| 第8章.視覚系 |
要約:慶應義塾大学解剖学教室 川村光毅
・視覚性インパルスの経路一般および視神経の部分交叉
・視覚系のおける局在性
・外側膝状体(LGB)
・視放線
・有線野(Brodmann's area 17)

視覚系の検査は、傷害の部位局位やしばしばその性質について神経学者が価値ある結論を導き出すことができるので、臨床神経学上極めて重要なものである。視覚系路は眼球から大脳半球の後頭極までつづいてひろがっているが故に、脳内のいろいろの場所での傷害で視覚系傷害にもとずく症状がおこりうるし、この種の症状はしばしば起こる。これらの症状を正しく把握する(evaluateする)ために、解剖学的構造の知識が不可欠である。
網膜(視覚受容器)はもともと(大脳)半球のevaginate(裏返しする、外転する、翻転するturn inside out, evert?)した部分である。胎生早期に最初の脳胞と交通しているoptic vesicle眼胞-前脳の側方突出-視溝optic grooveより発達-として発生する。この起源は構造に反映されていて、網膜は主として神経細胞で構成されている。神経細胞層の1つである双極細胞は樹状突起を網膜の色素層の方向に外方に出しており(眼胞の外壁より形成される層)。この樹状突起は、視覚受容器である。桿状体細胞(rod)および錐状体細胞(cone)とシナプス結合する(8-1図)。中枢方向へ行く双極細胞の軸索は神経細胞層(layer of ganglion cells, Nervezellenschicht)の細胞をシナプス結合をしており、これからの長い軸索は視覚性インパルスを外側膝状体にまで伝える。この繊維は最初、内側の網膜、神経繊維層(fibrillar layer)を通り(この部分では無髄神経繊維(藤田ら))視神経乳頭にむかいここで(papilla, = optic disk視神経円板)眼球の壁を貫き集まりて視神経となる。
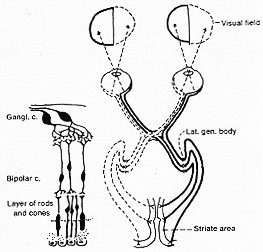
FIG. 8-1 Simplified diagram of the basic opcic pathways, illustrating the partial decussation of the optic fibers in the chiama. Below to the left, a figure indicating the arrangement of the cells of the retina.
cp. Fujitaより、ganglion cellsのaxonは乳頭にむかい(Retinaの神経繊維層では無髄)、ここで胸膜のつづきの薄い膜、すなわち、胸膜篩板lamina cribrosaを貫いて(貫くや否や有髄となる)眼球を出て、視神経となる(繊維の数は80-100万本で1億をこえる網膜の視細胞よりはるかに少ない。シュワン鞘はなく、gliaがある。註1)Arey と Gore (1942)によれば、イヌですべての視神経細胞は軸索を視神経の中に送る如くて介在神経細胞は(ないようだ)。実証できなかった。
視交叉で部分交叉してLGBに至る。この際可成りの繊維が上丘、視蓋前核に終わる。これらの網膜繊維は視覚系(性)の反射弓optic reflex arcsの基礎をつくっており、第7章で扱った。retinogeniculate, retinocollicular, retinopretectal投射の他に、網膜からの繊維は中脳の被蓋にあるいわゆる副視索核nucleus of the accessoryに終わる。早や100年前に副視索のあることはわかっていた(Gudden, 1970, 1981)。が最近新しく関心がよせられた(Marg, 1973の総論)。とくにPasik と Pasik(文献はPasik, Pasik と Hamori, 1973をみよ)の実験により、サルで、副視索系は(accessory optic system)は線条野なしでの欠除の際の基本的(basic)な残された(residual)視覚機能にとって必要なものと思われる。LGBからは、大脳半球口質を通り後頭野、線条野、鳥距溝上下両側へ(視放線形成)。ヒトにありて視覚visionの大切なること疑い少い。次のことこの証在なるべし即ち、すべての求心性と遠心性を含めて脳神経中ヒトの視神経(100万: Kupfer, Chumbley, と Downer, 1967)は実にその38%を計う(BrushとArey, 1942)。
視神経繊維は経過中規則正しく配列し混ぜず。meridians medius middle, +dies day
経線:軸線を通過する面が球体表面を切るときに、球表面上にできる仮想上の線。
右側の線条野striate areaは視野の垂線経線vertical meridianの左側にある物体の知覚に関与する。このことは、右側の大脳半球は、身体の左半の運動と知覚の活動に関連するという事実に相応ずるものである。先に進む前にHubel と Wiesel (1965, p.229)を引用することが適切であろう。即ち、日、「生理学的術語でvision視覚を理解することは生物学者にとってそら空しい問題formidable problemである。それはつまり、神経系が、入ってくるメッセージを、形、色、動き、深さというものが感じられ解かされるように、いかにとりあつかうかということを学び取るに等しいことだ」最近この目標に向かって可成りの進展がみられた。受容器である網膜や視覚に関連するCNSの一部について圧倒されるような構造上、機能上の複雑な仕組みを明らかにした。電顕で、生理学的に、網膜のくわしいこまかい仕事。しかしながら、たとえ網膜がすべての視覚インパルスの出発点であり、それ故にその構成が視覚系の中枢部分におこる過程を決定する上に基本となるものであったとしても、この間(主)題は臨床神経学にとって比較的興味は薄いものである。故に簡単に、視覚系路の解剖学的構成に関するところを第一義的にのべる。
| 視覚系における局在性 |
視覚系における局在配列は視交叉における繊維の半交叉にとどまらず更にその先にまで行く。全視覚系を通して網膜の各々の小部分の点対点の(非常にはっきりしたdefinite)が証明されている。網膜の種々の細部分に達した光刺激は明確に局在性を有する経路を通って線条体に伝えられる。幾分か簡単化して云えば、線条体には、網膜で形成された像の真のコピーである“像”(複写像)が形成されるといってもよい。この故にhenschenは線条野を“皮質網膜”と名付けられた。
Henschen (1890年頃)は視覚系の中の詳細な局在性の存在を主張した最初の人である。多数の人の症例から結論した。彼の最初の論文が、一流の神経学者が、大脳の局在性の考えに対して強く反対していた時に現れたために、彼の見解は激しく挑戦を受けた。次第に特に1920年以降、視覚系の実験的研究がなされてから、彼の意見は正しいと受け入れられる所となった。はじめMarchi法で、retina破壊後→視神経、視索、四半分の占める位置quadrants)
上四半分(網膜)→上方四半分(視神経及び視索)
下四半分(網膜)→下四半分
中心(横斑部)→中心(Brouwer と Zeeman, サル,1926)
視交叉では上部網膜繊維は背側で下半の繊維は腹側で交わる。交叉繊維と非交叉繊維は混じ再配列される。その配列像の様子はLBGにおける相対的な終止部位relative and stationに相応する。従って,視索では,網膜の中心の横斑部の同側半homonymous halvesからの交叉性および非交叉性繊維は、横断切片上の広い領野をを背外側に占め、[網膜下部四半分からの繊維は外側部に]上部四半分からの繊維は内側部を占めることが明らかにされた。8-2図にみられるように、同じパタンが外側膝状体にもみられる。横斑繊維はLBGの中心後部域に広範囲に終わり、網膜周囲部からの繊維はより前方に終わる。更に、上方四半分からの繊維は内側に、下部四半分からの繊維は外側に終わる。横斑部内でも、横斑上部と下部からおこる繊維の間に同様の区別が見い出されている。
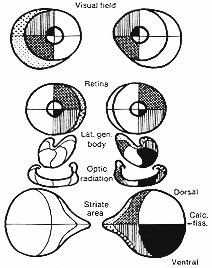
FIG. 8-2 Diagram of the porojection of the different parts of the retina in the lateral geniculate body, the optic radiation, and to the srtiate area. Redwawn from Polak (1932).
8-2図Polyakから。
ニューロン超え細胞変性(サル網膜障害後、LGBで)、Le Gros Clark と Penman (1934)。第1章の図1-17(p.25をみよ)。境界ハッキリ、3層(6層のうち)のみに変化
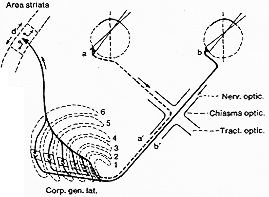
FIG. 8-3 Diagram to explain the anatomical basis for the impulse propagation from corresponding parts of the two retinae to the same region in the striate area. (See text.) Redraawn from Le Gros Clark (1941a).
図8-3. Le Gros Clark (1941a)より
HRP法で実際上(ほとんど)すべての網膜の神経細胞がLBGに投射するのをみた(Bunt et al. 1975)。LGBに網膜外側膝状体繊維が入る際には、交叉性の繊維は、しかしながら、まだ分離されていない。膝状体→皮質線維も同様の正確性をもった局在性あり。皮質小傷数週後LBGに(同側)逆行変化Polyak (1933)。
局在は
網膜上部四半分→鳥距溝の上部(又は/即ち、背側部)
網膜下部四半分→鳥距溝の下部(又は/即ち、腹側部) 再現 (図8-2をみよ)
この原則は網膜の周辺部に中心部・横斑部にも同様にあてはまる。また横斑部の再現域は線条野の後方の広い部分を占めており、この領野の線条体全域に対する割合は、横斑部の網膜全体に対する割合よりも相対的にはるかに大であることが判る。このように(すなわち)横斑は周辺部より線条体に占める領域が広く(more amply represented)この知見は、横斑はvisual acuity視感覚が最も高度で且、感覚細胞が最も密である網膜の部分であるという事実に完全に一致する。横斑域(皮質の)前方には、同名(側)性の網膜の周辺四半域が再現されている。即ち、網膜の上部は鳥距溝の上方に、下部は下方に。線条野の最前域は、網膜の再鼻(内側)側周辺部を代表しており、単眼視視野である最も外側(temporal)の半月部crescentに相当している。以上サル所見だが人も原則的に同じ様だ。線条体障害の際の視野欠損を説明する際実際的に価値がある。
上述の簡単な説明ではLBGは視覚の中継核のように取り扱ったが、実際は、LGBは、背側部(大きい)と腹側部(小さい)の亜核に分けられる。腹側部は細胞構築上更にいくつかのはっきりした亜核群に分かれる(Jordan と Hollander, 1972)。両方(dorsalとventral)とも網膜からの繊維をうけるが(Sanides, 1975)。大脳皮質へ視覚情報を伝えるのは背側核(LGBd)のみであるようだ。ARG法を用いて(Edwards, Rosenquist と Polmer, 1947; Swanson, Cowan と Jones, 1974; Ribak と Peters, 1975)LGBv(腹側核)はいくつかの皮質下核に投射することを示した。それらは
LGBvから
不確体zona incerta
視蓋前域pretectum
上丘superior colliculus
副視索の外側終止核lateral terminal nucleus of accessory optic tract
視交叉上核suprachiasmatic nucleus
網膜からの投射とLGBv域から投射域で上記のいくつかの部位が重複しているらしきことは注目すべきことである(Swanson, Cowan と Jones, 1975)。
LGBvの機能的意義と結合関係についてはなお理解が進んでない。故に以下と外側膝状体に言及されたときは、特にことわらぬ限り背側核のことを意味する。LGBの主な任務は、両側の網膜の対応する部分からのインパルスを融合fusionさせ得るようにすることであろう(図8-3をみよ)。6層のうち3層に各々小域にパッチ状に規則的に柱状にひろがりならぶ。これら6層(柱内の)の細胞は線条体の限局された領域(d. 図8-3)に投射する。ネコ、ウサギ、サルで証明。故に再網膜や相応点からのインパルスは線条体に到してはじめて合することになる。生理学的に証明多数の細胞がstriate cortexで両眼からの刺激に応ずるが(後述)、LGBレベルでは発見することは大変稀也(Erulkar と Fillenz, 1960; Hubel と Wiesel, 1961;也)。
しかしながら、LGBは単なる純粋の中継所に非ざる如し、(近年の研究から)。その皮質への投射も上述の如く簡ならざるべし。
ヒト・サルで細胞からなる6層は繊維により分かれた。LGBの主たる背側部に区別される。腹側から1.2.と番号づけられる。1層と2層は大細胞で他の層はより小さい細胞から成る(Figs. 8-3, 8-4)。LGBサルのものはヒトに似ているが、ネコのものはニッスル標本で、3層のみがはっきりしと区別される。
周辺部では(網膜周辺部-そこに桿状体細胞rodsのみ存在-からの繊維の終止部に相当)若干の層(3と5,4と6という風に)が融合し、4層だけしかないようになり。ここでは、大細胞の小細胞にに対する割合は、中心域より大となっている。
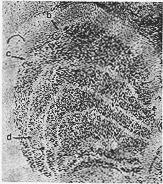
FIG. 8-4 Photograph from a section through the lateral geniculate body of a monkey. In the three small areas, labeled b, c, and d, the cells have disappeared because of three small lesions which were made in the striate area some weeks before death. From Polyak (1933).
(p) Retina大細胞 →LGBの大細胞へ。
LGBの大細胞→18野
LGBのすべてのサイズの細胞→17野
ヒトのLGB-cells数は、100万本で視神経繊維数100万本と1:1の比率だが、このことは直ちに1本の繊維が1つの細胞に結合するということを意味するものではない。結合はaxo-dendritic Golgi研究
LGB細胞reveral types大いさ、形、突き(dendrite axon)の分布状態で樹状突起の分布域は大体その層内にとどまるが、若干のタイプはとなりの層へ行く。Golgi II型の短-軸索細胞も可成りの数みられる。
HRP法
geniculocortical cells5つのタイプ、うち4タイプが長軸索で直接に且局在的に線条体に投射する(Laemle, 1975)。
LGBのシナプス構成、動物種間で原則的に同じ、視神経終末をとりかこむ”synaptic glomerulus” (Szentagothai, 1973)が目立つ特徴で小脳の糸球体と似ている。視神経繊維はGolgi II型にも視床皮質中継細胞にも(樹状突起に)終わる。ここには多数のdendrodendriticシナプスがみられる。
LGBの活動は後頭葉皮質により影響されうる。視覚インパルスの中枢性コントロール、Presynaptic cortical抑制も報告されている。
特定の経路をとり(ヒトで)神経学者にとり重要也。LGBを去ると繊維は短い距離を外方に向かい、やや前方に、内包の最後部の内を通る(いわあゆるpars retrolenticularis )こうして繊維は側脳室の前方に位置を占めるに至る。若干の繊維は、側脳室の側頭角(下角、temporal horn)と中心部(正中室cella media)の移行部の前にあるが、残り、即ち繊維束の下部は、下角の前上端部の前方に位置している(8-5図をみよ)。更に繊維は下角及び後角の外表面に至り。ついに線条体に至する。この部分では、脳室外側壁の直下にあり、いわゆるexternal sagittal stratum中にある(Pfeifer, 1925)。後頭葉と側頭葉の障害における症状の説明にとり興味あることである。
視放線内の繊維の配列にも局在性がみられる。線条体の上部に終わる繊維は視放線上部にあり、LGBの中心部からおこる横斑繊維は視放線の広い中心部を占める。多少の混合ありとも局在の原則は保たれている。ここの傷害は、よく限局された同側(名)性homonymous視野欠損(scotomas,[視野]暗点(視野内にある視覚の低下した部分でやや低下した、または正常な部分に囲まれている。))をおこす。後述する。
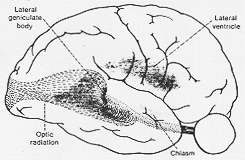
FIG. 8-5 Diagrammatic representation of the course of the fibers of the optic radiation. The geniculocalcarine fibers are seen to swing from the geniculate body (not visible) around the lateral ventricle to reach the striate area. From Sanford and Bair (1939) after Cusing (1922).
ヒトでは殆どすべてが内側面にある(10-3,12-2図をみよ)。下等動物では多少とも外側面(凸面)皮質にあるが、サルでは一定の部位、とくに横斑部はなお外表面にある。新鮮標本においてもlight strip薄い線条がみえる部分故この名あり。このGennariあるいはVicg d’ Azyr線条は有髄繊維の層から成り灰白質の中に白く見える。(之はIV層において外Baillarger線条がとくによく発達したもの-IVa,b,c亜層)髄鞘染色標本では皮質の中に黒い線として容易に認める(8-6図)。これは有線野に限られる。細胞構築上の、特徴があり、内・外の顆粒層がよく発達している。顆粒層、IV層はとくに厚く。IVa,b,cの亜層に分けられる。IVb層がジュンナリー線条に相応し、(一致し)Nissl標本でみると細胞が少ない。III層の錐体層も主として顆粒細胞を含んでおる。V層はハッキリ固定し得ぬ。有線野にあまりにも特徴的な豊富にみられる顆粒細胞と顆粒皮質granular cortexの高度の発育、程度は低いが、他の皮質野、主に知覚機能にあづかる皮質にみられる(12章参照)。17野をかこみ脳の視覚機能に関連する2つのareasありブロードマン18野(area parastriata旁線条野)、17野をすぐ(直接)かこみ境界明瞭、顆粒層が多いが、加え主にIII、Vに錐体細胞が存在。
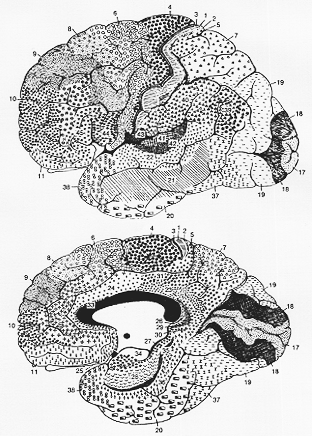
FIG. 12-2 Brodmann's cytoarchitectural map of the human brain. The various areas are labeled with different symbols and their numbers indicated by figures.
(12-2図、791頁をみよ。)
19野(area peristriata,周線条野)、構築18野に似るが広い(18野よりも)意識明瞭なヒトで後頭葉皮質が電気的に刺激を受けると主観的な視覚性sensationが通常引き起こされる(Brindley, 1973をみよ)。それらは刺激をうけた領野によって幾分異なる。単純な視覚性“幻覚”hallucinationが通常みられる。スパーク、フラッシュがみえるという。(幻視、花火、光の明減、片頭痛の際の閃光暗点、異なる視野に(刺激部位と一致して)みえる。網膜周辺部が再現(投影)さえている有線野の全部を刺激すると視野の周辺部から進まり中央部に進む“幻覚”をおこすが、これは片頭痛の際にみられる通常のタイプの視覚性前兆に似ている。(prodroma single→prodromata pl.)
18野およびとくに19野の刺激につづいておこる視感覚visual sensationはFoerster (1929)により行われたが、単純視感覚も稀ではないが、もっと複雑であると報告された。動物、ヒト、いろいろの物体などをみる。Penfield と Jasper (1954)によれば通常起こるのは単純な視感覚である。(かかる視感覚は、患者は通常実在にないものだと知っているので、幻覚と名付けるのは適当でない)。電気刺激によって視覚野(19野?)から眼球運動が誘発される。前章を。
有線領17野は視覚皮質で、ここに視覚印象が伝達される(Henschenの“皮質網膜”)。18野と19野、とくに19野は、視覚印象の解釈、および他の感覚印象との統合(解釈)にとって必要である如し、更に視覚領は皮質性視覚反射に関与しており、これは、関心がむけれている目標物への注視を保とうかとする正常の感覚機能により最も重要なものである。
ヒトの後頭葉傷害の症状は以上の見解と一致し、こまかい動物実験の結果とも軌を一にする(矛盾ない)。とくにすごく立振なのはHubel とWieselの仕事。一連の広範且細心の実験でHubelとWiesenは、光刺激を与えてネコ及びサルの視覚領皮質でニューロンの反応の研究を行った。視覚皮質におけるユニットunit(単一ニューロンの活動)は、細胞の発火に影響を与え得る網膜(又は視野)の領域として規定される(ところの)受容野receptive fieldを基にして特徴づけられている。2つのタイプの細胞(x細胞…伝導速度が比較的小、15-23 m/secで単純型細胞を興奮させるものと、Y細胞…伝導速度が大、30-40 m/secで複雑型細胞に興奮を与える。by Stone et al., 1973)が存在する外側膝状体と対照的に、(この)視覚領(17野)は多数の機能上異なった細胞型をもっている。1つのタイプは“単純型”で他は“超複雑型”とさえいう(ものがある)。
p. 光のしま(模様)stripes of lightとか。明暗野間の(端)縁edges between dark & light fieldなど静止又は動いているものなどいろいろの形の白色光刺激に対して網膜を露出(さらした)のちに、麻酔したサルに微小電極microelectrodeを用いて皮質ユニット(単一ニューロン活動)の反応が記録された。外側膝状体では受容野はほぼ同じ円状circularで、その中心部は細胞を興奮させ(on反応)且周辺部は抑制する(off反応)又はその逆もある。単純(型)の皮質受容野も興奮性と抑制性の区分をもっているが、外側膝状体でのもと対照的に、興奮(性)域と抑制域が平行(並行)して直線的に配列している。したがって(かくしてthus)これらの細胞は光のstripe(しま、スリット)に対して反応し、且つ、各々の細胞にとって、視内野のstripeの特定の方向(位)orientationが最も有効である(Hubel と Wiesel 1962,1968,1974ab)。事実、しばしばほんの少しばかり照(光)明の方向を変えただけで特定の細胞に対して刺激が無効となる。特別に光の線の垂直方向、水平方向、又は中間の方向に反応する細胞が登録された(見い出された)。外側膝状体と有線領のニューロンの受容野の形の差は以下のように説明されよう( Hubel と Wiesel 1962)。すなわち、網膜上に直線に添って配列している同心円状に“on中心”がある受容野を有する多数の細胞がLGNにあり、又、これらの(LGNの)細胞の軸索が特定の異質細胞(sing)に収斂convergeして、皮質細胞を興奮させていると推量(仮定)する。有線領への局在投射に一致して、単純(性)受容野をもつ細胞は網膜の特定域に与えられた刺激に対してのみ反応する。
単純型より大きい受容野
“複雑型”細胞は、受容野(これはreceptive fieldとした方がよい)内の位置に関係なく(そのすべての部分から)、適切(正)な方向をもったstimulus刺激に反応する(スリット図形の傾き(受容野の長軸方向に一致するものに最大に反応する)幅に対して(受容野の数分の1以下の細いスリット)鋭い選択制をもつ-外山)。
これらのユニットのあるものは、暗と明との間のコントラストのついた縁によく反応するが光のstripesに対しては全く反応しない。たくさんの他のvariants異変がみられる(遭遇する)。
これらの複雑(型)ユニット(単一ニューロン活動)は視覚領にあるいくつかの“単純型”細胞からの軸索をうけていると想定されている。かくして、“単純型”細胞は皮質統合integrationのはじめの階段をあらわし、early“複雑型”細胞はあとのlateおくれた段階をあらわしていると思われる。これらの所見は、視覚作用におけるある種の特徴を説明しようと試みるときilluminating啓蒙的である。たとえば複雑型細胞の階層性(ヒエラルキ)は、視野内の位置に関係なく、1つの形がいかに認識されているかを説明することを可能にする。
Hubel と Wiesel (1962, 1968, 1974a,b)により提唱されたこの説明は、unitsの位置に関する彼らにより得られたデータにより更に支持される。受容野の軸の方向が共通である細胞群はマンゼンと散在しているのではなく全皮質に互り垂直な柱を作りまとまってグループをつくっている傾向がある。それは丁度、第一体性知覚皮質におけるmodality-specific(様式特異性の)ユニットがそうであると同じ也(第2章をみよ)。しかしながら、単一の柱の中に“単純型”も“複雑型”のユニットもみられる。
Hubel と Wieselの生理学的所見は有線領の形態と繊維結合で作られる所と支持の一致を見る如し、(読者はSzentagothai(1973b)のreviewをいよ。シナプス結合の詳細な記載あり)。光顕変性法でLGBからの繊維は皮質の(有線領の)第IV層に主として終わる。ARGも認めた。EMでも主にIV層、しかに全層に古典的変性ブトンあり、とくにIII層深部とI層とVI層。樹状突起棘に大多数が終わる。Hubel と Wieselの“単純型”細胞が特にIV層に多いのは興味あり。星状細胞-”simple”でIV層、”complex” “hypercomplex” -錐体細胞IVよりも表層か又は深層に密という(Kelley と Essen, 1974)。機能的(上の)柱の2つのタイプが有線領に同定された。すなわち、
”ocular dominance” column [ODC]
“orientation” column [OC]
IV層の細胞は単眼性(monocular)で同側か対側の眼に与えられた光にのみ反応する。しかしながら、IV層より表層又は深層に在る細胞は、両眼性に反応はするが、その(狭い)垂直柱でIV層の細胞が発火をうけた(ドライブされた)側の眼からの光によく反応する。ODCの幅は250-500μm。左右の一対の隣接するocular dominance柱の全体の幅は、180°方向一周するOC[orientation column]の幅とたいへんよく似ている-10°orそれ以下の差で約50μmの幅、即ち180°回転が約1mm幅-Hubel と Wiesel(1977)。-
OD柱とOrientation柱の次元dimensionは、サルとネコの視覚皮質領のintrinsic fiber結合に関する細胞に亙る一連の実験研究(Fisken, Garey と Powell, 1975)に支持を見い出す如し。彼らの報告はHubel と Wieselの生理学的知見-とくに“複雑型”細胞の局在に関する-と有線領内部(intrinsic)の形態学的構成の現在の知識との間(にみられよう)可能なる関連を全般的に批判的に評価して考察さるべきである。18野、19野はもっと複雑な様式の視覚性認知にとって重要である。網膜を光刺激にすると有線領以外の所、しばしばV-II、V-III(Hubel と Wiesel, 1965)といわれる所で活動電位が記録できる。Otsuka と Hassler (1962), Sanides と Hoffman (1969)と領域的一致(ネコ)。霊長類と類同(相同)。第3の有線領外皮質域はネコでClare-Bishop area (Clare-Bishop, 1954; Hubel と Wiesel, 1969b)。ここは霊長類では上側頭回における視覚野に相当するらしい(Zeki, 1974, 文献をみよ)。
網膜→LGBv
→上丘、視蓋前域
→両側性にPulvinarへ(Berman と Jones, 1977,ネコ、ARG)。
LGBvは視覚領皮質には投射しないが、いくつかの視覚に関連した皮質下核、たとえば上丘や視蓋前域に投射する。これらの部位からLPやpulvinarへ投射があり(7章をみよ)。これらから次の経由でperistriate cortical areasへ視覚性インパルスを送るなるべし。(文献略)。
更に17野→18,19
→上側頭回(Martinez-Millan と Hollander, 1975, ARG)?
17野間の交連繊維はない様だがサルで18野に境する17野の部位で交連繊維あり(Zeki, 1971)。
外側膝状体は17野のみならず18,19野にも投射する。Hollander と Vanegas (1977)ネコでLGBで大細胞のみ→18野。すべての大きさ型の細胞→17野。更に大細胞は2分して17,18へという形態学証明がなされた。Garey と Powell 1967; Vanegas, Hollander と Distel (1978); Hollander と Vanegas (1977)。生理学的にも恐らく大きい早い伝達速度のrelay cells (いわゆるY-細胞)→17と18に分岐をもって。小さい遅い伝達速度のrelay cells(いわゆるX-細胞)→17のみへ。Stone と Hoffman(1971), Hoffman, Stone と Sherman (1972), Stone と Dreher (1973)。
視皮質とくに17野から皮質下への投射
強い投射がLGBと他の皮質か視覚中継センター(核)たとえば、上丘、視蓋前域、いくつかの視床核にある(文献多、省略)。Hollander (1974b)リスザルARG
17野→LGBの全層。いくつかの視床核、とくに、PO (posterior nucleus), inferior pulvinar, lateral pulvinar R (reticular nucleus of thalamus) pregeniculate nucleus retinotopic organizationを示す。
小~中型錐体細胞VI層(17野)→LGB
すべてのサイズVB層→上丘、inferior pulvinar,サル17→18,19連合繊維、IV層より浅い層から起こる。
最終更新日:2002/09/13
