| 第7章.脳神経 |
要約:慶應義塾大学解剖学教室 川村光毅
・脳神経一般
・(A)舌下神経
・(B)副神経
・(C)迷走神経(X)と舌咽神経(IX)
・(D)前庭神経(VIII)
・前庭器官と前庭神経と前庭神経核
・前庭神経核の繊維結合
・眼振-機能的側面
・(E)中間(神経)顔面神経
・(F)三叉神経
・(G)外転・滑車・動眼神経
・眼球運動
・動眼核への求心性結合
・副動眼神経核、視蓋前域、上丘
・視蓋前域

| 脳神経一般 |
進化の過程で、身体頭部(前方部)は、第一義的に特殊感覚器官の発達のために、不断に重要さを増してきた。これが神経系の構造の内に反映されている。身体前方部の優勢なる影響は大脳皮質の発達と共に、ピークに達する。この大脳皮質は、とくに高等哺乳類やヒトで、容積のみならずその構造上の複雑性において、神経系全体の機能に対して(とって)圧倒的な重要性を有する器官である。しかるが故に、脳幹の原始的primitiveパタンはその尾部、つまり延髄において、最もたやすく見分けがつき最も完全に保たれている。
髄脳と後脳において、境界溝sulcus limitansがヒトおよび発育が完了した脳にみられる。
これは第4脳室底ではっきりみられる。脊髄の発達における機能的意義に従えば、この溝は、脳幹においても、基板(basal plate, 註)原文のfloor plateは誤りというよりbasalとしたほうが良い)における遠心性核の帯域と翼板(これはここ-延髄-では外方に向かうが)における求心性繊維の終止域の帯域とを境する。胎生期の発育のときは、遠心性および求心性の諸核(を与える)となる細胞は、縦柱状に配列しているが、後になって別個の明瞭な細胞群即ち核として部分的に分離するようになる。しかしながら、その大部分は大体もともとあった場所にどどまる。且成人においてさえ、それ故に、異なる一次(原始)柱から分かれた核は、柱状様式の配列をとってみられる。
脊髄では4つの機能上分離した範疇繊維のものがみられるにすぎないが、すなわち、
身体の前方部では、特殊感覚special sensesと鰓弓branchial arches (or咽頭弓pharyngeal arch)を有する鰓器官gill apparatusが発達しているため、これにともなって繊維の種類(カテゴリー)神経核が相応して複雑となっている。ある脳神経核は遠心性のみ
他は求心性のみ
他は温合性
と一様ならず(次頁に7-1図を示す)。
[甲]遠心性の最内側の核(体性遠心性核somatic efferent)前角細胞と密接に関連あり、発生学的に連結したものである。大切の筋板myotome由来の横紋筋支配頭部体節由来の筋は
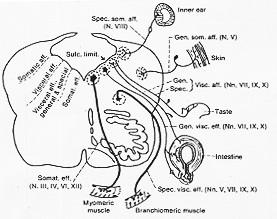
図7-1。XIは省略、Strong と Elwyn (1943)改。形態的に前角細胞と同型で第4脳室と中脳水道の床のすぐ腹側に位置する。
[乙]内臓遠心性核(visceral efferent nuclei)2カテゴリーあり、(A.B)やや腹方に核が移動した断続した柱、横紋筋へ(鰓弓の中胚様由来)。
第1鰓弓由来の咀嚼筋(+顎二頭筋M. digastricusの前腹と顎舌骨筋M. mylohyoideus)
第2鰓弓由来の顔面・表情筋(+顎二頭筋の腹筋の後腹と茎突舌筋筋M. stylohyoideus)
第3,4鰓弓由来の咽頭・咽頭筋
V,VIIの運動核、疑核(IX,-Xの一つ)と恐らくXI
同様のタイプの核、ときに“外側or特殊体性遠心性の核および繊維と呼ぶことあり。
境界溝の内側に在る。
E.W核(III)
上および下唾液核(VIIIとIX,各々)
背側運動核(X)
形態上、脊髄の交感神経性の中間帯外側細胞柱intermediolateral cell column(側角細胞)に似る機能上は副交感系の脳部分である。節前繊維を出しこれは自律神経節に終わる。
節後繊維は支配領域の平滑筋と腺に行く。(例えば涙腺とか唾液腺)。
[丙]内臓性求心性visceral afferent
境界溝のすぐ外側にある知覚性の細胞柱でただ一つの核・・・弧束核(延髄全長に見られる)。
VII(中間神経)
IX
X
それぞれのNNに神経節があり(この中にsomaあり)弧束核に入る前に下行する(弧束tracts solitarius)
弧束核に入る通常繊維に通常2つの型が区別され、一定の形をとった相当せる亜核を示す(Barnard, 1936)。
[丁]体性求心性somatic afferent
翼板の最外側、これに2型あり、即ち、一般(general)と特殊(special)顔面の表在(恐らく深部も)知覚を与える。(V)延髄を少しの距離を下行したのちに核に入る。主知覚核と脊髄路核細胞体は半月神経節内にV核と後角類同、V, tract nucleusは後角の膠様質(gelatinous substance)につづく、X,IX,VII(中間神経)の内にもこのタイプの繊維が若干ある。
VIII(前庭と聴神経)内耳からの感覚インパルスを伝える。前者が固有知覚性インパルス後者が外部知覚性インパルス後者が外部知覚性インパルスこれらの核は延髄の最外側と背外側部にある。右の7-4図にみる如く、運動核(遠心)のほとんどが分離しているが、知覚核(求心)はそうでなく連続している。とくに明らかであるのはV,VII,IX,Xをうける三叉神経核VII,IX,Xの内臓求心繊維をうける弧束核これは多分に機能的意味がある。Kappersこれを形態発生(形成)における一般法則の現れであると解釈した。すなわち、同一感覚様態を伝達し、同時に通常刺激をうける繊維群は、CNS中で同一経路をとり同一核群に終わる傾向がある。
脳神経一般を考察するとき忘れざることは、単に大脳皮質の働きである意識活動に関連した知覚と運動のメカニズムの連結であるのみならざることである。同様に、脳神経は、頭部とその諸器官の持つメカニズムを通して発現又は一部発動される非常に重要な(生命維持に必要な)多くの反射(機構)において演ずる役割も意義のある大切なもの也。
介在ニューロン網様体を介するものや相互結合etc.
側枝、運動核と知覚核
7-3図
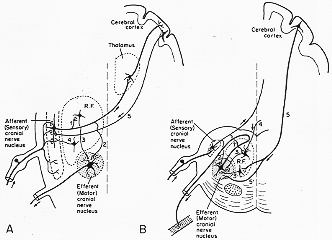
FIG. 7-3 Daigrams illustrating some principal features in the organization of cranial nuve nuclei. A: The neuronss in an afferent (sensory) cranial nerve nucleus (for example the trigeminal nucleus) send their main axons centrally to the thalamus (from which a new set of neurons projects to the cerebral cortex). The ascending fibers from the nucleus give off collaterals (1) to the reticular formation (R.F.) and to motor cranial nerve nuclei (2), which in addition may be acted upon by afferent impulses via the R.F. (3). (Routes 2 and 1-3 represent reflex arcs.) The sensory nucleus may be influenced both from the R.F. (4) and from the cerebral cortex (5). B: The neurons of an efferent (motor)cranial nerve nucleus (for example, the nucleus ambiguus) send their axons to the periphery. Its cells are acted upon by collateraals of ascending sensory fibers (2)and by axons (3) from cells in the reticular formation (R.F.), descending fibers from the cerebral cortex (5), as well as from other sources (4). Cells in the reticular formation sending axons (3) to the motor nucleus may further be influenced from collaterals of ascending fibers (1). In man some cortical fibers (5') appear to establish monosynaptic contact with the motoneurons.
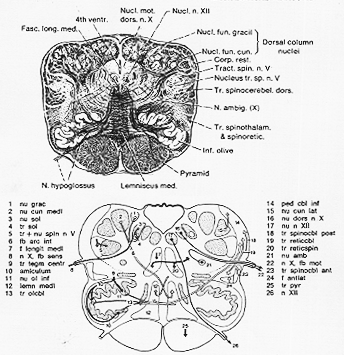
GIG. 7-4 Above, a drawing of a transverse section through the caudal part of the medulla oblongata in man. Weigert's myelin sheath stain. The fibers are clearly visible, the nuclear groups evident as light areas. (Approx. ×4.) Below, a schematic representation of nuclei and fiber tracts of the medulla oblongata (taken from Niewenhuys, Voogd, and van Huijzen, 1978) at approximately the same level as the Weigert section above. Exxplantations on the sides of the figure. (Some of the names are not used in the text.)
| (a)舌下神経(p.453~) |
解剖:
体性遠心グループ、脊髄の運動ニューロンと同型。多くの動物で、又ヒトでは胎生期に前角と連結、この神経核は縦柱配列、
perihypoglossal核
XII神経には繊維を送らず他の投射(5章をみよ)。
区分-亜核-種々の舌筋(Barnard, 1940)
求心性入力
大脳皮質から、ヒトでは若干の繊維が細胞に直接終わる(Kuypers, 1985c)。主に対側(交叉)性入力。reticular formationからも、V知覚核、弧束核、反射性吸飲、燕下、咀嚼の作用にあづかる。
XII.静脈叢にとりかこまれて後頭骨の舌下神経管を出る。
内舌筋のみでなく styloglossus 茎突舌筋…を支配
hyoglossus 舌骨舌筋…典型的運動終末
genioglossus オトガイ舌筋…舌の半側に限る。
(舌骨上筋の一つ)→geniohyoid オトガイ舌骨筋…運動単位は小
舌を突出させる傷側に曲がる。
vasoconstrictorも混じる(SCG or carotid plexusからのもの)。
延髄根と脊髄根 →頸静脈孔で合体(内枝を出してこれはX下神経節上端でXに合体)(残り外枝がXI固有。)
胸鎖乳突筋と僧帽筋を支配
副神経核(C2~C5(6))から外枝(external ramus)
体性遠心性タイプの細胞柱。尾方は前角の背外側部の細胞群。吻側方は、或る程度は独立性を達し、疑核の尾端に近づく
[註3]この故に(この記述はそれらの研究者からとったものだが)、副神経は特殊内臓遠心性グループに属し、胸鎖乳突筋と僧帽筋は鰓性間葉(中胚葉)組織由来という人もいる。XIは前根と後根の間でligament denticulatumと背側で側索より束として出る。上行して大後頭孔(foramen magnum)を通り -脊髄根として合体- 延髄根と頸静脈孔(jugular foramen内で)で合体し下行し、内枝を分離したのち、残りは外枝として下る。この途中C3-C4枝が入り、これらは主に僧帽筋の一部へ遠心運動性繊維の他に副神経は、いくらか求心性繊維を有す。固有感覚性らしい。
症状
sternocleidomastoid筋-萎縮すると、傷害側と反対側に向かって頭がまがり、顎が上がる。Trapezius傷害
肩甲骨がやや外側に位置がずれる。そして多少ゆれ動く。肩甲骨が正常に運動せず(Slowing?)
| (c)迷走神経(X)と舌咽神経(IX)(p.460~) |
共に以下を含む。
特殊内臓遠心性 …疑核(X,IX)
一般内臓遠心性 …X背側核(X)所謂下唾液核(IX)
一般体性求心性 …N.V核
特殊内臓求心性 …弧束核。(X,IX)
一般内臓求心性 …弧束核。(X,IX)
疑核…XII核と同型で柱状配列、移動した位置、大部分Xへ、少しIX., XI(内枝、再合する。)咽頭、喉頭、食道上部の横紋筋へ、区別在り(小印刷されている)。
迷走神経背側運動核
縦柱配列、第4脳室底。XII核の外側。下唾液核(IX)
散在性分布、節前繊維、耳下腺と平滑筋、(11章で後述)
IX,Xの求心性繊維の大部分は内臓性タイプ(一般及び特殊内臓性感覚のインパルスを運ぶ)。特殊内臓求心繊維(味蕾繊維)は一般内臓求心繊維と同じく弧束核に終止する。下神経節(Xではnodose節状、IXではpetrous岩様部、or extracraniale外…)に在り。[cp. 上はjugulare頸静脈ggl. ; 上はintracraniale内ggl]→弧束→弧束核(束の外側に位置する)の尾部に終わる。若干がVsp核とreticular formationに終わる。中間神経の若干の内臓求心繊維が弧束に合し弧束核に。小型の神経細胞より成る。X運動核(dorsal)より可成り小さい。これよりの上行繊維は脳幹を上行し(主として核の尾部から)paraventricular, dorsomedial, arcuate視床下部の諸核へ、medial preoptic area、扁桃核中心核(下行繊維)同レベルのものは(同じく核の尾部から)
疑核、X背側核へ繊維を送る。相互結合として視床下部からこの節前性、自律神経遠心性ニューロンへの直接投射が最近証明さえた(Saper, Loewy, Swanson, と Cowan, 1976)。以上の結合は弧束核尾部が主であり、吻側部は味覚インパルスの中継に特に関連している。加え、HRP法で
弧束核 →背側縫線核(Aghajanian と Wang, 1977).
→背外側(部)、reticular formation(多数の繊維)*
→小脳(Somana と Walberg, 1979)
→脊髄solitariospinal (Torvik, 1957b; Norgren,1978;他)核の尾部より、大きい細胞、腰髄まで、(cp. Loewy と Burton, 1978).この核には、大脳皮質、脊髄、小脳の前庭小脳部その他から繊維が来ることから考えて単なる中継核ではなく。可成りの程度の統御integrationがなされると思われる。
*reticular formationの背外側部は、燕下、嘔吐(物)vomiting,心臓血管、呼吸調節などの多くの反射機能に関連している。
迷走神経の一般体性求心繊維→三叉神経核へ
耳甲介concha部の皮膚の支配Xの他にIX,VII(中間神経)からも若干量、V核にはいる(fの甲で詳述する)。
以上の諸核は種々異なる源からの繊維により活動化される。まづは皮質延髄路(一部交叉、一部非交叉)IX,X支配筋(咽頭、喉頭)(横紋筋)の随意的運動に関連、他に求心性の内臓及び体性繊維や、内臓および体性知覚核からの軸索や側枝が咳、燕下、嘔吐など反射行為(運動)における咽頭、喉頭筋の参加に介在する。更に、皮質下レベルからの下行繊維が、恐らく大部分reticular formationを介して効果(運動)核に影響を及ぼす。
IX,X脳を出、小束がIX,下部分の大部分がX.共に、頸静脈孔(F. jug)を通。
2つの神経節(典型的な-偽-単極性ニューロン)。
IX superior, petrosus
X jugular, nodose (2神経節間でXIの内枝が入る)
| (d)前庭神経(VIII)(p.470~) |
Pars vestibularis→前庭の球形のう(saccule), 卵形嚢(utricle)および(平衡斑)、半規管からの固有知覚インパルスを伝える。(平衡維持と空間における定位にとって重要なインパルス)この伝達されるインパルスは意識に上がる程度が低い。発生学上一次性耳胞からすべての(聴→両生類で始めて現れるも前庭器官→すでにサメに(生じ)よく発達してお降りすべての脊椎動物に見られる。も)生じる。
比較動物論状 脊椎動物の内耳は proprioceptive “equilibratory”
上部pars superior 主として内耳の固有知覚性“平衡”性機能
下部pars inferior 聴覚の外部知覚性exteroceptiveに関与。
前庭インパルスは究極的に、種々の反射結合(連絡(結))によって、主に運動器官に影響を与える。前庭器官は機能上、脊髄と小脳に特に密接に関連している。(第4、5章で考察した)。
| 前庭器官と前庭神経と前庭神経核 |
内耳道動に入り、内耳道底で種々の小枝に分かれて膜迷路の種々の部分へ至る。内耳道底を貫く所で、前庭神経節-原始型の双極性神経節細胞を持つ(聴神経同様に)。中枢側突起は前庭神経(→上行枝と下行枝に分かれる(延髄で)分かれる前は前庭神経は下小脳脚とVsp.路との間をとおる。枝は前庭神経核に分布(7-2,7-7,7-13と7-15図)を形成するが遠位の突起は非常に短く迷路の感覚上皮に終わる。
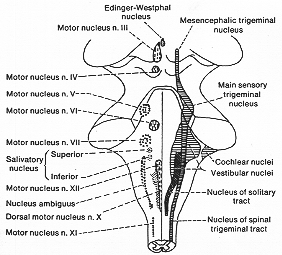
FIG. 7-2 Diagram indicating the collumnar arrangement of the cranial nerve nuclei. The nuclei belonging to the same categories are indicated by indentical symbols.
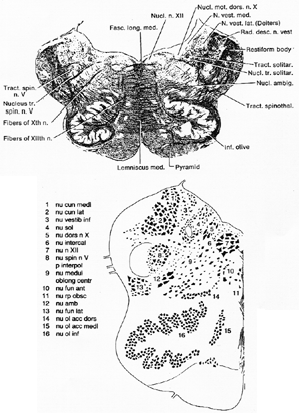
FIG. 7-7 Above, a drawing of a transverse section through the medulla oblongata at the level of the vagus nuclei. Weighrt's myelin sheath stain. Belo, a diagram (from Nieuwenhuys, Voogd, and van Huijen, 1978) of the nuclear groups in the medulla at a level somewhat beklow that of the Weigert section shown above.
核:主核は 上核(Bechterew)
外側核(Deiters)
内側核(三角核、Schwalbe)
下(行)核(下核、脊髄核)
他に小核群あり、
ネコ(Brodal と Pompeiano, 1957)
ヒト(Sadjadpour と Brodal, 1968)
細胞構築は諸核に異なり、同核でも部位的に差、特徴あり、(略)←1つの興味ある事実としてDeitersにみられる体性局在構成(4-7, 7-10図)
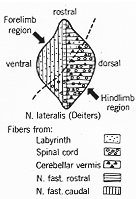 FIG. 7-10 A diagram of the lateral vestiblar
nucleus of Deiters in the cat in a sagittal view, showing the somatotopic pattern within
the nucleus and the approximate distribution within it of some of its main afferents to
particular territories. Note restriction of promary vestibular fibers to the
"forelimb region." From Brodal (1967b).
FIG. 7-10 A diagram of the lateral vestiblar
nucleus of Deiters in the cat in a sagittal view, showing the somatotopic pattern within
the nucleus and the approximate distribution within it of some of its main afferents to
particular territories. Note restriction of promary vestibular fibers to the
"forelimb region." From Brodal (1967b).
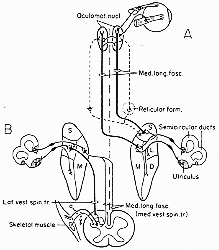
FIG. 7-15 Simplified diagrams of main featres in the organization of primary vestibular fibers to the venstibular nuclei and of the ascending (A) and descending (B) projections from these nuclei. D, L, M, and S: descending, lateral, medial, and superior vestiblar nucleus, respectively. For particularis see text.
一次前庭神経終止が“前肢域”に限局していることに注目。
前述の如く、前庭神経繊維は、前庭神経核に入った時に上行と下行枝と分かれる。これは(全部といえないが)ほとんどの一次前庭神経繊維が2分されるからである。この有髄繊維の多くは外側核に入り、そこから、上行繊維が上核(及び小脳)へ、下行繊維が下核と内側核へ至る。一般に信ぜられていたのと異なり、一次繊維は前庭神経核群の全域には終わらない。たとえばDeiters核では、前腹側部のみが一次前庭繊維をうける。これは生理学的にも確かめられた(Wilsonら, 1966)。(上核では一次繊維が終わるのは中心域のみで、且つ小群の若干のみが一次繊維をうける)。
明らかに、”vestibular nuclei”は一次繊維終止部に限られるべきだが、実際上の理由でこの術語をのこしておいた方がよい。
前庭器官の異なる種々の部位は機能的に同じでない故、迷路の種々の受容領域からの繊維の核内分布をみるのは興味あり、特定の終止域を重複しつつも有する。サル、ネコ前庭神経に遠心性の要素(成分)あり、(Gacek, 1960),数は少ないが、Rossi と Cortesina (1965a)によれば3つの小細胞群より、この遠心繊維は起こる。その1つが外側核内にある。HRP法でみると、この起始核は、
両側性、reticular formationの小域に、外転神経核の外側に、ダイテルス(外側)核腹側部の腹内側部にある(Gacek と Lyon, 1974; Warr, 1975)。前庭感覚上皮のすべての部位に終わる。前庭受容器の中枢性制御をmediateするなるべし、
(振動vibration receptorも)(位置の記録者)macula平衡斑。
前庭嚢は直線加速度的に応答する。静的迷路static; utriculus (+sacculus)、動的迷路kinetic; ampulla膨大部
三半規管角加速度に応答、(回転運動の受容器)。angular acceleration or deceleration
以上は単純化したものである。
半規管:”bi-directional angular accelerometers”
卵形嚢平衡斑(utricular macula):”position recorder”
一般に申して、前庭神経核は遠心求心繊維によって①脊髄②小脳③脳幹にあるある種の核群。④網様体と結合している。
前庭神経が脊髄の活動に及ぼす2つのよく知られた遠心性の経路。
外側前庭脊髄路
起始核は同側の外側核(大および小型細胞) 体性局在構成。仙髄レベルまで、RoxedのVII~VIII属。作用:伸展運動ニューロン(αとγ)に促通性作用
内側前庭脊髄路
起始核は主として内側核、交叉性と非交叉性。 胸髄上部レベルまで VII-VIII層(外側前庭脊髄路の終止域と同じ層内)。
以上のハッキリした下行路に対して弱い(modest)背髄前庭核投射がある。これはある程度に、DSCTの側枝であるかもしれぬ。そして下核、内側核、外側核の最尾域に僅少量終わる(restricted number)。ネコ、ヒトで多く(それらの大部分)が、x群(of BrodalとPompeiano, 1957)に終わる。これらの領野は、一次前庭繊維は僅かしか又は全く終わるものがない。
[註12]
脊髄求心繊維が終止するx群および他の前庭核領域は小脳に繊維を送る(Brodal と Torvik, 1957-恐らく主として前庭小脳へ(第5章参照)。これらの群は、補足的な、通常は無視されるような脊髄前庭小脳路の中継であろう。これに関して生理学上の何も知られていない。他の小部分(Vestibular complexの)について言及しておくz群は(Brodal と Pompeiano, 1957), x群と同様に、脊髄求心繊維を受けるが一次前庭繊維は受けない。視床を介して大脳皮質へのgroup I筋求心繊維(muscle afferent)の伝達にとっての一中継角であることが示されている(Landgren と Silfvenius, 1969; 1971)。
他方
y群は、Gacek (1969)によればsacculus球形嚢からの一次前庭繊維をうけるところであるが、生物学的にも(Highstein, 1974)解剖学的にも(GraybielとHartwieg, 1974)動顔神経核に投射することが結論づけられた。
生理学的研究(Wilson, Kato, Thomas と Peterson, 1966)によって、脊髄からのインパルスは他のルートで前庭神経核に伝えることが示された。主として、網様体を介してFredricksonら(1965)によれば関節の運動で特によく反応する。もう一つのルートは小脳を介するもので、前庭神経核に及ぼす(与える)脊髄からの影響についての複雑な問題の考察はPompeiano (1972b)をみよ。前庭神経核と小脳との関係についてはすでに第5章で考察した。いくつかの点をまとめてみると、
cp. vestibulocerebellar p.311
cerebellovestibular p.370 第5章
小量の繊維が前庭核から小脳に入り“前庭部”に終わる如し、これらの繊維は、内側核、下核くの内の限られた領域から(及びx群も)起こるが、ここは第一次前庭小脳繊維をうけないようだ(しかしながら5章をみよ。) cp. p.312 HRP法KotchabhakdiとWalberg (1978b)で以上を確かめた他にvestibular complexの他の部分からも繊維を送るとされた。又、片葉小節以外e.g. anterior and posterior vermisへも投射があることがハッキリした(Prechtら1977もみよ)。しかしながら、前庭インパルスは直接的に一次前庭小脳繊維を介して小脳にる。(5-10図をみよ)。即ち、小節、虫部垂の腹部、片様、(Dow, 1936)腹側旁片葉(Brodal と Hoivik, 1964)。(ネコ)サルも同様(Carpenter, Stein と Peter, 1972)らしい。がKorte と Mugnaini (1979)はネコでparaflocculusへの投射を確かめず。(5章をみよ)。このように前庭部を除いては、小脳は前庭器官により大きな影響はうけないようであるが、(逆に)小脳は前庭核に対して顕著な影響を与える。すでに5章でのべたが、“前庭小脳”は前庭核の一定のある部分へだけ少しばかりの繊維を送り返すだけであるが、小脳が前庭核に影響を及ぼすルートとしてより大きいものが2つある。両路とも虫部の一部から出て1つは直接繊維を他は、室頂核を中継する(7-13図をみよ)。特に興味ある観察として、外側核(ダイテルス核)への投射で5-30図(p.371)更に判ることは、この繊維を介して小脳の“脊髄部”(ここには前庭インパルスは僅かしか入らないようである)が脊髄の運動装置motor apparatusに対して体性局在的に限局した仕方でactするだろう。
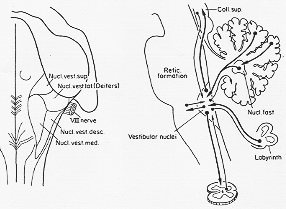
FIG. 7-13 To the left, a diagram of the four main vestibular nuclei in man as projected on the dorsal surface of the brainstem (cf. text). To the right, a simplified diagram of the main connections of the vestibular nuclei. Note that the vestibular nuclei have reciprocal connections with the spinal cord, the labyrinth, the cerebellum, the reticular formation, and the mesencephalon at the level of the superior colliclus (oculomotor and adfacent nuclear groups, see text). The connections indicated by heavy lines are nmore massive than those shown as thin lines. It can be seen that the major afferent input to the vestibular nuclei comes from the labyrinth and the cerebellum, while their main otput goes to the spinal cord and the mesencephalon (mainly the motor eye nuclei).
cp.小脳→前庭核Angaut と Brodal 1967図5-31.(373頁)。
前庭核とreticular formation
相互結合(Golgi法、Scheibel と Scheibel 1958).
Nauta法(Ladpli と Brodal, 1968)
Deiters核のみ→N. r. l.→小脳
主として
reticular formation(延髄と下部橋部)→VN(Vestibular nucleus complex)
このVN⇔reticular formationにより
前庭核は、前庭遠心繊維は長上行及下行繊維を出す領域に達するが故に、脊髄や視床のある領野の活動に影響を及ぼす。この前庭網様体結合は恐らく、前庭器官が刺激(irritation)されたときに観察される。嘔吐や心臓血管反応に関与する。
(fasciculus longitudinalis medialis) (Flm)
内側縦束を通る前庭神経核と外眼筋(ocular muscles)支配核との結合。これは前庭神経核と上部脳幹との結合内でとくに注目に価する。神経路ははっきりしているけれども複雑な結合系である。レベルは前方III核から尾方は脊髄まで拡がっている。系統発生的に非常に古い。個体発生上も早期に有髄化するので目立つ、正中の近く、第4脳室の下、外眼筋核と密に関連す延髄下部では腹方に移動し、前正中裂に近い位置で脊髄の前索へとつづく脊髄では普通sulcomarginal fasciculusと云われる。Flmには前庭核からの繊維の他に他の繊維が含まれている。これらのあるものは下行枝でCajal間質核(中脳)-III核近く-このあるものは前庭内側核の限られた部分(Pompeiano と Walberg, 1975)に終わり、他はinterstitial spinal tractとして脊髄に終わる(第4章、4-1図)。
Flm.の多数繊維は前庭核より来るものである。その一部が前述の如く内側前庭脊髄路として脊髄と至る。(図4-6,7-15b)。これらの下行路は内側核(下核から可能性ある)から起こるが、一方、上行路の起始核に関しては意見が一致しない。(7-15A図)。(for reviews see Brodal と Pompeiano, 1958; B. P. Walberg. 1962; Tarlov, 1967; Brodal, 1974)。Tarlov (1970), Gacek (1971)変性法 図7-16にTarlov (1970)の所見のサマリーをのせる。
7-15図 (A)上行路、(B)下行路。 上行路(Tarlov, 1970)
外眼筋の神経核(III,IV,VI)
cp. 新規の教科書:すべての前庭神経核は内側縦束に両側性に多数の繊維を与える。
cristae膨大部陵とmaculae平衡斑からの繊維が前庭神経核に終止する関連から云えば…cristaeからのインパルスは主に上核と内側核を介し
maculaeからのインパルスは一部内側核の吻側部と一部外側核の前腹側部を介する如し、半規管に自然刺激を与えると特定方向に眼球運動(眼振、後述)を誘発し、半規管からの神経を刺激すると、その規管の平面上に眼の共役偏位conjugate deviationが起こる(Fluur, 1959; Cohen, Suzuki と Bender, 1964; 他)。動眼神経の項でのべるが、共役眼球運動は両眼のいくつかの筋肉の正確な協同(を要求する)がなくてはならない。
個々の受容器からの一次繊維が終わる場所(核内)とVIII核(VN)の特定終止域から送る繊維のIII核内の分布との間に非常に正確な相関があり、III核内の外眼筋支配の細胞特定な配列。
基本的elementary3つのニューロン反射弓(Szutagothai 1952a, 1964b)(①一次知覚ニューロン ②前庭核ニューロン-③末梢性運動ニューロン)の結びつき。関節の前庭・動眼反射(VOR)(小脳、XII前位核、reticular formationの一部PPRFを介して)が関与。間接的VOR(前庭動眼反射)弓と眼球運動核へのそれらの影響(興奮性、抑制性その他)については(Cohen 1974; 又Precht 1978をみよ)の広範な総説をみよ。直接的三ニューロンVORの経路について若干述べよう。UemuraとCohen(1973)は前庭神経核の種々の部位を破壊すると、前庭動眼反応や姿勢に種々様々の効果を与えることを見いだした。
上核からの上行繊維は外眼筋核にただ抑制性作用のみを、一方内側の吻側部からのものは興奮性と抑制性とを共に介すると思われる(Cohen, 1974 for reference)。最近の生理学はもっと複雑なデータ。(petit略)
直接の前庭動眼経路
6, 直接、短潜時、興奮性
6, 直接、短潜的、抑制性
例えば水平半規管を刺激すると対側外直筋、動側内直筋を支配する運動ニューロンを興奮させ。
動側外直筋、対側内直筋を支配する運動ニューロンを抑制す。
Flmを上行する繊維は少量がIII核をこえてその付近の核に分布。Darkschewitsch核、後交連NPP, Cajalの間質核、最近のサルARG(Buttner-Ennerver と Lang, 1978)によればDarkschewitsch核へ終わるものは非常に少ないがCajal核には多数終わる。加え、前庭繊維は(いわゆる)内側縦束の“吻側核”にも終わる。垂直性注視にとくに重要(本章g.)。
前庭神経核から大脳皮質への路”polysynaptic”
古くは聴覚野の付近に、”vestibular cortical area”最近他の皮質域に、3野(Brodmann)のSmIの顔面域内の小域に、サルで後にウサギ、ネコでも。ここは、前庭インパルスとグループIA筋求心繊維と皮膚、関節求心繊維の末梢性体性感覚インパルスの収斂するところ。
意識にのぼる空間定位をさだめる(appreciation)、又、この前庭インパルスが錐体路細胞に影響し運動遂行の役割を演ずると仮定さる。VPLとVLに前庭反応、種々の形態学的研究から前庭核から視床のVL,VP(L,M,I…共に?)らしい(前庭核皮質路は視床由来ばかりではないだろう。他の系(小脳経由?reticular formation経由?)はあまりよく知られていないが。Darkschewitsch, Cajal核からの上行路の可能性もあろう。ネコ知覚運動野HRP注入でDarkschewitsch核に+(Avendano, 1976) 他に前庭神経核間の交連繊維がある(Brodal, 1972cをみよ)。機能的大きい意味がある。Ladpli と Brodal (1968)。4つの前庭核は交連繊維に関して大変異なる。e.g.(ネコ)外側核の間は結合は大変弱いが、上核と下核は強い。同名核以外の核にも繊維を送る。Tarlov(1969)もサルで似た所見を得。HRPでPompeiano, Mergner と Corvaja (1978).生理、総説Shimazu (1972).
Golgi法(Hauglie-Hanssen, 1968)介在ニューロンは少ないらしい交連繊維結合(Ladpli と Brodal, 1968)網様体前庭核路(Hoddevik, Brodal と Walberg, 1975)前庭小脳から前庭神経核へ(Angaut と Brodal, 1967, 図5-31, p.373をみよ)
| 眼振-機能的側面 |
卵形嚢(恐らく球形嚢も) “静的”static迷路 空間における頭の位置を記録する。
三半規管 “動的”kinetic迷路 空間における頭の核運動を記録する。angular movement
卵形嚢からのインパルスは主として身体の色々の部分の筋緊張の分布に影響を及ぼし、半規管は運動中に視覚の定位visual orientationが確保されるべく、主として眼の位置を調節adjustする。
“tonic labyrinth”緊張性迷路は前述した。第4章で除脳硬直(p.265)と緊張性迷路反射とその小脳の役割が述べられた(第5章)。眼球運動における迷路の役割を動眼神経との関連で再び考察しよう。
C1~3の麻痺で重篤な定位傷害、バランス欠、協調欠、など迷路摘出動物に似た症状を出すこのことは更に緊張性頸反応tonic neck reflex(4章考察した)の重要な役割を示す。ヒトでは前庭障害は動物における程大したことはない。迷路からの情報がないとさは、視覚性の定位と固有感覚の情報に頼る。視覚が奪われたり、凸凹平面や柔らかいマットレスの上を歩いた時にのみ、平衡を失う(Martin, 1967)。
眼振 迷路や前庭核の障害で出現することは可成りよく知られているが、脳の他の部位の傷害でおこる眼振については僅かしか知られていない。
眼振は前庭器官刺激で正常にみられる眼の共役運動又は特定の視覚刺激(optokinetic nystagmus視覚運動性眼振、視[線運]動[性]眼振、視運動性眼振、車窓眼振,section gをみよ)によりおこるものに対して用いられる。ここでいうのはvestibular前庭[性]眼振也。これは本質的には、角速度と減速度(angular acceleration and deceleration)(回転)の際におこる膨大部項(小帽, cupulla)の偏位(ゆがみ、片寄り)に起因した半規管の膨大部稜cristaeの感覚毛(平衡毛)の刺激に依存する。
Barany 内リンパの流動 endolymph
迷路誘発の眼振は、本質的には眼の律動的共同運動rhythmic conjugate movements of the eyes.臨床上、急速相と緩除相のうちfast componentの方向についてつけられる(命名あれる)が、これは全く便宜的なもので、実際には、緩除運動がactive phase能動相であり、早い相は原の位置にもどる反射に過ぎない。
軽度眼振…患者が急速(運動)相側を注視したときにのみ観察されるが
重篤眼振…注視部に関係なく起こる。
最もよく見られるのものは水平眼振であるが
垂直、斜、回転Nystagmus.もみられる。
自然におこる眼振はいつも(それだけで)病的なものである。健康人で種々な方法で誘発できる。(誘導眼振)。
回転後眼振postrotatory nystagmus.
温熱眼振caloric nystagmus.
小脳性眼振の存在するかどうかの問題(第5省をみよ)
この小脳疾患時に多くの場合に見られる“眼振”はいつも真の眼振であるとは限らず、眼筋の失調を解さるべきものである。しかしながら、前庭器官と眼筋との間の関連からみて、真の小脳性眼振を必ずしも否定し去ることもできぬ。忘れてはならぬことは、小脳傷害はしばしば(圧迫により)直接に脳幹を侵すか又は、二次的に(血管障害により)侵すもので、典型的な眼振はかくして前庭神経核への傷害によって引き起こされるのであろう。ヒトの小脳疾患で位置眼振positional nystagmusがよく起こる。(動物の実験でも)患者の位置を変えてみる。5章で述べたように、nodulus小節がこの関連で興味あり。
| (e)中間(神経)顔面神経The intermediofacial nerve |
特殊内臓遠心性繊維(表情筋へ)量的に最も重要であるが他に小部分顔面神経の知覚根と一般によばれる中間神経がありこれは内臓((特殊内臓)で味覚を伝えるもの)及体性求心繊維(少しあるらしい)と一般内臓遠心性(上唾液核より)とから成る。
茎乳突孔stylomastoid foramen出。耳下腺を貫く。扇状に拡。頭部の全ての表在性表情筋(第2鰓弓由来)を支配、他に①中耳のアブミ骨、②茎突骨筋sytlomyoid、③顎二腹筋後腹post. belly of the digastric muscle、④広頸筋platysma
VII亜核に、Vraa-Jensen, 1942, Courvelle, 1966b細かい運動-分化(核)。
顔面神経運動核に働くインパルスには色々ある。顔面筋の関与する反射には視覚刺激、聴覚刺激、並びに顔および口由来の知覚性インパルスにより起こるものもあり。この核内又は付近に終止する繊維は視覚の反射中枢である上丘、上オリーブ核(聴覚性インパルス)それに知覚性三叉神経核、弧束核からのものがある。たとえば強い光に為ったとき瞬いてついた閉眼したり、角膜にふれたとき閉眼(角膜反射、corneal reflex, blink reflex)したり、音の強さの変化に応じてアブミ骨筋を収縮させたり弛緩させたり(聴覚性中耳orアブミ骨(筋)反射)口に食物が入ったときかんじたり、吸飲したりすることは、これらの連絡によって行われる反射である。脊髄から顔面神経核に行く繊維をもtraceした(Kerr 1975).
この投射は両側性、主に頸髄レベルからVII,VIII,V層
単シナプス性、興奮性が主。
加え、顔面神経核は“運動領”皮質からの若干を含めて高位から影響を受けている。ネコではこれらの繊維は核に直接終止せず(Kuypers, 1958a; Walberg 1957)。大脳脚を刺激したときに又は多シナプス性に応答する(Tanaka, 1976)。ヒトでは若干が直接終わる如し(Kuypers, 1958c)。この皮質延髄路は恐らく顔面の随意運動を伝える。臨床所見から見て、眼のまわりとそれより上部にある筋を支配する亜核域に終わる若干の繊維を(これらのみ両側性!!)除いて、交叉性のように思われる。なぜならば、皮質延髄路の障害、たとえば半身不随の場合では顔面筋の下半のみが麻痺するから也。人脳の形態学的研究もこの所見を多少とも支持する(Kuypers, 1958c)。
他に高次レベルからの顔面神経核への求心路は赤核(Courville, 1966a; Edwards, 1972ネコ);Miller と Strorominger 1973サル;Mizunoら、1973,ウサギ)。で交叉性で特定域に終わる(perioral筋を支配する領域には終わらない)。その他、中脳の網様体から両側性に投射(Edwards, 1975)。他に、直接、多少関節投射であろうが、淡蒼球からあるかものしれないが、よく知られていない。これらの結合は情動的顔面(筋)運動に関連すると想定されている。
中間神経の構成は運動性VII神経よりも複雑也。小束で少く。末梢経過がこみ込っている。涙腺、顎下腺、舌下腺それに鼻腔と口腔の一部に在る腺に行く一般・内臓・遠心性繊維に関して云えば、それらは、節前繊維(副交感神経性、分泌性)繊維で読者は自律神経の章をみて欲しい。ここではわれわれは、中間神経の知覚性要素について考察する。
中間神経のsensory繊維は細胞体が膝神経節内にある。偽単極性ニューロンこの可成りの数の知覚繊維は、舌の前2/3から味覚インパルスを伝える。特殊内臓求心性繊維であろ。
中間神経の枝は、第1に大錐体神経Nerve petrosus majorという)(PNA)で(J.N.A.,BNAで大浅錐体神経Nerve petrosus superficial majorという)翼口蓋神経節ganglion pterygopalatinum, PNA, JNA[但しBNAで楔口蓋(神経節)ganglion sphenopalatinumという]へ行く。次に、鼓索神経chorda tympaniという枝となり鼓室内に入り…。そして舌神経と吻合して舌に達する。しかし、すべての求心繊維がこの2つの神経(大錐体神経と鼓索神経)に分布しているのではない。前述の如く、若干のもの(12-15%)は更にN.VIIを進む。以上の2つの神経の少なくとも可成りのものは味覚性(舌にもっと詳述する)であるが、一方、固有の顔面神経を通ものは層ではなく、一般・知覚・求心性繊維である。これは外耳に分布。耳介concha耳甲介および時には耳の後部の皮膚知覚支配である如し。(これらの領域は迷走神経の耳介枝によっても支配されている!)。
顔面筋反射(顔の知覚刺激に応じて顔面筋が収縮する)はN.V→Vsp.核を大部分が介して起こるものであるが(Iwata, Kitai, Olson, 1972),膝神経節に細胞体をもつ末梢性顔面神経中の求心繊維も、Willer と Lamour 1977)らの人による研究によって関与することもある。
顔面神経(中間神経)の内にと同様、舌咽神経、迷走神経にも若干の繊維が脊髄路三叉神経核に追跡できるものがある。これら3つの神経(VII, IX, X)は、かくしてしばしば述べてきていることとは反するが、体性求心繊維成分を含む。迷走神経では(若干は舌咽神経も)、すでに述べたように、外耳の領域の支配に関係し、一方、舌咽神経の大部分の繊維は舌の1/3、口蓋弓、咽頭壁を支配するものであろう。
顔面中間神経中の体性求心繊維の実際上の影響(結論-consequence)と関しては後述する。
味覚路舌2/3 中間神経
| (f)三叉神経Trigeminal nerve |
知覚繊維は:体性求心性
第1,2,3枝支配域の重複は少ないといわれる(デルマトームの重複と対照さる)
三叉神経は、通常耳介の耳甲介concha (VII, X, 恐らくIX,により支配)には繊維を送らぬ。が外耳道の前壁、鼓膜の前部の皮膚支配、(Cushing, 1904).
しばしば見過ごされるが、鼻腔、口腔、上顎洞、前顎洞の粘膜を支配する。
主として迷走神経によって知覚支配を受けている後頭窩のinfratentorial portionテント幕下部位の硬膜を除いて、硬膜は三叉神経からの硬膜枝の支配を受けている。
知覚核 C2-中脳。橋に入り、その底部を横走し、背内側方向に向かい、主知覚核へ、多くの繊維はそこで2分し上行性と下行性(dichotomize)となり、下行性の繊維のあるものseveralは、大変長く、V神経脊髄路を作る(ハッキリした束)。延髄下端で脊髄のLissauerの背外路をfuseする(くっつく)。
spinal trigeminal tract (tracts), solitary tractのtractの用法はその細胞体がCNS外にある細胞の中枢性突起から成るという点で、CNS内の核を結合する線維束に対して用いられる”tract”という通常の使用と異なっている。この命名は混乱を招くもの也。
下行の途中終止、側枝をnuclei of the spinal trigeminal nerve(脊髄の後角の網様質に連なる。gelatinous substance.)に出す。主知覚核(→後索核)(→触覚)は脊髄路核(後角の最背層と相同?)(→温痛覚の伝達に関連するとこの50年間も一般的に考えられてきたが、このように構成的ではないことが実験の結果判明した。)よりも系統発生的に新しい。
“中脳路核”いくつかの特徴あり、細い柱、主知覚核の吻側端から上丘にかけて、第4脳室上部と中脳水道のやや外側、主として偽単極性ニューロンより成る。半月(or他の)神経節のニューロンに似る。この故に(又他の故に)一般に知覚性と考えられる。中脳根はこの核からの主として下行枝、多くは太い有髄繊維、変性法で追跡してTrigeminal nerveの小部、下顎神経に入り咀嚼筋を支配する。故に固有知覚インパルスを伝えると見倣されてきた。支持される。三叉神経運動核
脳幹最上部にある特殊、内臓遠心性柱核、咀嚼筋(咬筋、側頭筋、外側及び内側翼突筋)←亜核区分在り。
他に鼓膜張筋(ツチ骨柄の上端につく) (cp.アブミ骨筋の支配はVII)
口蓋帆張筋
顎舌骨筋Muscle mylohyoideus
顎二腹筋前腹ventral anterior belly of the digastric muscle. N. mylohyoideus顎舌骨筋神経支配
皮質脊髄繊維により活動化、赤核からも来るようだ(Edwards, 1972)。生理学的(のみ)に、視床下部や扁桃核からjaw reflexへ影響を及ぼす如し、この繊維結合は不明。主たる入力は三叉神経の知覚枝(経由)から来る。しかし、一次性知覚求心繊維は運動核内の細胞には終わらぬ。しかしながら、中脳路核からの繊維はこの運動ニューロンに直接のシナプス結合する(後述)。
知覚性三叉神経核の構成と結合
脊髄路核
caudalis(下核)
interpolaris(中位核) Olszewski (1950)
oralis(上核)
caudalis下核は構造上後角に似る。
marginal zone (peripheral)
gelatinous part (intermediate)
subnucleus magnocellularis (a more massive deep part)
| (g)外転・滑車・動眼神経 |
純粋に遠心性と普通みられている。動眼神経の副交感部分を除き(これは内眼筋intrinsicを支配)。他の核は、体性・遠心性の核柱。
外転神経核:
第4脳室底。VIIに被る。内側にはFlmがある。海綿静脈洞の外側部を通る。上眼窩裂より外直筋支配静脈動内で頸動脈神経叢からの交感神経繊維と合す。
滑車神経核:
下丘レベルで中脳に在る。正中に近く水道やや腹側、すぐ腹側にFlmがある。背側より出る唯一の脳神経、核を出、軸索やや下行し、中脳水道の背側・正中交叉(Fig. 7-22。前随帆のやや前方で外側から脳を出る。それから橋の上縁(端)に沿って大脳脚のまわりに迂回し(7-6図)。小脳テントの前方付着ヒダの所から硬膜を出てくる(7-24図)。静脈洞で外側壁近く位置し、そこを貫いて上眼力裂より眼窩に入り上斜筋を支配する。
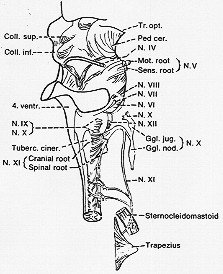
FIG. 7-6 Drawing of the caudal part of the brainstem with the rootlets of cranial nerves IV to XII, illustrating particularly the relations of the accessory nerve (see text).
動眼神経核:
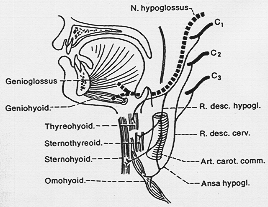
FIG. 7-5 A diagram of the hypoglossal nerve and its connections with spinal nerve fibers, spplying the infrahyoid muscles via the ansa hypoglossi (see text).
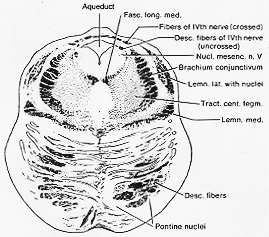
FIG. 7-22 Drawing of a tranverse section throgh the rostral part of the pons in a 20-day-old infant. Weigert's myelin sheath stain. A large proportion of the fibers from the cerebral peduncle and the pontocerebellar fibers are still unmyelinated and therefore not clerarly visible.
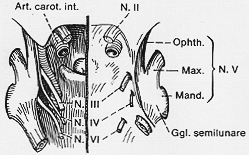
FIG. 7-24 Diagram of dissection of the cavernos sinus region, displaying the course of the IInd to VInd cranial nerves. In the right half of the diagram the dura is retained, in the left it is removed. (Cf. text.)
上丘レベル、複雑、正中近く、水道の腹側に在り、核の外側と腹側にはFlmが位置す(4-5図)。脚間窩をnucleus of the abducent nerve出ず。赤核を一部貫く。神経束の最外部は大脳脚の内側部を貫く。数束が合し、V. III.となり。前方に進み、硬膜を貫き海面静脈洞に入る。上顎窩裂。
上肢-上直筋、上眼瞼挙筋
下肢-下直筋、上斜筋、内直筋。
更に網様体神経節に行く節前繊維(←内臓遠心繊維)(瞳孔括約筋、毛様態筋へ)(後述)。海綿静脈洞の中で動眼神経(III)、滑車神経(IV)に僅かの交感神経繊維が加わる。
小型、節前・自律神経タイプの梨状細胞pyriform cells
外側核と内側核の一部は主として体性運動タイプ(IV, VIと同じく)。
e.g. VIIと同じく亜核区分、各筋の支配在り。
内直筋、下直筋、下斜筋…同側性
上直筋…対側性
上眼瞼距筋…両側性 との同意一致した意見がある。
Ⅲ、Ⅳ、Ⅵすべてがmotor efferent neuronでなく介在ニューロン(Tarlov と Tarlov, 1971; Gacek, 1974)あり。これらは種々の動眼神経(亜)核間の“interneurons”かも?(internuclear neuronと呼ぶべき)。外転神経内の“介在”細胞が対側動眼神経核 内直筋支配(antagonist領に終わる(Bienfang, 1978)へ投射することもARGで証明されている(Graybiel と Hartwieg, 1974; Baker と Highstein, 1975)。HRP法で若干の(III, IV, VI)細胞が小脳、とくに前庭小脳へ(Kotchabhakdi と Walberg, 1977)。動眼神経の遠心繊維は一部同側性で一部交叉性、核の種々の部分(亜核区分)でとも交叉、非交叉の比率に対する知識はヒトで欠如。サルではWarwick (1953)の仕事がある。
眼筋は反射的にも、随意的にも動く。随意性支配は恐らく皮質の眼野cortical eye field(後述)からおこる皮質線維によるものであろう。しかしながら、両眼使用の運動の多くは厳密な意味では随意的とは云えない。すなわち、眼軸を一定点に定める固定fixationの運動は反射的におこる。更に前庭インパルス、頸筋からのインパルス、および聴覚や他の刺激が眼の反射(的)運動を引き起こすことがある。眼球運動は種々様々である。
・あるものは、ゆるやかなslow movements (運動)で(追跡運動pursuit movementsといい、動く対象を目が追っていくとき及び、近くの物体に焦点を合わせる軸輳運動convergence movement)。眼振の緩除“能動active”相もこのタイプ也。
・他の運動は迅速なものfasts(サツケード、様運動で(目)標的の像にに対してfovea(中心窩)をもってくるおこと、及び眼振の迅速相quick phase).サッケードsaccadic movements: 視点の変化と同時に両眼が不随意に急激に、急速に、小運動したり、痙攣的に反射運動する一連の現象。最後に、外眼筋は固(注)視の間、両眼をその位置に保つことが出来なくては成らない。
眼筋、動眼ニューロン、reticular formationからの記憶など数多の仕事が近年なされているが、とくに眼球運動の中枢性制御の機構に関しては未解決の点が多い。専門書としては 註56]. The control of Eye movement (1971), Control of Gaze by Brain Stem neurons(1977) 他。ここでは主としてこの主題に関連ある解剖学データについて主に考察する。個々の外眼筋の動きの機械的特徴は、その眼の光軸optical axisに対する相対的な位置をみれば原則的には理解できる。petiteにその説明あり。外側直筋および内側直筋は視神経の周囲から起こり眼球の外側面および内側面に付着しているので、これらは実際的には各々純外転勤および内転筋として作用する。しかし、上直筋および下直筋に関しては、これらの長軸が休止時の眼の光軸からそれているので問題が複雑になる。眼が約25度外転した時始めてこの両軸は平衡となり、従ってこれらの筋はこの位置においてのみ眼を純粋に上に上方または下方に引くのである。もし眼軸が真直前方に向かっている時は、上直筋と下直筋は副作用として内転作用を表す。上斜筋と下斜筋はその付着腱が内方および前方から眼球に達しているので、内転時限をその横軸の周りに動かし、約50度内転した時に最大の作用を発揮する。この位置では上斜筋は視軸を下げるだけの働きを有するのに対して、下斜筋は純粋な挙上筋である。中間位では上斜筋は下直筋と共働きし、下斜筋は上直筋と共働する。すなわち斜筋はその外転要素によって直筋の内転傾向を補正するのである。故に大抵の眼球運動では(純外転および内転運動を除く)斜筋と直筋は共働きするといえばよい。図7-27には各眼筋の作用を模型的に示してある。
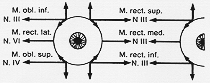
FIG. 7-27 Starling's diagram of the action of the extrinsinc eye muscles.
外眼筋 非常に精密に調節された協同する運動(眼の)
豊富に神経支配
動眼神経 24,000本
滑車神経 3,400本
外転神経 6,600本
筋繊維は細く、運動単位は小、d.h. 各神経細胞はほんの少数の筋繊維を支配する。恐らく6ヶ。 (Whitteridge, 1960)。
最近の仕事によれば
外眼筋の筋繊維は、5つのタイプに分けられる。
1つは”phasic”で速い眼球運動(サッケード、眼振の迅速相)に
3つは”tonic”で注視の維持、正確なゆっくり追う運動、眼振の緩除相に眼筋の筋電図。
眼球運動に作用する神経要素は多いが、2-3の点のみ。
第1に、1つの眼筋の収縮は拮抗筋antagonistの弛緩を伴う(Sherringtonの相互神経支配の法則)。
第2に、眼筋には常に一定度のdifferential tone(応差緊張)があり、この故に両眼の軸を空間の同一点に向けるように維持する。この強制的なfusion注視の傾向は1眼を閉じると一時的に失われる。ほとんどすべての健康人でこのとき軽度の斜視が起こる(heterophoria眼球斜位)。pherein to bear + ia視覚融像刺激を除いたとき、両眼の視軸を平衡に保ってない状態。また斜位を単にphoriaと呼ぶ。これは通常機械的要素のみでは眼球の正しい位置を確保することが充分でない証明である。同じ理由で、1眼の視力がすごく減退してしまい両眼の相応する聴覚印象が1つの像に融合することができないときに斜視が起こりやすい。この眼球斜位heterophoriaはある場合、眼球麻痺の診断を不確実にする要因となることは銘記すべきである。
模式的機械的配列からの説明からでは納得できぬ。機能的要素を春過為不可。
上丘からの繊維が
又上丘からの繊維をうける中心灰白質から)→動眼神経(III)、滑車神経(IV)、滑車神経の付近、樹状突起が拡がる領域へ。
動眼神経(III)、滑車神経(IV)、滑車神経核へ直接に行く求心繊維結合の中で、前庭神経核からの投射が最もよく調べられた。
cp.本章の(d)項。特別のパタン特定の域を支配。直接の前庭・動眼反射弓における第一番目に重要なlinksである。他の直接投射は小脳からtraceされた(第5章をみよ)。又、perihypoglossal核(とくにnucleus prepositus前位核)から動眼(III)核へ(第5章をみよ)網様体ニューロンの側枝が動眼核oculomotor nucleiへ動眼神経 (Golgi法)
実験でも滑車神経へ(Buttner-Ennerver と Henn 1976; Graybiel 1977)生理学的に橋reticular formation→動眼神経核へ単シナプス性(Highstein, Cohen と Matsunami 1974, 他) 。頸部からのインパルスが眼球運動に影響を及ぼすことはよく知られている脊髄→前庭神経核は少量入力あるが、 →動眼神経核への直接投射は知られていない。サルを用いたHRP, 動眼核への仕事についてはSteiger と Buttner-Ennerver (1979)をみよ。外眼筋からの固有知覚性インパルスについて多少詳しく。筋紡錘 動物やヒトに(Cooper と Daniel, 1949 a review)あることを昔から、しかし正確な知識はこの30年間。(review. Bach-y-Rita, 1975).しかし未解決(ほど遠い)。何らかの種の固有知覚受容器はすべての動物に見られるが、筋紡錘が証明されたのはほんのいくつかの動物で、例えばブタ、羊、山羊、ヒト;ネコ、イヌ、ウサギでは証明がされてない。Cooper, Daniel と Whitteridge (1951)や他の人々の生理学的研究によれば眼筋からの固有知覚性インパルスのあるものは動眼神経ocular motor neurons (動眼神経、滑車神経、外転神経と思われるKawamura)を介して脳幹に至ると思われる。
いくつかの研究から(e.g. Batini, Buisserat, Buisserat-Delmas, 1975)このインパルスは大部分ocular motor nerveから眼窩内域には海綿状脈洞にある吻合によって三叉神経の眼神経に入るようだ。ヒトおよび若干の動物で動眼神経内に神経節細胞群が観察されている(Bortolamiら1977)。Golgi腱器官からもあり。脳幹に入りたる後の固有知覚のルートは個々別々なるべし。→中脳路核(V)、Alvarado-Mallartら(1975): HRPをネコ外直筋注入しmesencephalic Vに標識細胞を見た。
Manniら、:外眼筋を伸展し三叉神経に反応を記録。(山羊、ブタ、ネコ)神経節細胞の神経突起はtrigeminal nerve(V), pars oralis (上核)(Manni, Palmieri と Marini, 1972).に主として終わる。上核からこのインパルスは同側性に内側毛帯経由で一部、VPL,VPMへ。対側性に上丘へ、(petit, Abrahams と Rose, 1975)。小脳中部視覚野で(ネコ)外眼筋を伸展させたとき登上と苔状繊維反応が得られる。小脳は外眼筋のコントロールに役割を演じるという観察と一致する。
| 副動眼神経核、視蓋前域、上丘 |
accessory oculomotor nuclei (Carpenter と Peter, 1970/71をみよ)。
これら3核はいくつかの領域から求心繊維をうけるが網膜から繊維は入らない(図7-31参照)。
petit. Flm→Cajal他これら3群への求心路はあまりよく知られていない。
8野→Cajal
中心前回、中心後回→Darkschewitsch
上丘→Cajal, Darkschewitsch(←小脳核、小脳核、前位核(XII))
網膜→some nuclei of視蓋前域
脊髄→Darkschewitsch
Cajal間質核刺激で頭部の回転、眼球の回転(ocular torsion),眼球の偏位(在)(deviation of the eyes)がみられる(rotation of the head)。
間質核(Cajal)から
脊髄(interstitiospinal tract第4章)
前庭神経核(本章の(d)項)
舌下神経周囲核、
paramedian reticular formationの背側部。 Carpenterら(1970), Loewy と Saper (1978) HRP: (Cajal, DarkschewitschでHRP+細胞
更にthe accessory →III核proper.
Darkschewitsch(の孤立した)傷今まで成功していない。後交連核からIII核へ投射するかも不明。
HRP法でGraybiel と Hartwieg (1974)は
間質核→III核を確かめた。
後交連核→III核を示唆した。
後交連のレベルで上丘のすぐ吻側にある。亜核区分(著者により少し異なる)。Kanaseki と Sprague, 1974
網膜繊維が主として対側性にsublentiform核に終わる(7-31図)。(小域の)pretectal olivary核におわる。nuclei of pretectal areaにも終わるという人達あり、retinaの他に、視蓋前核は、大脳皮質や上丘から繊維をうける。pretectal nucleiからIII, IV, VIへは投射せず。求心路は、視蓋領、8野、LGBv., 又恐らく視床のposterior領域、橋reticular formationから遠心繊維系は、或る程度、相互的reciprocal to thin efferents.
上丘、LGBv, accessory oculomotor nuclei, Pelvic、R核(視床)、髄板内 群の一部、reticular formation(脳幹) 視床下部、;Itoh(1977)により大部分記載された。reticular formation, Pons, inferior olive
視蓋前域は瞳孔対光反射弓に関連する領域と考えられてきた。故にEdinger-Westphal核(内臓・運動核)に投射するか否かは興味ある質問である。サルでは毛様体神経節摘除去の逆行性細胞変性に基づき(Warwick, 1954; Pierson と Carpenter, 1974)III核群が内臓性支配を供にするらしいことが決定された。Carpenter と Pierson (1973)によって、olivary pretectal nucleusからこれらの内臓性核群に繊維が追跡されたが、このこれらの細胞群の全域に終わるものではない。Benevento, Rezak と Santos-Anderson (1977)によればサルではsublentiform核とolivary核(この核は網膜繊維をうける視蓋前核也)。から出ず。これらの繊維の若干のものは後交連を交叉し対側の内臓核に至る。これらの近年の所見は対光反射と(両眼)調節accommodation反射の理解に貢献せり、この問題は後で立ち戻る(7-31図をみよ)。亜核間に結合上の差あり。
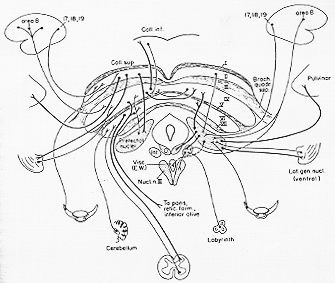
FIG. 7-31 Simplified diagram of the main afferent and efferent connections of the superior colliculs (left side of figre) and of the pretectal nuclei (right side). The various nuclei of the pretectm are not indicated (see text). Efferent connections from a region are shown by heavy lines, its afferent connections by thin lines. Note that the layers of the superior colliculus differ with regard to the origin of their afferents and the destination of efferents as frther described in the text. The connection shown as a broken line from the pretectal nuclei to the visceral (Edingertion shown as a broken line from the pretectal nuclei to the visceral (Edinger Westphal) nucleus is considered to be part of the elementary reflex arc for the light reflex. Many known connections are nort included. To simplify the drawing, all connections except the tectospinal tract are shown as being ipsilateral. For particulars see text. Concerning layers of the sperior colliculus see Fig. 7-29. D: nucles of Darkschewitsch; Int.: interstitial nucles of Cajal.
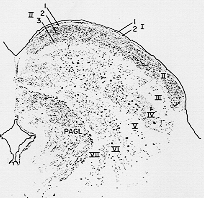
FIG. 7-29 Projection drawing of the superior colliculus in the cat, showing its lamination as seen in a Nissl-stained section. I: Stratum zonale; II: stratum griseum superficiale; III: Stratum opticm; IV: stratum griseum intermedium; V: stratum albm intermedium; VI: stratm griseum profundum; VII: stratum album profndum; PAGL: lateral nucleus of periaqueductal gray. Layers I and II can be further subdivided (1,2,3). From Kanaseki and Sprague (1974).
第3番目に出場の上丘
下等脊椎動物で最も高度に分化した視覚系構造物。”corticalization”しかし問題はそう簡単でない。哺乳類
I層(帯層stratum zonale) I.
II層(灰白層stratum cinereum) II. Stratum griseum superficiale
III層(視神経層stratum opticum) III.
のこりまとめて(IV, V) IV. stratum griseum intermedium
(毛帯層、stratum lemnisci) V. stratum album intermedium
stratum = lamina VI. stratum griseum profundum
- stratum album profundum
(From Kanasaki と Sprague 1974)
上丘求心繊維のうち機能上最も重要なものは恐らく網膜から、LGBへ行く繊維から分かれる。視索の内側部(大部分は視索交叉で交叉をしている)にあるこれらの繊維は、superior quadrigeminal brachium (superior collicular brachium)上丘腕を通り、同側上丘に至るが、若干は(又?)交叉し対側の上丘に至る。これらの繊維は視神経層にはいる。が少量(は)帯層(I層)に入るものがある。下等脊椎動物や他にラット、ウサギ、オポサムなどで厳格に局在ある配列retinofectalをするが、高等哺乳類になると上丘へ行く視神経繊維の数は減り、clear-cutな局在を決定することが難しくなる。最近の仕事でretinotopical投射がネコ、サルにもあることが判った。
網膜の下半(すなわち、視野の上半)…上丘の内側に投射
網膜の上半…上丘の外側に投射
水平経線(horizontal meridian)は前外方から後内方に走る。(7-30図)。視野の中心部は上半の前外方に再現され、周辺域よりも大きい領野を占めるJulia Apter (1949)パイオニアの仕事(電気生理学)Feldon, Feldon と Kruger (1970), Goldberg と Wurtz (1972)たしかめられた。
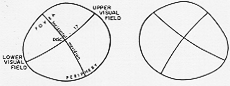
FIG. 7-30 Diagram of the representation of various parts of the visual field in the superior colliculus, as determined in an autoradiographic study in the monkey by Hubel, LeVay, and Wiesel (1975).
(上丘求心繊維)
皮質V層→主にII層にある細胞に終わるしかし網膜繊維よりも深く終わる。(後頭様)同側性(厳密に)他の皮質域からも
脊髄上丘路。
Mehler, Feferman と Nauta (1960) 深層へ、対側性。
下丘上丘路
Moore と Goldberg, 1963, 1966; Powell と Hatton, 1969. 深層視蓋前域
上丘から繊維をうける領域、相互的結合、中間と深層へ。LGB v
主に側性、小量 中間層小脳(室頂核)movements elicitable from the SC.
深層へ、上丘からおこる運動の小脳性コントロールを示唆(5章をみよ)
黒質(網様部)
中心灰白質
zona incerta
視床下部、
reticular thalamic nuclei
最終更新日:2002/09/13
