��ł������̓m�o�@�ۂɂ��activate�������̂͏��Ȃ��Ґ������H�̑��}�������ďd�v�łȂ��]���ł͔w���������ɕ�����A�I���}�Ƒ��}��reticular formation�זE�̑命����dendrites�Ɠ����ʁA�]�������ɐ����ɏo���B�������A����2/3�A�l�R�ŁAR.gc, R.p.c.�|R.p.o.�̂Q��ɑ����I���(Rossi, Brodal, 1957)�B����areas�͑�́A����s���@�ۂ̋N�n�̈�ɑ�������B���̂��Ƃ�reticular formation����ĐҐ����王���ւ̒��ړI�o�H�����邱�Ƃ������Ă���B���Anucleus subcaeruleus�ɂ������I���Ƃ����Breticular formation�ɓ���]�_�o�m�o�@�ۂ̃C���p���X�ɂ��Ă͂悭�m���ĂȂ����Ȃ��悤��(��ꎟ�m�o�@��)�B��m�o�@�ۂ́A�������Ȃ���A���̑��}��reticular formation�ɏo�����Ƃ�Golgi�Ŏ�����B�O���_�o�Ґ��H�j����A�O��_�o�j����R��B�O��(���)�ԗl�̑@�ۂ͔�r�I����ȃp�^������(Lapdli �� Brodal, 1968)�B�S�̎�j���o���B���������z��͈قȂ�B���R.gc.��Rpc.(PPRF)�|�������ዅ�^���ɂƂ�neural substrate(�V��)�B��s��acoustic�H�����}�A���o��impulses��u��reticular formation�@��u���������B
�@�k�o���C���p���X�͂悭�͔���ʂ��\�ȘH��Guillery 1956, 1957; Nauta 1956, 1958�ɂ�苓���B
�@�O�͂ŏq�ׂ������]����̑@�ۂ�����B���Ɏ����j����A����2/3�A���ɉ���reticular formation(Walberg, Pompeiano, Westrum, �� Hauglie-Hanssen 1962)���̑@�ۂ͑O�����ˈ�A�����ɋ�������B
�gHigher level�h�����
�@�O����������(Nauta, 1958,����тP�P��)
�@�W����(Johnson �� Clemente 1959; Nauta �� Mehler, 1966) �@��ɒ��]�ɏI���B
�@��u����A�Q��maximum termination area�玿reticular formation�@�ۂ̎傽��I�~��Ɉ�v����B�ԗl�̐Ґ��@�ۂ��o���Ɠ��肳�ꂽ��̃j���[�������܂߂āA�����̃j���[����
�@��u�h���ŒP�V�i�v�X���ɋ�������(Udo �� Mano, 1970; Peterson��1974)
�@�玿�ԗl�̑@��(���W�ԗl�̂���impressive)
�@2-6 msec�̒Z���������A(Hugelin et al., 1953)��ɒm�o�^���삩��A�玿�Ґ��H�Ƌ��ɉ��s���]���Ōo�ߒ�����𗣂�reticular formation�ɁB(�玿�ԗl�̑@�ۂ̂�����͔̂玿�Ґ��H�̑��}�����)�B��l�ɖ��ɏI���̂łȂ��Q��R.gc��Rp.c.-Rp.o. (Rossi �� Brodal, 1956a).���̓����ł����llateral parvicellular reticular formation�ւ�(�ア���˂���F�T���E���b�g)���]reticular formation�ւ�(Szentagothai �� Rejkovits, 1958; Pearce, 1960; Valverde, 1962).
�@���������(�ԗl��)�����]�A���A�������x����reticular formation��(Rinvik, Grofova �� Ottersen, 1976; Jayaraman, Batton �� Carpenter, 1977).���ɂ͖D���j��������˂�����B
�@���܂Ō��ė������A�@�ی�������reticular formation��diffuse�U���ɍ\������Ă���̂ł͂Ȃ����Ƃ��킩��B�������̎傽�鋁�S�@�ۂ̏I�����Ƃ������S���@�ۂ̋N�n���Ƃ��͎�Ȃ镔�ʂ����Ȃ�n�b�L���ƌ��܂��Ă���B�����I�~���N�n���Ƃ��͓���j�Q�ɃL�b�J���ƑΉ����Ă����ł͂Ȃ�����ǂ��B���Areticular formation�̓���2/3�ƊO��1/3��Ƃł͒��ڂ��ׂ����ق��݂���B�O��1/3�͏��זE�݂̂ŏ�s�@�ۂɂ��Ă����s�@�ۂɂ��Ă������͔�r�I�Z���B����2/3�����������̑�^�זE���܂݁A����̐��̏�s�y�щ��s�@�ۂ��o���Ă���B�]���āAreticular formation��effector�@�\�ƌĂԂ��̂́A��Ƃ��āA���̓���2/3�ɂ���ĉ��Ă���A����O��1/3�͍זE�����ɐL�т�axons�ɂ��A�啔���O��(����2/3)�ɉe����^����悤�Ɏv����B����reticular formation�̊O�����͂�����loosely�ɘA���̗̈�Ƃ�����Ƃ���̋@�\���s���Ƃ������Ƃ͂��肻���ȉ����ł���B����2/3��ɂ́A�X�ɁA�s���S�ł͂��邪�A���x���Ԃɕ������݂��(segregation)�B���郌�x���͎傽��e����Ґ��ɗ^���A���̃��x���͔]�̂��ƑO����act����(�U�|�S�}���݂�)�B������reticular formation�̌`�Ԋw��̍\���́Areticular formation�͋@�\���̎U�����ɍ\�����ꂽ�P��entity�ł���Ƃ��鉼���ɋ������_�ƂȂ�B
�@�זE�\�z��@�ی����̌��������ł͈ꕔ�I��Golgi�������d�v�B����ˋN�p�^���Ǝ����̕��q�Ȃ�Golgi�����Ŗ��炩�ɂ�������������Breticular formation�̓T�^�I�זE�͉��蒷������ˋN�������Ă���AGolgi�W�{�ŁA�זE�́hdendritic fields�h�������ɏd�����Ă��邩�����Ɍ�����B�X�ɈقȂ��X�̋��S�n�ɑ�����@�ۂ̑��}���L���͈͂Ɍ݂�d�����Ă��邱�Ƃ����邱�Ƃ��ł���BScheibel �� Scheibel (1958)�ɂ�苭�����ꂽ�ʂ�A����(�d��etc.)�͓�����(reticular formation) �ŗD���ɂ݂���B�����Ă������璷����s�y�щ��s���ˑ@�ۂ��N����̂ł���B��������̂��̏d���́g������͂������ɕۂ���邩���݂邱�Ƃ��ނÂ���������Ă���h(Scheibel �� Scheibel, 1958, p.34)�B
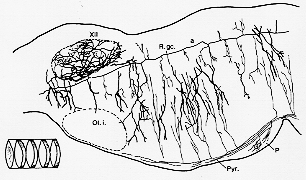
FIG. 6-3 A drawing of a Golgi-impregnated parasagittal sectioon through the lower brainstem of a young rat, illustrating the orientation of the dendrites of cells in the reticular formation in planes perpendicular to the long axis of the brainstem. Collaterals of axons descending in the pyramid (Pyr.) take off in the same plane as do also the collaterals of a long axon (a) of a reticular cell. (Note different organization of the dendrites in the hypoglossal nucleus, XII.) Ol.i.: inferior olive; P: pons; R.gc.: nucleus reticularis gigantocellularis. The inset at the lower left shows how the reticular formation may be considered a series of neuropil segments (cf. test). From Scheibel and Scheibel (1958).
�@Reticular formation�͎U�����ɍ\������Ă���Ƃ���������ۂ�^����B�����̏���reticular formation�̍זE��dendrite���ߕӂ̊j�Ɋg����B�t���R��B���ׂĂ̔]�_�o�j��reticular formation, collaterals��Golgi II�����A(�命���̍זE�]���̒����ɉ����Ď������̂��BScheibel (1958)(p.42)reticular formation�̓��������ɂ��ċ�������F�����������B�g�ނ���A�]���̔��\���͈ȉ��̔@���Ȃ�ׂ������A�K���Ȃ鐶���w�I�������ł́A��̃C���p���X�p�^���́A���v�f�̑��������L���Ȃ邪�̂ɁA�����Ȃ�l�����邤��o�H�Ƃ�reticular formation���ɋ��炭�L�����悤�h�������Ȃ���Golgi�Ǝ�����������ނ�͋�����������ɒ��ڂ����Breticular formation��6-3�}�ɗᎦ�����@����A�̕��߂�萬��@���A�\���̌����͓����ł��A�e�g���߁h�̋@�\�͈قȂ�ł��낤�@��s���זE�A���s���זE�̕ʂ����x���ɂ��قȂ�B�A���S�@�ۃ��x���ɂ��قȂ�Breticular formation���S�@�ۂ̎傽�镪�z��ɂ��Ă̌�������
�@�]���āA�玿�A���](�����j����)��u�A�O��C���v�b�g�͍ł���Ԃ�(���ꂾ���ł͂Ȃ��ɂ��Ă�)�Ґ���reticular control�ɉe��������ڂ��Ă���ƍl���邱�Ƃ��ł���B
�@�����w�I�f�[�^�@��v����B
�@��X�̌�����̒m�o����reticular formation�S�̂ɓ��B����悤�ɂ݂��邽�߂�reticular formation�ւ̋��S���C���v�b�g��diffuse�ł���ƈ�v�ɉ]���邪�����ɂ͐������Ȃ��BScheibel, Scheibel, Mollica, Moruzzi (1955). Reticular formation��units��ɋ��S�C���p���X�̍L�͂�convergence���������Ƃ��Ă�����convergence��not unlimited���B
�����d�ɖ@��p��������
�@�זE���x���ł�reticular formation�̋@�\�ɂ��Ă̒m�����]�܂�邪�A���̏������Տ��ɂƂ��Ă͒��ڂ̊֘A�ƂȂ�B���̖��ɓ���O�Ɍ����ȈӖ���reticular formation�ɂނ��떧�ڂɊ֘A���Ă���R�̏���D���j�A���S�D�����A���j�ɂ��čl�@���邱�Ƃ��K�����낤�B
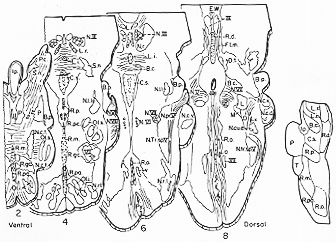
FIG. 6-8 The raphe nuclei of the cat, as seen in four equally spaced horizontal sections through the brainstem, to the right as projected on a midsagittal section. The eight nuclei distinguished differ with regard to cell types and cell density (roughly indicated by differences in size and density of dots in the drawings).Some abbreviations: B.c: brachium conjunctivum; C.s.: nucleuis centralis superior; lp.: nucleus interpeduncularis; L.i. and L.r.: nucleus linearis intermedius and rostralis; P: Pons; R.d.: nucleus raphe dorsalis; R.m.: nucleus raphe magnus; R.o.: nucleus raphe obscurus; R.p.: nucleus raphe pontis; R.pa.: nucleus raphe pallidus; T: trapezoid body. From Taber, Brodal, And Walberg (1960).
�@399�łłӂꂽ���A�זE�\�z���ʂ���邢������minor�זE�Q���琬���Ă���B��r�I���܂������Ƃ��A�����Ă��鉄�����[���璆�]�O���܂Ŕ]���̐������ɉ����ĂЂ낪���Ă���(raphe = seam�D��)�זE���͂ł���B��X�̈��j�Q�݂͌��ɂ��郌�x���ŗZ�����A�ǂ��ł�reticular formation�Ƃ͂����苫�E�Â�����Ă����ł͂Ȃ��B6-8�}�̓l�R�����f�ؕЂ̎傽��Ǎ݂������}�B�S���łW�̊j����ʁA��������O����
�@the nucleus raphe obscurus (R.o.)
�@nucleus raphe pallidus (R.pa.)
�@nucleus raphe magnus (R.m.)
�@nucleus raphe pontis (R.p.)
�@nucleus centralis superior (C.s.)
�@nucleus raphe dorsalis (R.d.)
�@nuclei linearis intermedius (L.i.)
�@nuclei linearis rostralis (L.r.)
cp. Taber, Brodal, �� Walberg (1986)���̓����A�q�g�A�������҂̂��́A�ד_�����A��v�A
�@��U�w�I�\���ƌ����Ɋւ��āA�D���j��main reticular formation�Ƃ������̗ގ��_�������B��X��raphe nuclei�͂�����x���ʂ̒P�ʂł��낤�B
�@�D���j(����R.d.��C.s.)�̓Z���g�j��(5-HT)�ܗL�זE���܂ޔ]�̎�v�������Ȃ�ׂ��@(Dohlstom �� Fuxe, 1964; Bjorklund, Falck �� Stenevi, 1971)�D���j�̉��S������(efferents)(6-9�}���݂�)Brodal, Taber �� Walberg (1960) (modified Gudden�@)�l�R
�@��N�@�t��@�AARG�@�A��s�����S�H�͖L�x�Ŕ��ɍL�Ăł��邱�Ƃ��m���ꂽ�B
�@���b�g(Conrad, Leonard ��Pfaff, 1974) �l�R(Taber, Foote��Hobson, 1976; Bobillier��A1976)�B�I�~(or��������)���ʂƂ���(��s��)
���]�̂������̍זE�Q(e.g.���S�D����)
���������̊j
�����j����ё��̊j(����)
���]
�G���̂̈ꕔ(�������)
�C�n�`���̂������
���u
����j�A��k
��]�玿�����F���ɑO���t
�@Falck-Hillarp�@�ő�̂Ƃ��ď�L��ɃZ���g�j���@�ە��z(Fuxe, Hokfelt �� Ungerstedt, 1970)�B���ׂẲ��S�@�ۂ��Z���g�j���쓮���łȂ���������ʁB����raphe dorsalis (R.d.) �� centralis superior (C.s.)����Ƃ���R.d.����N����B�O���t�ɂ�����(Lorens �� Guldberg, 1974).����raphe efferents�̓��ʂȖڕW�Ƃ��đ�]�̔]����
�@��ߍזEependymal cells�̕\�ʂ�cilia��microvilli�ɑ@�ہB�ׂ���varicose�̐_�o�@�ۃZ���g�j���܂ށB(Falck-Hillarp�@)�B��rostral raphe nuclei����A
���s���̉��S���@��(�D���j�����)
�@��s�������ʓI�ɑ����Ȃ��BARG�@�ŁA���E������reticular formation,�]�_�o�j�A���]�A�Ґ��֏����Aetc.
HRP�@��
�@����j�Ɣ�k �����R.d. (Nauta, Pritz, �� Lasek, 1974; Miller��, 1975)
�@�C�n �����C.s.��R.d. (Segal �� Landis 1974a; Pasquier �� Reinoso-Suarez 1977)
�@���j �����R.p.��R.d. (Sakai��A1977)
�@VI���t�AVII���t��raphes (Shinnar, Maciewicz �� Shofer, 1975)
�@Crus II. �����R.p.��R.o. (Taber, Hoddevik �� Walberg, 1977).
�@�Ґ�I, II, V�w�����R.m.��R.pa.
�@�����̖D���j�Ґ��@�ۂ����ԊO���זE��(intermediolateral all column)�ɏI���A�����_�o�ߑO�@�ۂɒP�V�i�v�X���}����p������B(Cabot, Wild �� Cohen, 1979)
�@raphe afferents���S�����������l�ɓ���p�^�����邪�@���Ȃ���̂��̗l�悭�m���ĂȂ��B�t��@��p����Brodal, Walberg, Taber (1960)�́A�Ґ��A���]�A��]�玿����̋��S�@�ۂ����ʂ���A�D���j���j���S���j�݂̂��������邱�Ƃ������o�����B(���̕��z�p�^���͂R�̌�����ő����قȂ�)�B�Ґ����S�@�ۂ̎匹�ł���nucleus raphe magnus (R.m.)�͂��ׂĂ���̍ł��d�v�ȏI�~�ړI�ł���悤�Ɏv����B���]�֓��˂���raphe pontis (R.p.)���A���]����̎傽�鋁�S�����͂��Ă���(���]�����R.m.�ւ��s����)���Ƃ͈Ӌ`�����낤(B.W. Taber, 1960; Miller �� Strominger, 1977)�B�X�ɒ��ڂ��ׂ��́Araphe dorsalis (R.d.)�͐Ґ��A��]�玿�A���]����̒��ڂ̓��˂͖w�ǎĂ��Ȃ����Ɩ�B�D���j�ւ̎傽����͂͋��炭�����O�q�ȊO�̕��ʂ��痈��B��������̗̈悩��̋��S�@�ۂ��L�q���ꂽ�B�����A���u�j�A�O��preoptic��A�O�����������A��j�j�A�O���O��A����������̂ǂ̕��ʂɏI��邩�͂����L����ĂȂ��B�������Ȃ���
Edwards (1975) ARG. ���]reticular formation��R.m., R.d.
Usunoff��(1975) ����j��R.m.
Chu �� Bloom(1974) ���j��R.d., R.p., R.m. C.s.
����
�ʑ��j��R.d.�̂�
���u�j��R.d.�ƥ��
�@���l�ɂ����̖D���j��reticular formation�y�є]���ɂ��邢�����̏��j�Ƃ̊ԂɖL�x�Ȍ���������BScheibel, Tomiyasu �� Scheibel (1975)
�@Golgi�@�AR.p., L.r.�Œ��ׁA�j���[���������w�����ɑ��錌�ǂɉ����ďW�c������Ă��邱�Ƃ������B�j���[�����̎���ˋN�A��(-)�A��������ɖD���j���[�����͌��t�����Ȃ���镨���ɑ���chemosensor(���w��e)�Ƃ��č�p���A�Ⴆ�ΐ����|�o�����Y���Ɋ֘A���邩������Ȃ��B
�@minor units�Ԃł͑傫�ȍ��Ⴊ����ɂ��s�S���A�D���j�����̂́A�傫���]���āA���(�P��)�{��entity; something with objective reality; an obstruction or arche-typal conception�̂悤�Ɏv����B����́A�L�͂ȉ��S�@�ۓ��˂ɂ���čL�͂Ȕ]�̗̈�ɖ��炩�ɉe�����y�ڂ��B�}6-9�ɊȒP�Ɏ����Ă���������B��s���Z���g�j���n���A�j�j�čs������(rat����)�B�^�������������A�s���ǁA���������i�A�U�搫etc.�c
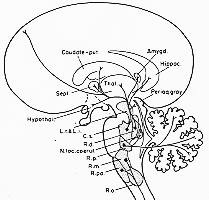
FIG. 6-9 A highly simplified diagram of the main efferent connections of the raphe nuclei. While many connections appear to come from more than one raphe nucleus, only the main nucleus of origin is indicated for each fiber contingent. For abbreviations of the raphe nuclei, see legend to Fig. 6-8 (cf. text).
�@�́A�זE�̋����F�f�����̌̂ɒ���(������nucleus pigmentosus pontis�ƌĂ��)�B���ׂĂ̚M���ނɂ���@���A��l�]���̑O���̏��̋߂��ɂ���B�O���_�o��m�o�j�̂��O��������[���n�܂�A�O���ɎO���_�o���]�H�j�̕�����(�ꕔ�זE�������Ȃ���)�̂тĂ���B���j�͎�ɒ��^�j���[��������\������Ă���A�זE���ɂ��e�ȃ����j�����������B�����A���^�̃j���[����(�ʏ탁���j�����܂܂�)���݂���B�����jnucleus subcaeruleus�́A���j�̕��O���ɂ݂�����U�ݐ��̍זE�Q���������̂ł���B�m���A�h���i�����ɕx�ޔ]���̈�ɑ�����B1964�N��Dahlstrom��Fuxe�̓��b�g�ł�����זE�Q��12, A-1�`A-12�Ɩ��t�����B�m���A�h����(�m���G�s�l�t����)Falck-Hillarp�@�ŗ̌u���BA6�����j�ɑ�������Bnoradrenergic neurons�𑽂��܂ރl�R�A�T���A���炭�q�g���A�������Ȃ���A���~���ׂ���NA�זE�̈ʒu�̓l�R�ł��T���ł����j�̗̈�Ɍ����ɑ�������̂ł͂Ȃ��B�Ⴆ�ΐ����j�⌋���r�t�߂ɂ܂łЂ낪��B������ɂ�荷�Ⴀ��B
�@���ɍL���A�g�D�u���@(����W�{�y�ѐ��j���͉��S�H�j���ɑ@�ۂ�ǐ�)�Ɖ��w�I���͂Ƃ̑��ւƂ���A�Q�̎傽���s��NA�_�o�H����߂�ꂽ�B�w���̌o�H�́A�D��ŁA�S��]�玿�A�C�n�A�G���̂��x�z����B�����Ď������p�jthalamic relay nuclei�ɂ�������̑��}���o���B�������͒��Ԃ̌o�H�́A�����������x�z����B���̑@�ۂ͏㏬�]�r��ʂ�A���]�֍s��(Olson �� Fuxe, 1971,��)�B���ꂩ������ւ̓��˂͖ԗl�̂��܂߂ĉ����]���ɍs��(Olson �� Fuxe, 1972)�B�P�̃m���A�h���i���������b������ɂ��A�S��]�玿������������(Olson �� Fuxe, 1971; Maeda �� Shimizu, 1972)�B�I���}�͂����ւ�ׂ��Bvaricosities�����B����HRP�ȂǁAARG.�c
�������ɍL�͂ȑ�]�玿��
�������ɑO����ʂ�A�����܂ŁA
�D���j�ւ��s���B
���S�������B�c�c�������������Ă��Ȃ��c�c
��������(�����ɏ���NA�זE���܂�)����E�T�M���j��(�i��ƒ���1970)EM.
�я������˂��� Domesick (1969)
���]���� Snider (1975)
�D���j(����R.p., R.d.)���� Sakai��(1977)
�������� Sakai��(1977)
�G���j���� Hopkins �� Holstege (1978)
�@���j�ƕt�߂̈�́A�D���j���l�ɁA���ɍL�͂ȉ��S�n�A�قƂ�ǂ��ׂĂ�CNS�ցB���������B���b�g�őS�̂�1400���Ɛ�������(Swanson, 1976)�B����Ƃ��āA�����n�̑�ȕ����Ȃ��B���A���Ƃ��A�玿�̊o��cortical arousal, �t������paradoxical sleep�ւ̓���(Jouvet 1972���݂�)���������ǂɑ��鈳�E������mediate���߂���B(�Ґ��ւ̓��˂Ɖ��)(Ward �� Gunn 1976)
�@Nissl�W�{�ŋ�ʂ���鏊�Ȃ�������E��U���葽���A��r�I���ȁA��Ƃ��ď��^�́A�זE��萬��B�������ɋ敪,�����A�O���A�w��(��)�j(Hamilton �� Skultety, 1970; Hamilton, 1973a).�q�g�ł�Olszewski �� Baxter (1957)�ɂ�鎦���ƈ�v�B
(���S�H��)
���������W�ցA�O����Forel��A(VTA)�ցA
�w���j���瓯���̎��W��A�O����j�j�O���j����A�㕔���������A�������̎����j�F���I���[�u�j(Walberg, 1965, 1974a).
�@�����͏��Ȃ���(���S�@��)�������j�A�w���j�ƈقȂ邪�\������B(�S�̂Ƃ��Ă݂��)�я��A�O���O��A�C�n�A���u�j�A�O�����������A�r�Ԋj����A���ɐҐ��Areticular formation���獕���A�s�m�сA�����ԗl�j�A��������(VM�j)�B
�@�����w�I�ɂ́A�{�蔽���A�H�a�����A�N���g�[�k�X�ւ̉e���A�ɂ݂Ɋ֘A�A���S�D������d�C�h������ƁA�N�Q�h���ɑ��锽��������������B���̌��ʂ́A�ɂ݂�`����Ґ��j���[�����ɑ���}��(����)action�ɂ����̂ƈ�ʂɍl����Ă���B���S�D��������Ґ��ւ̒��ړ��˂͂Ȃ��悤�����A�D���j�Ƃ���raphe magnus (R.m.)�̎h���Œ��ɍ�p�������N�����̂ŁAc.g.��R.F.��spinal cord�Ǝ��������B
�@���S�D�����͖D���j�Ƌ��ɁAmorphine-induced analgesia�Ɗ֘A����Ɛ�������Ă���B�]�h���ɂ����ɂƃ����t�B���ɂ����ɍ�p�Ƃ̊Ԃɗގ�������Ɖ]���Ă��A�����̃��J�j�Y���Ɋւ��Ă͑����̓_�łȂ������̈���o�Ȃ��B
�@�����̋@�\���ԗl��(reticular formation)�ɂ��e���������邱�Ƃ��m���Ă���B�����A���]�̔@���A�]�̑��̕����Ƃ̑��������l�����Ƃ��A�_�o�n�ɂ���ăR���g���[������Ă���قƂ�ǂ��ׂĂ̋@�\�ɑ���reticular formation�͏d�v�Ȃ���̂ł��낤�Breticular formation�̋@�\�I�����̂������̑��ʂ������ōl�@���Ă݂悤�B��s�������n�̊T�O�ɂ��Ă͎��߂ŏq�ׂ�B�����ł́A��Ƃ���reticular formation�́g���s���h�̍�p���Ȃ킿�A�̐��A���������ɐҐ����J�j�Y���ɑ���e���ɂ��čl���Ă݂悤�B
�@�������]�����A�̃g�[��(�ْ�)�A���ˁA�ċz�A���̎����@�\�Ƃ��ɐS�����ǃR���g���[���ɕK�v�Ȃ邠��ꕔ�̕��ʂ������Ƃ��m���Ă����B�����_�o�n�̍\���Ɋւ����ʓI�������O����Ƃ��āA40�N����O�́A���̂悤�ȋ@�\�̃R���g���[���Ɋ֗^�������́g���Scenter�h��]���������Ă���Ɛ��ʂ���Ă������Breticular formation�̓��蕔�ʂ��h�����͔j�ĉ^�������A�����A�ċz�́���������I�ɒ��ׂ�ꂽ�B�ǂ�area���Ƃ����悤�ɁB��ł��̒��S��(�y�A)�͋�ԓI�Ɉ�v����B���Ƃ��A�ؗ}���|���Ǘ}���|�z�C�|(���Ƃ���Test-figure 17, Brodal, 1957���݂�)�B
�@��N�ɂȂ��Ĉȏ�͋ɒ[�ȊȒP���Ȃ邱�Ƃ�m��B�|���ݓI�ɝh�R�I�Z���^�[��|��������ċN����B�ω��������ɋN����BBach (1952)�creticular formation�����R���g���[���̕��G���@���G�L�x�ȑ�����������integration�͗v������B�u�ԁX�ŕω�����B�z���������q�ɂ���Ă��e������悤�B�Ƃ͂����Ă��A���̂��Ƃ�reticular formation�̈�蕔�ʂ����ƈႤ�����̓���̂���@�\������Ƃ����\���܂Ŕr�����Ă�����̂ł͂Ȃ��B��{�I�ɂ́A�]�̈�Ԃ̂��ׂĂ�cooperation�����́A���ځA�Ԑڂ̑������Ɉˑ����Ă�����̂��B�m���͂������Ȃ���A�f�ГI��B
�@Magoun �� Rhines (1946), Rhines �� Magoun (1946).����reticular formation�̕��������}����(myotatic reflex �� muscle tone)�c���]�����ɂ�����d�������l�����o��flaccid�����L�̗}��������O�����A����ѕ����̋��E���]�������̐����j����ђ��S�D�����A�������j�A���������c���ʈ�B
�@�`�Ԋw�I�ɂ݂ċ���������̂͂���Magoun�̗}����́A�ԗl�̐Ґ��@�ۂ��o�����Ґ��̖�Ɖ���悭��v����B���̗̈���h������ƁA������ѐL�W�^���j���[������IPSP���L�^����B(���Ƃ���Jankowska, Lund, Lundberg, �� Pompeiano, 1968���݂�)�B
�@���i���ʂɊւ��Ă͂����ƕ��G�Ȃ��́B
�ŏI�X�V���F2002/09/13
