| 第9章.聴覚系 |
要約:慶應義塾大学解剖学教室 川村光毅
・聴覚受容装置の支配
・蝸牛神経について
・(聴覚)上行路一般
・蝸牛核
・上オリーブ核複合体と外側毛帯核
・下丘
・内側膝状体
・聴覚皮質
・下行性聴覚路

この章の主題は聴覚受容器、蝸牛神経、および中枢における聴覚伝導路である。コルチ器官とともに蝸牛管は簡単に述べられる。
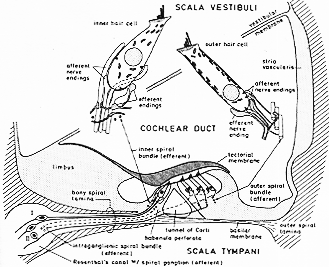
FIG. 9-1 Diagrammatic cross section through the cochlear duct with the organ of Corti and closeup views of an inner and an outer hair cell. Afferent nerve fibers are indicated by fully drawn lines and by dots where transected; terminal swellings are undotted. The efferent nerve fibers are stippled and shown as open circles when transcted. Their terminal swellings are finely dotted. The intraganglionic and inner spiral bundles are composed of efferent fibers; the outer spiral bundle of afferent fibers. Note the inverse relationship of afferent and efferent nerve terminals on inner and outer hair cells. See text. Redrawn and simplified after Smith (1975).
聴覚受容器は、蝸牛管内の有毛細胞である。ヒトの蝸牛管ductus cochlearisは全長35mm、らせん状2 3/4回転、骨迷路(骨性の蝸牛部)の中にある。蝸牛管の基底回転の基端の知覚で球形嚢saculusと交通している。蝸牛管は横断面で3角形をしている(9-1図)。(外側壁は)広く蝸牛管の骨性の壁に密着している。そして
基底板lamina basilarisは(藤田、藤田、組、各論よりp.386)
鼓室唇とらせん靱帯の基底稜の間に張る板で、無構造の基質の中に多数の繊維が平行に走っている。この繊維は膠原繊維とも弾性繊維とも性質を異にする弾性に富む強靱な*フィラメント(径30-100A)であり。基底(basal)回転では短く(65-150μ位)→(振動数基底板の長さに関係し)振動数…多い…高音と共鳴
螺旋状に向かうにつれて長くなり蝸牛頂(apical)では300-500μ→(基底板は弾力性を有し、外リンパの振動によりよく振動する。(らせん器はこの基底板状にのっていることにN.B))振動数…少ない…低音と共鳴
*集束(径1-2μ)して走る。これは聴弦auditory string又は基底繊維basilar fibersとよばる。
前庭感覚装置同様、コルチ器官は2種類の形態的に異なるタイプの感覚細胞を持っている(9-1図)。
内有毛細胞:ビン状battle shaped核が中心部、小器官が細胞質内に散在性分布。
外有毛細胞:より特殊化、核は基底部、核上部の細胞質内にはミトコンドリアをもった平滑ERのsublemmal cisternal系がある。共に、基底部で神経繊維とシナプス結合をしている。有毛細胞の自由路には感覚毛、(不動毛stereocilia)あり、cp.聴毛hairlets Horhaare, 一種の微絨毛microvilliである外有毛細胞では、不動毛が3列又はそれ以上の列を作り(W型を作って)配列している(9-2図)。内有毛細胞では不動毛は殆んど直線の列で蝸牛管に対し平行に配列している(2-4列Fngitasによる)(9-2A)。前庭器官における情況と対照的に(成人で線毛-運動毛が1本で多くの不動毛が並んでいる)。運動毛が存在するのは、胎生期とか非常に初期の生後期間にのみである。
ヒトでは約3,500ヶの内有毛細胞、15,000ヶの外有毛細胞(Guild, 1932; Bredberg, 1968)であり。内有毛細胞は1例に、外有毛細胞は3列平行に配列している(7-2A図)。支持細胞、コルチのトンネル(内トンネル)により分けられる。又、この内を外有毛細胞に向かって神経繊維が横切る。
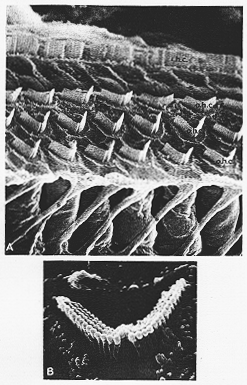
FIG. 9-2 Scanning electron micrographs of the organ of Corti of the guniea pig. A: The single row of inner hair cells (i.h.c.) with sensory hairs arranged in a straifght line and the three rows of outer hair cells (o.h.c.) with a W pattern of sensory hairs. Abbreviation; S: supporting cells. B: The surface of one outer hair cell with four parallel rows of sensory hairs in a typical W pattern. Coutesy of H. Engstrom and B. Engstrom.
聴覚メカニズム
感覚細胞の不動毛がゼリー様(状)の物質から成る蓋膜(らせん上皮の分泌物から出る)と接している如し。蓋膜は、limbusらせん板縁を経て、骨らせん板によって支えられている。聴覚刺激により起こる基底板の振動によって、振動が(おこって)ない蓋膜はおそらくぴんと張ったstiff(固着した)不動毛を傾斜させることになり、それによって有毛細胞は刺激をうける。いかにして基底膜が振動するか、またこの運動がいかにして神経繊維内に電位(potentials)を生ぜしめるかは、まだ完全に理解されていない問題である。最終的には、蝸牛を刺激して細胞内記録してその結果をみるべき。ある期間(for sometime)音の周波数の空間的再現、いわゆる音局在の構成tonotopical organizationがコルチ器(らせん器、organum spirale or Cortis organ)の中にあると知られてきた。蝸牛神経繊維は、その特徴ある周波数により定められている音の周波数(振動数)に対して相当する特殊性を示す。これらの現象は中枢性聴覚経路の構成にとって関連があるので簡単に考察することにする。もともと、周波数discrimination識別に関して2つの相対する説が打ち出された。その1つはHelmholtz (1863)のplace theory空間理論で、他はRutherford (1886)のfrequency theory周波数理論である。この空間理論は、振動数の機能としての蝸牛管に沿って活動の空間的分布を想定するものである。他方、周波数理論は、神経衝撃impulsesの周波数により(る)、刺激周波数stimulus frequencyの直接的再現を想定するものである。最近の研究結果、一定の制約をもって、両方の説(理論)が実際無効であることが示され、且つ、第三の理論としてVolley theory連発(or斉謝、いっせい射導、バレー)理論が2つの折衷としてcombination提察された(Wever, 1964)。
ヘルムホルツの空間理論に従えば、コルチ器の基底端(回転)basal endは高い周波数音により刺激され、頂(尖端、apical)は低い周波数により刺激を受ける。聴覚刺激を与えたとき。鼓膜、耳小骨、内耳液の振動を通じて基底に伝えられてそこに伝達波がset upすえられる(Bekesy, 1960)。基底板上の最大襲撃excursionの点は、刺激音の周波数が減ずるにしたがい(程)回転尖端(蝸牛頂)に漸次移動する。鈍音により近似閾値刺激を与えると、活動電位は恐らく、基底板の最大振動部分上に位置している感覚細胞によってだけ生じる。この仮定はは蝸牛より中枢部にある神経中の求心繊維から記録により支持される(King, 1965whitfied, 1967, 1967)。コルチ器内の空間的周波数再現は、有毛細胞の局在的傷害により起こる特定周波数への聴覚喪失(Held と Kleinknecht, 1927; Schuknecht, 1960)や動物における過剰音刺激時にみられる局所的有毛細胞消失の出現(Bredberg, Ades と Engstrom, 1972)-によっても支持される。患者、死後蝸牛の研究ヒトで特定周波数聴覚障害例でコルチ器の局在的障害、相応する求心繊維の障害もみられるヒト、16~2万サイクル/秒
単一繊維が可成りの広い周波数帯に対応する(蝸牛繊維を強音刺激して記録したとき)。しかし、近似閾値刺激near-threshold stimulation時は、反応帯は非常に狭い。
空間理論は受容器および中枢の聴覚路における音周波数の空間的再現を説明するが、一方、周波数理論は、低周波数の聴覚の理解にとって重要なものである。低い周波数の音で刺激すると、活動化した蝸牛神経繊維の発火は刺激音の相に対して固定される(be locked to)。この現象は、両耳性位相或いは時間的差の探知(発見)(検出)にとって、それ故に後で説明する空間における音の局在にとって重要である。
蝸牛は3セットの神経により供されている。
VII(中間神経も)VIII vestと一緒に内耳道に入り(この内では顔面神経が上部に、蝸牛神経は下部に、前庭神経は中間部位置する)。内耳道底→らせん配列(Truclus spinalis for aminosus, らせん状に配列した小孔)→双極性神経細胞集団一列に並ぶ(らせん神経節、これは蝸牛軸らせん管canalis spirals modioli中にある
(Rosenthal’s canal)といい骨性のらせん層bony spiral laminaのbaseにある。
蝸牛神経は、双極性らせん神経節細胞の中枢突起又は軸索から成る。故に定義に従えば、末梢神経ではなくむしろ知覚神経根sensory nerve rootである。この軸索は厳格な蝸牛局在パタンを構成している(Lorente de No, 1933a; Sando 1965; Arnesen と Osen, 1978)。末梢突起は骨性らせん板を通りコルチ器に向かい真直に放射状分布をしている。骨部を離れるとき、habenula perforataで(9-1図)。無髄となる。(特殊・体性・求心性のカテゴリに、前庭神経同様、属する。
求心繊維の大多数が内有毛細胞のうち比較的小さい集団を支配するが、他方、少数の繊維がはるかに多くの数の外有毛細胞。蝸牛神経中有髄繊維数
約32,000本(ヒト)
約50,000本(ネコ)
90-95%が内有毛細胞を支配、5-10%が内有毛細胞を支配 20本繊維が1つの内有毛細胞に収斂。いくつかの外有毛細胞にシナプス(分枝する)。
この他に遠心繊維efferent fibersが上オリーブ核より、olivocochlear束(Rasmusen, 1946, 1953,ネコ、オポサムで最初に記載された)。ヒトにもある。
この神経束は恐らくコリン作動性cholinergic (Guth, Norris と Robbinの総説を)。起始細胞はアセチルコリンエステラーゼ陽性の大型および小型の細胞(Osen と Roth, 1969)大型細胞は、periolivaryオリーブ近傍核(後述をみよ)にある。その軸索は恐らく有髄性で第4脳室底で交叉する(9-5図)。これらは前庭神経根に入って脳幹を去るが、前庭蝸牛吻合(内耳道内で)を通じて蝸牛神経中に入る。この繊維は外有毛細胞へ行き大きいaxosomaticシナプスを作る。EMで2つのタイプ有小胞と無小胞の神経終末。がみられるが、有小胞vesiculated終末は、一方、遠心繊維に無小胞nonvesiculated終末は求心系に属する。他方に小型の細胞は外側上オリーブの縁marginにある(ネコ)。軸索は細く、恐らく非交叉性で無髄(後出)で、同側の前庭神経根中の(で)交叉したオリーブ蝸牛繊維に加えられる(Rosmussen, 1960)。コルチ器内分布は不明だが、恐らく、内らせん束中(a)、内有毛細胞の下でらせん状をしている遠心繊維の内に束ずるものであろう。これらは内有毛細胞からの求心繊維とen passageシナプスを作っている。
オリーブ蝸牛系の役割は不明だが、交叉性のolivocochlear束を電気刺激すると音刺激により起こった聴神経活動を抑制reduceする。この細胞体は恐らく、上行性と下行性の聴覚繊維(後をみよ)より共に影響をうける。不必要(いらぬ)聴覚シグナルを抑えるに役立つフィードバック機構におそらく関連する。又、内耳の周波数解析力を鋭くする(先鋭化)の役割も討論されている。論議されたところであるが、オリーブ蝸牛束は内臓神経系の一部とは見倣されない直接遠心路であることほとんど疑い得ない。これらの繊維の他に、しかしながら、蝸牛は交感神経幹の神経節からノルアドレナリン(作動)性繊維をうける。2つの異なる機能-即ち、vessel-dependent と vessel-independent系-をもった2系あり。機能的意義は不明、交感神経系のアンバランスによる聴覚障害の可能性は否定できない。
最初の上行系に関する総説はHarrison と Howe (1974a)をみよ。受容器から皮質までの経路中、中断される場所としては、蝸牛核、上オリーブ核、下丘、内側膝状体がとくにある。これらは中断核としてのみ奉仕しているのでない。各々が、聴覚上方の中枢への伝達に対しての影響において、又、他感覚種の情報との相互作用をmediate仲介することにおいて、特別な役割をもっているようにみえる。各中継核は高次レベルからの下行性影響下にある。更に、聴覚刺激によって引き起こされる種々の反射にとってこれらの核は重要である。
求心性蝸牛神経繊維→上行枝と下行枝に分かれれて蝸牛複合体(核)に入る。音周波数の空間的複現の基本である厳密な蝸牛局在パタンがこの核にも存在している(strict cochleotopic pattern)。一次求心繊維は側枝を出してこの核の特定の域にある種々のカテゴリーの第二次ニューロンを支配している。次いでこれらのこれらのニューロンは異なる投射をする。下小脳脚の腹側を通る台形体を経るか。下小脳脚の上(背側)を曲がる中間又は背側聴条intermediate or dorsal acoustic striae(聴条又は髄条)を経る。このようにして、蝸牛神経の単チャンネル系は脳幹を通じて種々の投射を持つ多チャンネル系に変換される。
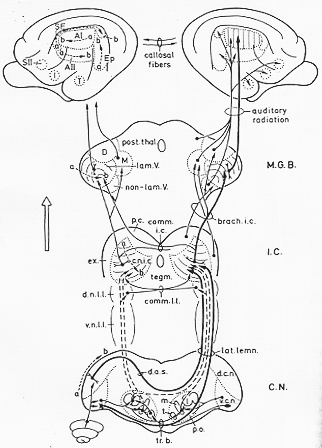
(図9-4)
聴覚上行路
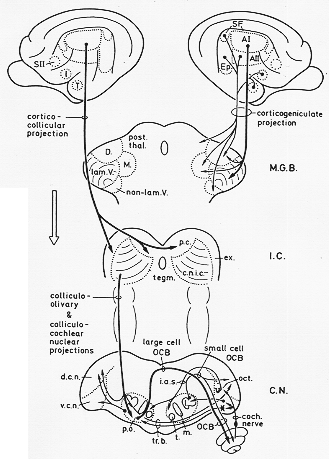
FIG. 9-5 Diagram of the descending auditory pathway of the cat. See Text. abbreviations see Fig. 9-4.
(図9-5、あとの頁に)
聴覚下行路
これらの中枢の経路を通る間に、蝸牛神経核に起こったいくつかのチャンネルはすべて対側の外側毛帯に向かって集中し、ここから下丘中心核に行く。
チャンネルの若干のものは(蝸牛核中の錐体細胞と多極性細胞からの軸索から成るもの)直接上行して下丘に至るが、他のもの(球型細胞、小球状細胞、globular、タコ型細胞octopus cellから起こるもの)は、上オリーブ複合核の種々の亜核でシナプスを作り経路が中断する。
これらの核中で、一側耳からの信号が他耳からの信号とinteract互いに作用し合いとりわけ、空間における音の局在に対して重要な点となる(後をみよ)。上行路経過中、大多数の軸索は恐らく側枝を主たる標的以外の他の聴覚核に出している。多種の異なる経路を進んだ信号は、究極的に、下丘の中心核に到達するが、ここは多くの点で、上行聴覚路の鍵となるオプションである。
下丘から下丘腕を通り前方へ内側膝状体。聴放線、auditory radiationを経て聴覚皮質(側頭葉中の)へインパルスが最終的に伝える(9-4図)。下丘およびそれより上位で、上行性聴覚経路は中核となる投射a core projectionと周辺帯投射a belt projectionに分けられよう。中核となる投射の最終標的は聴覚性の顆粒皮質Koniocortex (9-4図のAI)である。顆粒皮質は、直接の知覚系が終れる皮質のタイプである(Chapter 12参照)。他方周辺帯投射の標的は、第一にこの顆粒皮質のまわりの(とりまく)聴覚皮質域である。これらの領域はRose (1949)によって、-まとめに聴覚性皮質周辺域auditory cortical belt areasとよばれた。周辺帯投射のことばから来ている。Casseday, Diamond と Harting (1976)によって指摘されたように、中核および周辺帯領域への上行性聴覚路の区別はすべての哺乳類に特徴的なる如くで、かくして比較解剖研究上必要と考えられている。ある面では単純化しすぎかもしれないが、中核および周辺帯構造物という風に適(合)した場合には、その区別をこの説明なのかで強調されよう。
中核投射:下丘の中心核、MGBの腹側部の層的構造部、一次聴覚皮質(経路は9-4図で太字で示されている)。第2者の核は解剖・生理学的方法により、空間的周波数複現の基盤が確立したことをみる層的構造が特徴となっている。この特徴は皮質レベルでも保存されている。
周波帯投射:下丘の中心周囲域と背側中脳被蓋で共にMGBの非層的構造部に投射する(9-4図で細字)。これらから聴覚の皮質周辺帯に主として投射する。中核及び周辺帯投射は、後で述べられるように繊維により相互結合がある。専ら聴覚性である中核投射と対照的に、周辺帯投射は視覚および体性知覚somestheticの経路からも入力をうける。これらの入力とりわけ、言語speechのように(な)運動機能における種々の感覚様態の相互作用interactionおそらく(もしかしたら)重要であろう。この相互作用の興味ある面としては、聾小児での言語訓練で、聴覚以外の感覚刺激が有益且つ恵みある(恵益?)らしいという点であろう。
下行性聴覚路の特徴は、ほとんどすべてのレベルで交連繊維がよく発達していること也。台形体や聴条(intermediate and dorsal acoustic striae)の交叉の他に、聴覚繊維は、延髄および橋網様体で正中を横切り、それは、外側毛帯の交連、下丘の交連、それに脳梁である。であるから、皮質下レベルの一側性障害後に(も)、両大脳半球の聴覚領は聴覚刺激により活動化される。しかしながら、各々の耳の対側性皮質複現は優位であるように思われる。それは、おそらく、対側下丘への第二次ニューロンのより豊富な、同側下丘へよりも、投射によるのであおる。
チンパンジー変性繊維実験研究(Strominger, Nelson と Dougherty, 1977)によれば、霊長類では第二次ニューロンで同側下丘到るものの方がより多いし(それ以上に)更に下丘を通過して直接内側膝状体に終わる上行繊維の成分が多くなるようだ。
哺乳類では、第4脳室の外側の奥底(recess, すみ、壁のくぼみ)にある下小脳脚(索状体)の外側に。ヒト脳幹(7-2図. P.451)では蝸牛核複合体(群)は外側にはなれているので、通常の死後の研究では小脳と一緒に切り離される。腹側核は前腹側核と幸福側核とに入っていくる蝸牛神経恨により分けられる。背側核は後者(=後腹側核)の背外側核に位置する。蝸牛神経繊維は腹側核の内で分岐して上行枝と下行枝となる。これは一次性求心繊維一般と同様である。これらの枝は規則性をもった蝸牛局在性(cochleotopic)の順序に配列している。又これがこの系の音局在(tonotopic)構域の基盤となっている。
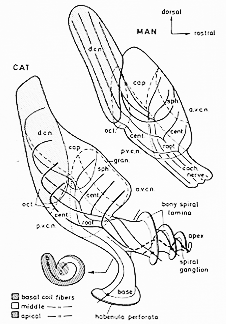
FIG. 9-6 Diagram of the cochlear nuclei in man and in the cat, in lateral views. In the cat the cochlear nerve fibers can be followed centrally from their perikarya in the spiral ganglion through their bifurcation in th ecochlear nerve root and to the sites of termination of their branches. The ascending branches terminate in the area of spheriacal cells (sph.), the descending branches in the dorsal cochlear nucleus (d.c.n.). Note the gradual increase in thickness of the fibers from the basal to the apical coil and the spatial representation of the cochlea in the nerve and the nuclei. In the cross section of the cat cochlear nerve, to the lower left, the cochelotopic organization is indicated by different symbols (key below) and by an arrow pointing from low- (a; apical) to high- (b: basal) frequency regions. In man the distribution of fuibers appears to be the same as in the cat, except for the orientation of the descending branches. On entering the dorsal nucleus, these fibers take a straight corse instead of making a 180° turn, as they do in the cat. The relative size of the various subdivisions of the ventral nucleus differs in the two species. The areas of octopus cells (oct.) and the peripheral cap of small cells (cap) are well developed. A granule cell layer (gran.) is present only in the cat. Abbreviations: a.v.e.n.: anteroventral cochlear nucleus; p.v.e.n.: posteroventral cochlear nucleus. CAT: from Arnesen and Osen (1978). MAN: from Moore and Osen (1979).
これら3つの亜核すべてにコルチの蝸牛底(basal)(高振動数-繊維)→背側 蝸牛頂(apical)(低振動数-繊維)→腹側 (左図9-6、ネコ)。
9-7図の振動数(周波数)を記録した蝸牛核マップと一致。(Rose, Galambos と Hughes, 1959)。しかしヒトの背側核では周波数の軸が異なる。これは、下行枝の特殊な経過からも示唆される(9-6図、ヒトをみよ)。数々の細胞構築学的研究から、蝸牛核は多少とも混在するいくつかのニューロン群に分類されるもの(それら)から成っている。介在ニューロンか、種々の(前述)二次聴覚チャンネルを与えている主たるニューロンで鍵となるものである。伝統的な亜核(分類)から細胞のタイプ分けた注目されるようになってはじめて脳幹聴覚中枢の解明がずっと前進した。
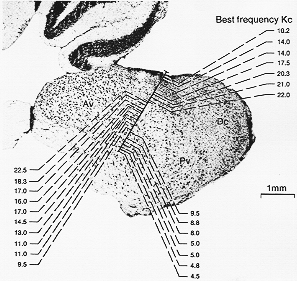
FIG. 9-7 Photomicrograph of a sagittal section through the cochlear nuclei in the cat, showing an example of the spatial representation of the cochlea as determined by mapping of characteristic (best) frequencies. See text. Av, Pv, and Dc: anteroventral, posteroventral, and dorsal cochlear nucleus, respectively. From rose, Galambos, and Hughes (1959).
ニッスル標本で、Osen (1969)により9つのタイプの神経枝細胞、これらの大部分は、電気生理学的特性や求心・遠心の繊維結合からも又区別しうる也。(後をみよ)。多少修正するとしてニッスル像はゴルジー像とも軌を一にする(矛盾しないcompatible with)。Osen(1969)によって記述されたように、この核複合体内にある特定の細胞タイプの分布域は上に言及した3つの亜核の境界とはすべての場所で相応するとは云えない。われわれの知識ネコに負う所だ。故に9-6, 9-7, 9-8図にふれてネコについてのべる。ネコの腹側蝸牛核は細胞学的根拠から3部に分けられよう(9-6図)。すなわち、
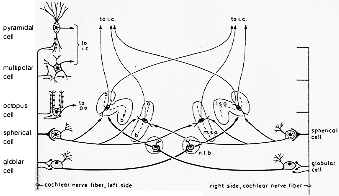
FIG. 9-8 A simplified wiring diagram of the cochlear nuclei and the superior olivary complex, on the basis of observation in the cat. The cochlear nuclear neurons are supplied from the cochlear nerve by different types of synaptic endings. Each cell type has its specific projection onto higher auditory brainstem centers. As shown in the diagram, where only main targets are indicated, the spherical and globular cell axons are responsible for the binaural input to the medial superior olive (m.s.o.) and the lateral supeiro olive (l.s.o.), the latter nucleus receiving its contrralateral input via the nucleus of the trapezoid body (n.t.b.). Abbreviations: i.c.: inferior colliculus; p.o.: periolivary nuclei.
固有の腹側核の背外側は小形細胞の帽子cap of small cells (cap)である。ネコでは、帽子をかぶった腹側核は、もう一つ、顆粒細胞層(granular layer)でカバーされている。霊長類以下の哺乳類(subprimate mammals)では、背側核は層的に構成stratifiedされている(ここでは例示されてない)。(一層)表層の分子層と4つの深部の細胞層に細胞 構築上区別される(Lorente de No, 1933b)。主たる投射ニューロン、すなわち双極性錐体細胞、は放射線状に位置を位置を定められておりoriented、これは、小脳皮質でのプルキンエ細胞の位置に似た位置を側蝸牛核内でとっている。
非霊長類の背側蝸牛核の錐体細胞は、顆粒細胞を介して、小脳皮質のそれといくつかの点で共通性をもっている神経回路により影響をうけていることから、第二次知覚性ニューロンのうちでユニークな細胞である(Mugnaini, Osen, Dahl, Friedrich と Korte, 1980)。この蝸牛核中の顆粒細胞の樹状突起は、高次の聴覚センター(核)又は恐らくは蝸牛複合体内から起こる苔状繊維終末に集る糸球体シナプス配列glomerular synaptic arraysに関与している。顆粒細胞の軸索は、錐体細胞の尖端樹状突起と通過中シナプスsynapses en passageを作りながら、背側核の分子層内で平行繊維を形成している(Mugnaini, Warr, と Osen, 1980)。錐体細胞は、その基底樹状突起の上に、一次求心性繊維をとりわけ下丘からの下行繊維をうけている。背側蝸牛神経核は聴覚系において恐らく、ある点で(は)、前庭系における小脳の小節nodulusのそれに似た役割を演じている可能性がある。しかしながら、現今のところ、聴覚auditionにおけるこの興味シンシンの核の機能機能的意義について何も判ってない。
ネコとラットの所見をもとに、蝸牛複合体の主な細胞型の簡単化した結合図を9-8図に示した。蝸牛神経繊維は、蝸牛核細胞の種々のタイプのものと種々のタイプのシナプス結合をする(文献あり)。
蝸牛核中の細胞型は中枢性投射においても(それぞれ)異なっている。
spherical cells
globular cells
octopus cells 主たる標的は上オリーブ核群(Harrison と Irving, 1964, 1966a,b)(Warr 1966, 1969, 他に)
pyramidal cells
multipolar cells 対側下丘へ(Osen, 1972; Adams, 1977, 1979, 他の)
ヒト
ヒト蝸牛核群のサイズは大約ネコのものと同じしかし、背側核/腹側核の比がヒトでやっと大となる。ヒトでは球型細胞領域がやや狭いことによるものならん。ヒトで上オリーブ核が小さいのとも一致する。ヒトの背側核では小細胞帽は比較的に大、しかし、顆粒細胞層を欠く。ヒトの背側核ではネコと異なり、下行蝸牛枝のorientationが異なる。層的構成もみられない。錐体細胞はその典型的なorientationを失い、分子層の痕跡が残るのみ。であるからヒト では背側蝸牛核は小脳皮質と構造上類同とは考えられない。この差異は霊長類で次第に顆粒細胞が消失していった結果なるべし(Moore, 1980)。恐らく聴覚情報の過程-進行が、大幅に高次センターにとって変わった為、ヒトで下丘に直接投射する多極性ニューロンや小幅細胞small cap cellsのようなニューロンが腹側核で優勢なることは、同じ傾向の一部であろう(Adams, 1977, 1979)。
上オリーブ核は台形体中、ヒトで橋下部に右り、橋核のすぐ背部、動物により可成り異なる。ネコでは境界が明瞭で3部
内側上オリーブ核 medial superior olive (m.s.o.)
外側上オリーブ核 lateral superior olive (l.s.o.)
台形体核 nucleus of the trapezoid body (n.t.b.)
このまわりに散在性細胞域ありてこれをperiolivary nuclei上オリーブ周囲核(p.o.)外側上オリーブ核と台形体核とは相互に関連在り、共にヒト脳で発達が悪い(Moore と Moore, 1971)しかし、内側上オリーブ核はヒトとネコで同大きさ。
脳幹横断面上で、内側上オリーブ核には、横断面上に方向を有する双極性細胞が矢状方向(縦方向)に並んでいる。各々のニューロンは、蝸牛核中の球型細胞spherical cellsの軸索により両側性に影響を受ける。双極細胞の外側にのびた樹状突起は同側からの求心繊維と、内側に伸びた樹状突起は対側からの繊維とシナプス結合をしている(Stotler, 1953;他)。内側核は主として低周波数音に反応し、その細胞(ニューロン)は両耳間の時間や位相差異に明らかに敏感である(Goldberg と Brown, 1968; Moushegian, Rupert, と Langfor, 1967)。内側核は、恐らく、音、とくに低周波(低い振動数の意味、以下同じ)の空間的局在に関連している。同核は同側性に下丘中心核に投射する(Stotler, 1953; Elverland, 1978Adams, 1979, 9-4図)。
台形体核は、上オリーブ核部の腹内側にある。その主体となるニューロンは、蝸牛核内のglobular小球型細胞に似る。9-8図に例示された如く、台形体核は対側の小球型細胞によって太い軸索で支配されている。終止像は(ブトンの)大杯状でin large calyces1対1の対応関係がある。この台形体の細胞は同側の外側上オリーブ核に投射する。
外側上オリーブ核 ネコでS字形に双極細胞の列を作っている。内側核同様両耳から影響をうける。同側性入力は球型細胞spherical cellsから直接くる。対側性入力は、台形体核経由で、小球型細胞globular cellsから間接的に入る。内側核における情況を異なり、すべての聴音可能は周波数が複現されている。ここには厳格は(はっきりした、きびしい)音局在順序tonotopical orderありて、低周波数は外側に高周波数は内側に複現されている。ここのニューロンは両耳間の強度intensityの差によく反応する如し(Boudreau と Tsuchitani, 1973)。外側上オリーブ核は、それ故に、空間における音の局在にも関与しようが、内側核とは対照的に、高周波数音に最もよく作用(動)するようである。この外側核は下丘中心核に両側性に投射する(Stotler, 1953; Adams, 1979)そして恐らく外側毛帯の背側核へも(同側性に)投射がある(Elverland 1978)(9-4図)。
上オリーブ周囲核は多くの種々の細胞型を含み、あるものはコリン作動性で他は明らかにそうでない。これらの核は、恐らく大部分が下行路に関係しており、後述ここでは、下丘からの繊維を、それに恐らくタコ型細胞を含む蝸牛核ニューロンの数種のタイプから側枝による入力をうけていることをのべれば充分也。
外側毛帯核は、大部分が外側毛帯の繊維束内にある細胞群である。通常2つの核が腹側と背側に区別される。上行路の中継核の如くなるも、確かなる知識は乏しい。機能不明。聴覚ニューロンの相当数を全体で構成している。外側毛帯核の腹束核は、恐らく側蝸牛核および両側上オリーブ複合核からを含む(めて)、種々の源泉からの毛帯繊維の側枝の支配を受けている(Warr, 1969; 他)。そして同側下丘の中心核へ投射している(Adams 1979)。背側核の方は、これと対照的で蝸牛核支配をうけてないいだろう。ARG法によれば背側核は外側上オリーブ核から両側性に繊維をうけるようである(Elverland, 1978)。この核は、両側性に上丘中心核に投射し、その繊維で外側毛帯の交連中を交叉し走るものもある(9-4図)。
中脳被蓋の一部、重要なる聴覚(中枢)センター、腹方は背側中脳被蓋に接しているが、下丘は、この背側中脳被蓋に形態的にも機能的にも関連をもっている。背方部では、下丘交連により両丘が互いに結合している。下丘は、外側毛帯の繊維と内側膝状体へ向かう下丘腕の繊維の間にある聴覚路における中継核である。聴覚構造物一般と同様に、外側毛帯を下丘腕も共に脳幹の表層部に位置している。下丘の機能的役割は多くの点で視覚系における上丘のそれと異なっている。すなわち(しかして)下丘は、中脳レベルに到達するすべて(又はほとんどすべて)の上行および下行聴覚繊維の(にとって)中継されねばねらぬ部位obligatory relay stationとして聴覚路の中で鍵をにぎるポジションを得てる。
全体として、下丘は、背側及び腹側蝸牛核、上オリーブ核、外側毛帯核それに聴覚領皮質から繊維をうけている。下丘は、内側膝状体、中脳被蓋、中心灰白質、下オリーブ周囲核、背側蝸牛核、および背外側橋核にその繊維を与える。これらの結合は、同側性か対側性か又、両側性のものもある(Fig. 9-4,9-5をみよ)。通常、しかしながら、下丘への上行性投射は対側の耳からのものが優位で、反対に、下丘下行投射は同側優位でこのようにして対側の耳からの皮質複現をfavor(支持、えこひいき)している。下位聴覚センターから下丘に至る繊維はすべてその中心核(後をみよ)に終わるが、一方
下位のどの部分でもMGBへの上行性投射に参与する。そうはいっても、下丘の種々の亜域は内側膝状体内に異なる標的(終止部)をもっている。前のところで述べたように、中脳レベルおよびそれより高位レベルで、聴覚路はa core and a belt投射に分かたれる。中核(となる)(の)投射は下丘の中心核から起こる。他方周辺帯投射は、中心核をとりまく末梢部(周辺部)の灰白質から起こる。後述されるように、この周辺部灰白質は聴覚皮質からの下行繊維の標的であるという点でも、中心核と異なる。(cp. p.630,皮質線維は下丘pericentral核に両側性に終わる)。
下丘中心核の細胞はまづ第一に外側毛帯の繊維によってactivateされる。この核は毛帯繊維と平行に向かっている平円盤状細胞が層状に配列することが特徴となっている。ゴルジ法(サル、ネコ、ヒトを含めて)で示されている。fibrodendriticな層構造に相応する(同)等周波数の層isofrequency lamellaeが存在することを生理学的に特徴ある周波数をマッピングして証明した(Roseら、1963;他)。中心核に表面から微小電極を挿入して深部に押しし進めていくと、低→高へと振動数が規則的に漸次特徴的に進んでいく(a→b.左図)→
中心核からの主たる遠心性投射はMGBの腹側部の層構造部分laminated portion (Moore と Goldberg, 1963, 1966; Casseday, Diamond と Harting, 1976; Oliver と Hall, 1978a; Kudo と Niimi, 1978)であり、ここは更に第1次聴覚野に投射する。この投射はのちに考察されるように単に、周波数の判別のみならず。他の聴覚器脳にとってもきわめて重要であるに相違ないということは中心核の解剖学的構成についてしられているところ軌を一にする。
毛帯入力→中心核は、多くの核から、
dorsal & ventral cochlear nuclei
medial & lateral superior olives
ventral & dorsal nuclei of lateral lemniscus
外側毛帯核を恐らく除外(例外)として、以上の核内すべて音局在性あり、又、それらの投射にもあり。両耳複現が両下丘にあるも対側優位、中心核→MGB v. (laminated p.)は両側性、同側優位、又、中心核はMGB m(内側核medial division)と中脳被蓋に両側性に下丘の中心灰白質に同側性に繊維を与える。反対側への投射投射は(下)丘間交連を正中交叉する。下丘の周辺灰白質 区分研究者により異なる(総説としてHarrison と Howe, 1974aをみよ)。しかし一致ある如く。中心核の外側に独立した核、外側帯域又はexternal nucleus外核。Morest (1964a)によると、この核は中脳被蓋の一部である。他はpericentral nucleus中心周囲核(Casseday, Diamond と Herting, 1976により言及)、で、これは、この説明では、中心核の背内側部をも含み、5層皮質構造a five-layered cortical structureとしてGeneic と Morest (1971)によって記載された。
上述の如く。外核と中心周囲核とは繊維結合が中心核と可成り違う。外核と中心周囲核は、のこりの中脳被蓋域と共に、聴覚性入力を下丘腕経由の繊維により中心核からうける。この2核は、体性感覚インパルスsomesthetic originを伝える脊髄視床路と内側毛帯の繊維をうける(van Noort, 1967; Schroeder と Jane, 1971; Oliver と Hall, 1978a)、更に下行路のところでのべるが聴覚領皮質からの投射をうける。これらの核は、聴覚性周辺帯投射の一部としてMGBの非有層部に投射する。(もっと詳しくは、Olive と Hall, 1978a,bのHRP研究をみよ)。更に周囲核と外核は上丘の深層に繊維を送る(HRP研究、Edwards, Ginsburgh, Henkel, と Stein 1979)。これらの層は又、とりわけ、聴覚皮質(後述)や視覚や体性感覚に関連するいくつかの脳幹のセンター(核)(中心)から入力をうける(Edwardsら、1979)。(上丘について)多くの遠心路のうちに、視蓋網様体路(Kawamura, Brodal と Hoddevik, 1974)と視蓋橋核路(4,5章と7-31図を参照)がある。後者(?)は加えに、下丘から直接繊維をうける(後をみよ)。視蓋橋核路は背外側橋核(Kawamura と Brodal, 1973)における中継点を介して、小脳虫部における聴視覚領野を含む中脳から小脳への遠隔受容teleceptiveインパルスを導く多くの経路のうち1つを構成している(Hoddevik, Brodal, Kawamura と Hashikawa, 1977)。虫部と室頂核からの出力は複雑であるが、上丘や(Edwardsら、1979)内側膝状体の非有層構造部(Carpenter, 1960)への直接投射が核から。後者(MGのnonlaminated portion)は、上丘からも直接の繊維をうける(AltmanとCarpenter, 1961; OliverとHall, 1978a)。それに、前述したが下丘からも。下丘上丘はこのように就中(among other things)小脳を含む神経回路に関与している。この神経回路のもつ機能的意義については理解が不足であるis poorly understood。しかしながら、一極が驚愕反応。他極が言語speechの如き聴覚性(又は視覚性)にguided導かれる運動機能における役割を示唆することは理のある所reasonableであろう(Fadiga と Pupilli, 1964; Mortimer, 1975)。
下丘→pons. 投射少が下丘のすべての部分から(Kawamura と Brodal 1973; Kawamura, 1975; Aitkin と Boyd, 1978) 。下丘から中心灰白質への投射(Moore と Goldberg, 1963; Matano と Ban, 1976)があるので下丘の(と)自律神経系のとの関連も示唆されている。しかしながら、中心灰白質と中脳被蓋は霊長類や他の動物で共に証明されたように種特異性の呼声(call)のproduction産出関与している如くである(Jurgens と Pratt, 1979をみよ)。下丘の上オリーブ核と背側蝸牛核への下行性投射については下行路のところで一緒に考察される。
視床の核でその尾部、柔膜下面に在る。ヒトではLGBの内側、ネコではLGBの腹側。下丘腕と聴放線間の中継核、内包の後部を経て聴覚領へ放線。
BGMの層状構造部→聴覚領の顆粒皮質koniocortexへ、中核投射の一部として
非層状構造部→視覚領の皮質周辺帯領 周辺投射の一部として、
全く正確にこの上行聴覚領者に平行して反回性recurrent皮質投射あり(後述)MGBには、より上衣又は下位レベルと対照的に、交連繊維はない。膝状体皮質投射は完全に同側性。MGBは細胞構築と繊維結合より亜核に区別されるが普通の組織切片で容易には区別しにくく区別にも又、posterior thalamic群に対しても境界と一定の了解一致がみられぬ(reviewsについてMorest, 1964b; Harrison と Howe 1974bをみよ)。区分に一つにネコGolgiをGolgiをbaseとしたMorest (1964b, 1965a)のものが形態学電気生理学的研究にもっとも有益(更にはOliverとHall, 1975, 1978aを)Morest (1964b)の分類は、3つの主な領域に、
内側核(部)divisions medial
背側核(部)divisions dorsal
腹側核(部)divisions ventral (lam.)(lam. V.)およびnon-lam. V.
(註5)VとDは大略Rioch (1929)のprincipal division (1929)
MとDは大略Riochの大細胞部に一致する。
dorsalとventralは更に各々、3つの亜核に、腹側核(V)は更に大きなventral核と2つの小核からなるventral核はMGBの他のすべての亜核と明瞭な層構造を有する点で区別される。 ∴laminated portion of the ventral division(lam. V. in 9-4, 9-5図)として言及される。
この層構造は主たる細胞(tuftedふさ状の)構造突起パタンと求心繊維の方向性により目に付く(Morest, 1971, 1975; Majorossy と Kiss, 1976a,bをみよ)。lam. V.ネコ、サルでの周波数マッピングが示唆する処では、下丘の中心核と同様に、蝸牛の空間的複現に関連し、
尖端部(低周波数)→外側部
基底部(高周波数)→内側部(cp. 9-4図)
この部のニューロンは下丘中心核から起こるニューロンで活動化。このlam. Vは、相互的な皮質視床繊維(後出)を除けば、他の領域からは、ほんの僅か、もしあったとしての繊維しかうけない。故に優勢的に聴覚性の構造物である。これはAIに(auditory koniocortex)投射する。聴覚・中核投射の最終結合路として。厳密に音局在性を有する投射也。
以上のlam. V部以外の内側膝状体の領域はすべて非有層部で内側部、背側部、および腹側部非有層部からなり。亜核分類と結合関係は複雑であり、機能的にもよく判らぬ。
内側部は大細胞を含みしばしば大細胞部とわあれる。これは多少の繊維を下丘中心核からうける。この繊維を除くと、非有層のMGB領は、すべて聴核インプットを下丘のbelt領から、すなわち、中心周囲核・外核・中脳被蓋からうける。聴覚入力の他に、内側部と背側部は、脊髄視床路、内側毛帯(Nauta と Kuypers 1958; Lund と Webster, 1967a,b,; Oliver と Hall, 1978b)および上丘腕(Altman と Carpenter, 1961; Morest, 1965b)からの繊維(又は側枝)を通じて、他の感覚様態によっても影響される。前に述べたように、MGBのこれらBelt areasは、前庭小脳や小脳虫部の聴視域に関連する室頂核からの繊維もうけている(Carpenter, 1960)。単一ニューロンに体性、前庭、聴性入力が収斂するという電気生理学的証拠がある(Blum, Abraham, と Gilman, 1979)。この機能的意義は解明されるべく残っているが、このnet effect(正味の効果)は、MGBの標的である聴覚領に必然的に(必ずや)伝えられていなければならぬ。
決定は細胞構築、視床との結合、深いbarbiturate麻酔下で誘発電位。ヒト、サル側頭葉の上壁、シルビウス溝内、ネコ、suprasylvian溝よりも下(腹側)の半球外側面に異変あり。完全に順序立てられた可聴周波数の複現をもった顆粒皮質。このkoniocortexは、逆方向の、しばしばより不正確な周波数再現を示すbelt areasにかこまれている。(Woolsey, 1960)。ネコで最もよく研究されている。
以下ネコで。サル・ヒトをbrieflyに。
Rose (1949) comprehensiveな細胞構築学。
AI、小形細胞、多数の細胞部、顆粒皮質koniocortex belt fieldsに囲まる(隣接域への移行をこれら周囲帯細胞構築像を示す。SF, AII. Ep. (Rose)その後電気生理学的記録法で、I (insular), T(temporal area), AIII(SIIの一部含)がWoolsey (1960, 1964)により含めらる。以上域すべてMGBより繊維をうける。core projectionとbelt projectionの2者はこの最終投射である視床皮質結合では完全には独立して分離していない。AIはMGBの有層部からの中核投射の標的であるが、MGBのいくつかの非層構造部分からも又繊維をうける。聴覚領は収斂と発散があり(繊維結合上)、AIを含めてすべての聴覚皮質域はMGBの1つ以上の部分から繊維を供給される。ネコのAI域は音周波数の厳密な空間的再現あるという特徴を示す。脳軸に対し垂直の周期性帯あり。音周波数(低音)が後方(Fig. 9-9の矢印)。
Tunturi (1950)イヌ音局在構成を示した。深麻酔下にストリヒニン法。
Echolocating bats. Doppler-shifted echoの特定の周波数が餌物をlocateするに用いられ、皮質表面上広域を占めることを、めずらしいこととして付記しておく(Suga と O’Neill, 1978)。
音の強度の空間的再現。in Dog AI, Tunturi (1952)によって最初に提唱、Jones と Powell (1970a)サル、視覚、体性知覚系同様にStepwise, outward progression of connections
AI⇔belt areas⇔area 8a⇔area 9⇔areas 10,12
⇔area 22⇔sup. temp. sulcus.(Brodalはsupratemporal sulcus)
ここは、multisensory regionsで聴覚情報が視覚・体性知覚入力と統合される。
ヒト:サルよりももっと判らぬ。シルビウス溝内のsupratemporal平面にある。
gyrus temporal transversus(横側頭回)(Heschl’s gyrus)と尾方にあるplanum temporale(側頭平面?)(temporal plane)(新見によると)AI.横側頭回の内側部(41野)area 3のように顆粒皮質。belt areasは横側頭回の外側部(42野)これにつづいて上側頭回(22野)(cp. この後部に39野ありて22後部+39(角回)=感覚性言語中枢)
von Economo と Horn (1930) の後半に亙る細胞構築学的研究によれば、ヒト大脳皮質の聴覚野は、横側頭回(Heschl回)と後部につづくplanum temporale側頭平面を多少とも占める。大体ブロードマンの41,42に相当する(p791. 12-2図をみよ)。von Economo と Hornは2つの感覚域を記述した。すなわち、
内側域(TD, area supratemporalis intercalata)
外側域(TC, area supratemporalis transversa) (9-9図をみよ)。
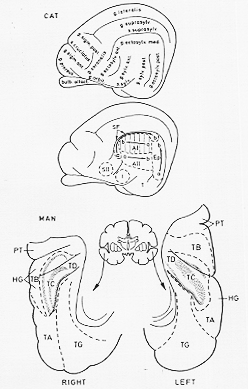
FIG. 9-9 Diagrams of the auditory cortex in the cat and man. In the upper drawing the upper dwawing the main gyri and sulci on the lateral aspect of the hemisphere of the cat arfe shown and labeled according to Hassler and Muhs-Clement (1964). The second drawing shows the auditory cortical areas of the cat, as defined by Rose (1964). The second drawing shows the auditory cortical areas of the cat, as defined by Rose (1949). These are the primary auditory area (A I), the secondary auditory area (A II), the posterior ectosylvian area (Ep), and the suprasylvian fringe sector (SF), with the probable addition of the insular area (I), the secondary somatosensory area (S II), and the temporal area (T). The spatial representation of tonal frequencies with each area is indicated by arrows pointing from low- (a) to high- (b) frequency regions. Vertical lines in A I in dicate isofrequency strips. Below are shown the right and left supratemporal planes of the human brain (simplified from Fig. 5 of von Economo and Horn, 1930). The auditory cortex is composed of two fields, TD (area supratempooralis intercalata) and TC (area supratemporalis transversa). On the right side these occupy a double transverse temporal (HG: Heschl's) gyrus; on the left they correspond to a single Heschl's gyrus and a part of the more caudally situated temporal plane (PT). Areas TD and TC are composed of markedly granular (dotted) and less heavily granular (blank) subareas. Note distinct right-left asymmetries, with a larger temporal plane on the left side. Abbreviations: TA: area temporalis superior; TB: area supratemporalis magnocellularis; TG: area temporapolaris.
この2域の類同性は不明。しかしながら、これらが顆粒皮質及び皮質周辺帯域を共に含むか否か不明。TDとTCは構造上似ているようだ。
PT: temporal plane (planum temporale)側頭平面cp. IX-19(下).
HG: Heschl’s gyrus (transverse temporal gyrus)
TB: area supratemporalis magnocellularis
TD: area supratemporalis intercalata
TC area supratemporalis transversa
TA:: area temporalis superior
TG: area temporopolaris
ヒトHeschl回の電気刺激(Penfield と Jasper, 1954) ringing, humming, clicking, buzzing etc対側の耳にもっともしばしばきこえる。Pfeifer (1936)側頭葉傷害例を研究してもしヒトにtonotopical局在ありとせば高音は内側に、低音は外側に復現されている筈と結論した。これはサル、チンパンジーの所見と完全に一致する。von Economo と Hornは最初にsupratemporal planeの大きさとconvolutionの差異variationと左右差大なることを指摘した。Heschl回はsolitaryのもdoubleのもある。
感覚野皮質の柱状構造は近年大いに注目されてきた(8章、9章をみる)。感覚野ともcolumnar organizationがあるといういくつかの実験的証拠がある。Merzenich, Knight と Roth (1975)の電気生理学的研究によれば、一次感覚皮質のisofrequency stripsは恐らく皮質全層のすべてのユニットに対して同じ特徴ある周波数をもったisofrequency columnsを記述された。これは、両耳性の加算summationか又は抑制inhibitionを示す(Imig と Adrian, 1977; Middlebrooks, Dykes と Merzenich, 1978)。この加算と抑制の柱は、同周波数条(又は柱)を横切る方向に相互の帯alternating bandを占める。両耳性加算柱の内に、小さい(部分の)minor同性又は対側優位柱dominance columnのモザイクが存在する。
9-5図(in p.611をかっておく) cp. 9-4図(in p.610: 上行性聴覚路)(IX-7をみよ)
最終更新日:2002/09/13
