
川村光毅
| まえがき | |
| I.小脳の帯状構造-その概念の変遷 | |
| II.前葉にみられる帯状構造 | |
| III.下オリーブ核について | |
| IV.オリーブ小脳線維と小脳帯(細)域 | |
| おわりに | |
| 文献 |
菱脳峡(isthmus rhombencephali)を境にして脳全体は大脳を含む前方部と菱脳を含む後方部の二つの基本部分にわけられる。小脳は胎生期菱脳の翼板(alar
plate)の背外部が内方へ曲って形成された菱脳唇から背方向に発達したもので、オリーブ核や橋核は翼板にある他の細胞群が腹方に移動して形成されたものである。哺乳動物の脳でとくに明らかなように、小脳は正中部に虫部と左右の半球部からなるが、系統発生的にみれば、前庭神経の原始的な延髄核から、または、第一次の前庭域から発達したものである。下オリーブ核群には脳内の種々の領域からの神経線維が核内の特定の部位に主な終止域をもって入ってくる(後述、図9参照)。一方、オリーブ核から起こるオリーブ小脳線維は恐らくすべて登上線維として小脳皮質分子層のプルキンエ細胞樹状突起に(分枝は小脳核などにも)終わる(註:カエルの第一次前庭神経線維には苔状線維と登上線維の2系が存在するとされているが28、39)、少くとも高等脊椎動物では、オリーブ核以外の部位から登上線維が起こるという確実な証拠は現在得られていない)。
さらに、大脳皮質の発達にともなって小脳皮質との間の中継核として橋核が発達してくる。橋核細胞からは苔状線維が起こり、顆粒細胞に終わる。このように、小脳皮質内の唯一の出力ニューロンであるプルキンエ細胞は、単純にいって、下オリーブ核から起こる登上線維からの入力を直接的に、オリーブ核以外のいわゆる小脳前核から起こる苔状線維からの入力を顆粒細胞に介して間接的にうけている。
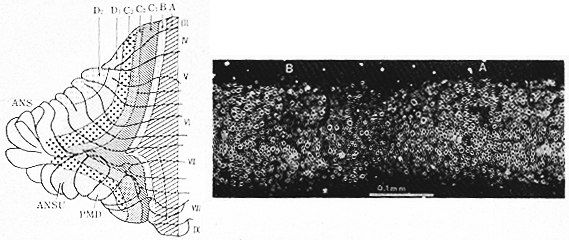
図1 左図:哺乳動物の小脳で見られる前後方向の帯状区分を示すVoogd56)の模式図(シロイタチ小脳の背側面)。上図:A域とB域の境界部。シロイタチ小脳髄質。Haggquist染色。(Voogd56)より)
小脳の形態的区分については、Larsell38)の詳しい記載がある。また、機能的には前庭小脳部(vestibulocerebellum)、脊髄小脳部(spinocerebellum)および大脳小脳部(cerebrocerebellum)と一般的に分けられてきた37)。この稿の主題である小脳にみられる帯状構造、あるいは、縦割り構造を示した最初の研究は、皮質から小脳核への投射に三つの帯域を区別したJansenとBrodal30、31)のものであろう。彼らは小脳皮質の虫部が室頂核(内側核)へ、中間領域が中位核へ、そして、外側領域が歯状核(外側核)へそれぞれ投射するというパターンを示した。その後、損傷後に現れた症候を観察したChambersとSprague14、15)の仕事によって支持された。
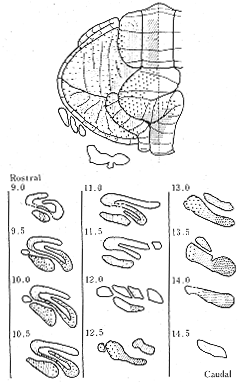
図2 小脳皮質刺激後、逆方向性電位をオリーブで記録することにより明らかにしたネコのオリーブ小脳投射の模式図。マークのない部分は調べなかった部分である。(Armstrong, HarveyとSchild6)より)
1964年、Voogd55)は小脳の正常標本および求心性線維と遠心性線維の実験的研究をもとにして、ネコの小脳で前後方向の構成パターンをはっきり示した。虫部ではほぼ矢状方向に、半球部では外側に凸に帯状域がみられる。さらにVoogd56)は、小脳の帯状に内側から外側にA,
B, C, D,
の名称を与えた。この帯域のあるものはさらに分けられた。(図1のシロイタチのHaggquist髄鞘染色による上の正常標本と、左の模式図をみよ)。彼によれば、白質は皮質求心性の小径線維束("raphe")と皮質遠心性の大径線維束の交互に帯状に並んだ集団からなる。そして、ある種の求心性(または遠心性)の結合はある特定の帯域に終わる(または、から起こる)。この原則は生理学的研究によっても確かめられた。すなわち、Armstrong,
HarveyとSchild6)は小脳の種々の部位を電気刺激して、下オリーブ核内において逆方向性電位の局所的分布を記録する方法を用いて(図2)、また、Oscarssonのグループ41,42)は腹側脊髄オリーブ路(VF-SOCPs)や後索核経由でオリーブに入る脊髄からの神経路(DF-SOCPs)の種々の成分を電気的に記録する方法を用いて(図3)、登上線維の帯状終止域を小脳皮質内に示した。最近オートラジオグラフ法を用いた研究26、27、32)により、オリーブ小脳投射の構成パターンが、一層明らかにされたが、この線維群はA,
B, C1, C2,C3, D1、D2、の帯域に終止する(図4、図5)。また、Brodal,
WalbergらOsloグループ10,12,13,36,58)はhorseradish peroxidase(HRP)法を用いて同様にオリーブ核と小脳との間の結合関係を明らかにし、小脳皮質に帯状構造が存在することを示した。
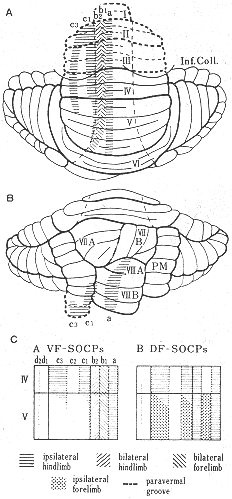
図3 ネコの小脳前葉(A図)およびVII小葉と正中旁小葉(B図)における5つのVF-SOCPs(後索-脊髄-オリーブ小脳路)の終止帯域(a,b1,b2,c1,c2,c3)を示す模式図。C図:VF-SOCPs(左)とDF-SOCPs(後索-脊髄-オリーブ小脳路)(右)の種々の成分を刺激したのち、前葉のIVとV小葉に生じた登上繊維反応の帯状分布をしめす図。(OscarssonとSjolund46,47)より)
このほか、Chan-Palayら16)はオートラジオグラフ法を用いて下オリーブ核から(登上線維)と外側網様核から(苔状線維)の小脳皮質への両側性帯状投射をラットで示した。サルでの報告はまだみられないが、ネコやラットに存在する帯状投射分布のmappingを半球部ですることは、半球が外側に拡大しているために多少の困難性をともなう。したがって、この意味からも、ラットやクジラの胎児を用いた発生学的研究で、その初期に虫部のみならず半球部にも明瞭な縦割りのパターンが認められていることは34,35)、非常に興味あることである。
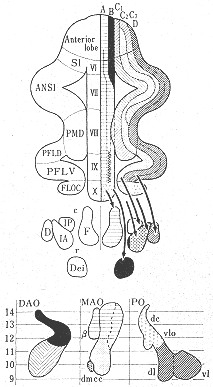
図4 軸索変性鍍銀法およびオートラジオグラフ法を用いて研究したネコのオリーブ小脳投射にみられる帯状パターンの模式図。FLOC;片葉、PFLD;背側旁小葉、PFLV;腹側旁片葉、PMD;正中旁小葉、SI;単小葉。(Groenewegen, VoogdとFreedman27)より)
本稿で、帯状構造とか矢状方向または前後方向の帯域、縦割り構成などの言葉を用いるが、元来(para-)sagittalないし、longitudinalという用語を小脳全体に用いるとなると、厳密な意味では必ずしも適正な言葉とはいえない。オリーブ小脳投射の終止帯域の表現に使われる場合も小脳小葉の長軸に直角の方向に現われる帯域の表現に正しくあてはまるのは、前葉虫部や小節、正中旁小葉の一部に限られるからである。しかし、この術語の使用を誤法とする見解は完成した形からのみみている面があり、われわれはこのsagittal,longitudinalという用語を一般的に承認されてよい言葉と考えている。
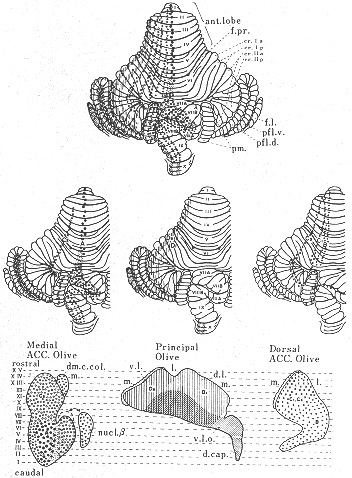
図5 オートラジオグラフ法による実験結果をもとに、HRP法や電気刺激法などの所見を加えて検討した結果得られたネコのオリーブ小脳投射を示す模式図。(KawamuraとHashikawa32)より改変)
以上みてきたように、小脳の皮質にも髄質にも帯状構造が存在することは承認されるに至った。1940年代には虫部、中間部、外側部と3帯域にmacro的に分けられていた概念が、研究が進むにつれてますます細分されるようになった。60年代に入って、A,B,C,Dの4域に、60年代後半期にはさらにC域がC1,C2,C3,の3亜域に区分されるようになり、D域もD1とD2の2域に分けられて70年代に入り、A域とB域もそれぞれ二分46,47)されるに至った。Oscarssonらのa(a1とa2)b(b1とb2),C1,C2,C3,d1,d2,の矢状帯はVoogdが提唱した形態上の帯域(A,B,C1,C2,C3,D1,D2,)とよく一致する47)。ごく最近、Oscarssonグループはaとbとの間にx域21)をみいだしたり、b1とb2との間にみられる重複22,46)についても論じている。また、AnderssonとOscarsson1,2)によってb域が五つのmicrozonesに区分されるにおよんで、zonesの40年代のmacro的概念は過去のものとなりつつあり、幅200~600μm単位のzonesの概念を越えて、さらに細かい単位のmicrozonesを実際の存在としてとらえて考察することができるようになった。
小脳への苔状線維求心系や皮質構造の均一性の問題にたちいることをせず、ただちに、前葉へのオリーブ小脳路の構成をみながら小脳の帯状構造について考えてみたい。この部分は小脳のうちで研究のもっとも進んでいる領野の一つである(註:他の領域についての考察はBrodalとKawamura11)の綜説を読まれたい)。
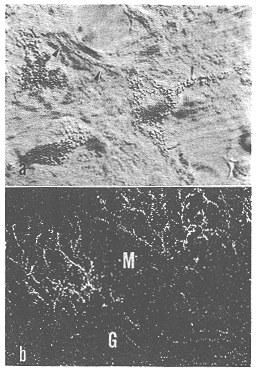
図6 オリーブ小脳繊維の起始細胞像(a;オリーブ細胞のなかに多数のHRP顆粒が見える。矢印は軸索。干渉顕微鏡。×1,300)と分子層(M)にみられる標識された登上線維の最終分枝像(b;×150)
最初に、トリチウムで標識されたアミノ酸(プロリンとロイシン)をネコの下オリーブ核群の前方部と後方部に微量(0.3μl,6μCi)注入した2例32)の小脳皮質内にトレースした登上線維の終末分布を示す(図6b,図7)。登上線維の終止域は明らかな前後方向の帯域を示している。前葉の出現したこれらの帯域の幅は、A域が200~400μm,B域が400~600μ,C1,C2,C3,D1,とD2,のそれぞれの亜域が200~400μmである。
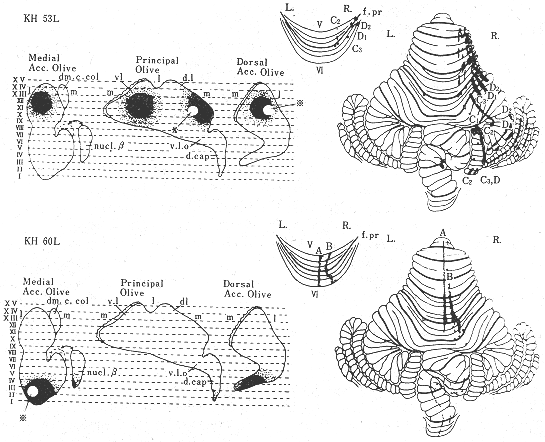
図7 下オリーブ核の前方部(上:ネコKH53L)と後方部(下:KH60L)に3H-アミノ酸を注入したのちにみられた登上線維の皮質内分布を示す模式図。(KawamuraとHashikawa32)より)
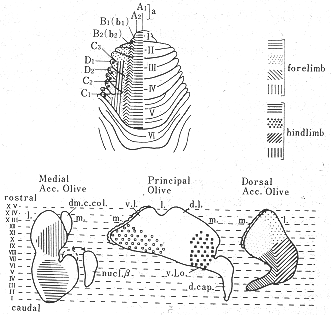
図8 現在入手できるデータをもとに作成した前葉へのオリーブ小脳投射の模式図。C帯域投射のうちC1+C3域とC2域が前葉の前方部に存在しないことに注意。(BrodalとKawamura11)より)
最近のオートラジオグラフ法26,27,32)およびHRP法12)を用いた研究(図6a)、さらには生理学的研究結果6,46,47)を詳細に比較検討した結果得られた下オリーブ核から前葉への投射の模式図を図8に示した。この図にみられるように、虫部では内側帯は内側副オリーブ核の尾半部から、外側の帯は背側副オリーブ核の尾外側部からそれぞれ求心線維をうける。オリーブ核内の起始細胞の位置から考えてこれらの両帯域はVoogdのA域とB域に相当する。半球部では、主オリーブ核から外側のD帯域へ投射する。内側副オリーブ核の吻側半からと背側副オリーブ核の吻内側部からは中間部域(C域)へ投射する。これらの内側および背側の副オリーブ核から線維をうける終止帯域は、それぞれ、C2域およびC1プラスC3域に相当する。主オリーブ核からの投射についていえば、背側板(dorsal lamella)からはD1域に腹側板(ventral lamella)からはD2域に線維を送っている。C2,D1,D2,の帯域はI, II小葉にはみられない。小脳皮質を電気刺激してオリーブ核内の起始部位を逆方向性の反応を指標として調べることによってオリーブ小脳投射系を研究したArmstrongら6)の結果(図2)では、恐らく電流滑走(current spread)のために、B帯域へ終わる線維の起始部位が内側副オリーブ核にも同定されたり、C2帯とD帯が共に主オリーブ核の腹側板や内側副オリーブ核の吻側半にも存在するという所見も出たが、基本的構成パターンは図8と大体一致する。Oscarssonと共同研究者たちのIV小葉とV小葉の各帯状域にみられる前肢と後肢の再現領域(representation area)の模式図(図3)も、下オリーブ核への求心路についての考察結果(図9)から推察される体部位局在の領域とよく一致する。すなわち、Oscarssonの脊髄-オリーブ小脳路の研究によれば、VF-SOCP(前索-脊髄-オリーブ小脳路)経てc1とc3(C1とC3)域にくる入力は、下肢域からのものでV小葉には達しない(図3C)。DF-SOCP(後索-脊髄-オリーブ小脳路)を経てc1+c3域に入る前肢と後肢からの情報は、それぞれV小葉とIV小葉に到る。すなわち、小脳皮質の終止域に前後の体性局在の関係がみられる。これは、背側副オリーブ核の吻側部(ここからc1とc3域に投射する)への前肢と後肢からの入力がその異なる部位に終わるという事実に符合する。A域への起始域は内側副オリーブ核尾側半の中央部にあり、この領野に脊髄からの入力は後肢域から入る点で、A(a)帯域の後肢域再現(図3C)は符号するが、前肢域からの入力もあるのでA域の外側部とx帯21)(註:現在,x帯はa帯に属すとも確定されていない)も含めて前肢域の再現域も存在すると思われる。VoogdのB帯は背側副オリーブ核尾外側部から起こるオリーブ小脳線維をうける領域で、ここは直接に脊髄から、また、間接に後索核を介して後肢から入力をうける領域であるが一部前肢にも関係している(図9中央図)。ともに後肢に関係する領域であるa帯とb2帯との間にOscarssonとSjolund46)はVF-SOCPsとDF-SOCPsから前肢のみの入力をうける領野(b1)を区別した。b1という名称を与えたことは、彼らがこの入力は背側副オリーブ核を介すると考えていることを意味する。この副オリーブ核は外側部が後肢に、内側部が前肢に関係する脊髄からの入力をうけている(図9中央図)という事実に合致する。C2帯の起始領域である内側副オリーブ吻側半の部分には、脊髄からの入力はほとんどみられず(存在するとしてもわずかなものが楔状束核と三叉神経脊髄路核から)、むしろ、大脳皮質から少し入るのみである(図9上図)。OscarssonとSjolund46,47)のVF-SOCPsとDF-SOCPsの図表(図3)ではc2帯域が空白となっていた。ただし、最近彼らが発表した図表ではc2帯(V小葉)に前肢と後肢の入力が両側性に入るとしている21,45)。
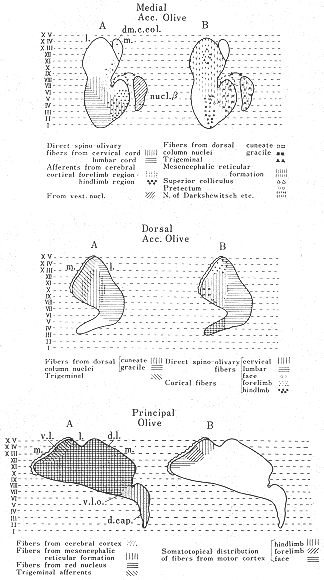
図9 ネコの内側副オリーブ核(上図)、背側副オリーブ核(中央図)および主オリーブ核(下図)への主な求心線維の終止分布域を示す簡略な図。(BrodalとKawamura11)より)
以上みてきたように、A(a)帯とB(b)帯では体性部位局在(前肢と後肢という大きな範囲で)の配列は、左右に内側と外側の方向に存在するが、一方、C1とC3の両帯域ではその配列が前後に大体VとIVの小葉間あたりを境にして存在する。C2帯には、すぐ上にみた通り、局在配列がみられない。D帯に関しては、その起始領野である主オリーブ核が脊髄レベルからの入力をうけない(図9下図参照)が、DF-SOCPsからの入力がd1とd2に入る21)。これは、現在不明であるが、脳幹のどこかの中継核を経由するものと思われる。
このように、オリーブ小脳路の終止領域に関して、ネコで、形態学的に大きく分けてA,B,C1,C2,C3,D1,D2の7帯の帯状構造が存在することが証明され、生理学的にも前葉において少くとも10個のzonesが区別されている。先にのべたようにb域がさらに五つのmicrozonesにまで分けられるに至った1、2)。b域だけではなく他の帯域にも同様のmicrozonesの存在が今後の研究によって明らかにされることが十分予想される。
ここで、小脳のもつ帯状(細)域の構造の主要な部分を構成していると考えられる登上線維の起始核(細胞体の集団)、すなわち、下オリーブ核群について二、三ふれておきたい。
Nissl標本でみると、オリーブ核の神経細胞は全体として小型で、多角形や西洋ナシ形のものが多いようにみえるが、細胞の大きさや分布の密度に関しては部位的にかなりの差異がみられる。たとえば、ネコの主オリーブ核の腹側板では少数の細胞が低い密度で分布している。ネコの1側のオリーブ核群には約70,000個の細胞があるという23、41)。Golgi標本で観察すると、異なるタイプの細胞が区別され52、53)、長い分岐の少ない樹状突起をもったもの(isodendriticタイプに相当)と、多数の分岐した短かい樹状突起をもったもの(allodendriticタイプ49,50)に相当)がある。これらの移行型もみられる。isodendriticタイプは一般に他のタイプよりも未熟で分化の遅れたニューロンのタイプと考えられている。Golgi法を用いた研究53)によれば、この二つのタイプのオリーブ核内分布には部分的差異がみられる。たとえば、主オリーブ核にはisodendriticタイプの細胞は認められず、そのかわり、両副オリーブ核、背内側細胞柱(dorsomedial
cell column)、β核や背帽(dorsal cap)にはこれらの細胞が多数みられる。一方、オリーブ核求心線維の軸索終末像にも、①細かい終末枝が"くさむら"状に分岐しているタイプ、②かなり太い終末枝が"ロゼット"状にみられるもの、③多数の副枝(collaterals)をもって細い終末線維などいくつかの種類がみられる53)。また、オリーブ求心路は多く存在するが、これらのオリーブ亜核内にそれぞれ異なった終止分布域をもっている。今までに発表された論文を検討した結果が図9にまとめてある(詳細はBrodalとKawamura11)を参照)。しかし、オリーブ核内で求心線維と遠心ニューロンを同定して――たとえば変性法とHRP法を併用させて――行ったシナプトロジーの研究は非常に少ない33)。これまでのオリーブ求心路と遠心路の研究成果の上に立って、今後上記電顕レベルでの研究を発展させていく必要がある。オリーブの組織化学的研究も期待される40,59)。
下オリーブ核が小脳へ登上線維を送る唯一の神経核であると確実にはいえないが、オリーブ小脳線維が圧倒的多数の登上線維の終末を小脳に供給していることは疑いのないところであろう(註:逆にオリーブ小脳線維が登上線維以外の線維、すなわち苔状線維として全く終わらぬという保証もないのだが)。現在、下オリーブ核以外に登上線維の起始核といわれるものは、少なくとも哺乳類では知られていない。
Cajalの時代から、1本の登上線維は1個のプルキンエ細胞の樹状突起に終わると考えられ、一般的に容認されてきた。したがって、プルキンエ細胞の数だけオリーブ細胞が存在していれば話は簡単であった。しかし、実際、オリーブ細胞とプルキンエ細胞の数を調べてみると、両者の比は、ネコ41)では1:10、ヒトでは1:15(100万個:1500万個)9、23、42)である。とすると、1本のオリーブ小脳線維が分枝をもち、少なくとも数個のプルキンエ細胞をカバーしなければならない。この登上線維の分枝像は、種々の動物で形態学的に認められた25、48、51)。恐らく、1個のオリーブ細胞の軸索は4~5個のプルキンエ細胞を支配する。生理学的にも、同一帯域内に登上線維の分枝となって終止することが逆方向性電位を記録する方法を用いたArmstrongら4~6)によって示された。この形態学的証拠もある7)。また最近、共通の登上線維群が異なる帯域に分枝を出して終わるという若干の生理学的証拠が前葉皮質で報告された20)。
一方、ネコの登上線維の終止帯域間に示された幅400~600μmの帯状の空白域(図5、図7を参照)32)は、ラットにも存在する16)。ラットのオリーブ小脳線維は両側性であるが、同側性と対側性の投射域は重なっている16)。この空白域に存在するプルキンエ細胞はどこからの入力を直接うけているのであろうか?胎生または生後の一時期に登上線維の供給をうけていたのであろうか?下オリーブ核以外から登上線維が起こらないとすると、たとえばモノアミン系の線維(のみ)によって支配されているのであろうか?Falck-Hillarp法24)によりモノアミン線維の終末が小脳分子層にみられるという報告はいくつかある8、17、29)。このオリーブ線維によって供給されない小脳皮質の帯状域に関する満足な説明は今のところない。
以上みてきたように、下オリーブ核から起こる登上線維の終末が小脳皮質分子層に帯状(細)域をつくる。小脳皮質それ自体には、形態上帯状域の集合体を示唆する構造上の単位は認めがたいが、下オリーブ核からの直接の入力の受容領野としてみると、小脳皮質に矢状方向の帯状構成がみられることは明らかである。皮質と核の間にも相互の結合がみられ、ここにも帯状配列様のパターンが存在する19、57)。また、小脳皮質前葉の虫部から前庭神経核への投射系においても、その起始核であるプルキンエ細胞群は、前後方向に帯状に配列している18)。さらに、外側網様核から起こる苔状線維の皮質内終止域においても、その強い投射域は前後に帯状に配列がみられることがラットの小脳で示されている16)。しかしながら、下オリーブ核、小脳皮質、小脳核それに一部の苔状線維系や皮質遠心系を含めて一般化した理論的考察を試みることは、魅惑的ではあるが、現段階では、必ずしも挑戦に価する仕事であるとはいいがたいように思う。さらに細かいデータの集積が必要であろう。とくに、小脳核と苔状線維系に関するデータが不足している。
登上線維と苔状線維の入力系では、小脳内における分布の状況やシナップス結合が全く異なる。小脳皮質の非常に細かい領野にこの二つのタイプの情報が並列的に入ってくることにより小脳へのいわゆる二重支配が成立している54)。プルキンエ細胞に対して、直接的に非常に強い興奮性刺激を与えている登上線維が極めて正確な帯状投射系をもっていることは、小脳の帯状構造についての存在を浮き彫りにさせている。小脳皮質内に恐らく幾万と存在するmicrozonesで種々の分析がなされる過程に直接的影響をおよぼして、その働きを修正することにより適正な結果を与えうる一種の機能単位としての役割が、この内に蔵されていると考えこともできる。しかし、下オリーブ核群内における求心線維の分布様式(図8)をみても、また、オリーブ小脳線維の分岐投射の事実をみても、小脳皮質のある帯域(zone,
microzone)が一つのソースから"純粋"な情報をうけているとは考えにくい。このことは、一つ以上のソースからの入力により一定の小脳領域がactivateされるという生理学的事実によって支持されている(いくつかのデータについてはArmstrong3)をみよ)。
小脳の"帯状構造"についてこの数年間、解剖・生理学的に詳しく研究された。ある程度の強い機能的関連づけが小脳(micro-)zoneと一定のオリーブの領域との間に確立されつつあるように思われる。
主題を前葉にしぼって小脳の前後方向の帯状域(longitudinal or sagittal zones),さらには、帯細域(microzones)について述べてきた。これに皮質核投射、小脳核・オリーブ投射および小脳核-視床核-大脳皮質などの系に関する詳細なデータが今後の解剖・生理学的研究によって加えられたとき、小脳の皮質-核複合体で機能的単位としてのmicrozonesの役割が一層明らかにされてくるだろう。一方、この方面の仕事の進展が現在期待されている小脳内の領域は、前葉のほかに、本稿でふれることができなかった片葉であろう。下オリーブ核、片葉、室頂核、前庭神経核群に関する現在のデータの集積(文献省略)の上に、microzonesの解明に迫る研究結果が発表される日がさほど遠くはないように思われる。
1) Andersson,G. and Oscarsson,O.:Projections to lateral vestibular nucleus from
cerebellar climbing fiber zones.Exp.Brain Res., 32:549-564, 1978a.
2) Andersson,G. and Oscarsson,O.:Climbing fiber microzones in cerebellar vermis and their
projection to different groups of cells in the lateral vestibular nucleus.Exp.Brain Res.,
32:565-579,1978b
3) Armstrong,D.M.Functional significance of connections of the inferior
olive.Physiol.Rev., 54:358-417,1974.
4) Armstrong,D.M. and Harvey,R.J.:Responses in the inferior olive to stimulation of the
cerebellar and cerebral cortices in the cat.J.Physiol., 187:553-574,1966.
5) Armstrong,D.M., Harvey,R.J.and Schild,R. F.:Branching of inferior olivary axons to
terminate in different folia,lobules or lobes of the cerebellum. Brain Res.,
54:365-371,1973.
6) Armstrong,D.M., Harvey,R.J.and Schild,R.F.:Topographical localization in the
olivocerebellar projection:an electrophysiological study in the cat.J.comp.Neurol.,
154:287-302,1974.
7) Berkley,K.J., Brodal,A.and Walberg,F.:Anatomical demonstration of branching of
olivocerebellar fibers by means of a double retrograde labeling technique(in
preparation,1980)
8) Bloom,F.E., Hoffer,B.J. and Siggins,G.R.:Studies on norepinephrine-containing afferents
to Purkinje cells of rat cerebellum.I.Localization of the fiber and their synapses.Brain
Res., 25:501-521,1971.
9) Braitenberg,V.and Atwood,R.P.:Morphological observations on the cerebellar
cortex.J.comp.Neurol.,109:1-33,1958.
10) Brodal,A.:The olivocerebellar projection in the cat as studied with the method of
retrograde axonal transport of horseradish peroxidase.II.The projection to the
uvula.J.comp.Neurol.,166:417-426,1976.
11) Brodal,A.and Kawamura,K.:The olivocerebellar projection.A review.Adv.Anat.Embryol.Cell
Biol., 64:1-140, 1980
12) Broral,A.and Walberg,F.:The olivocerebellar projection in the cat studied with method
of retrograde axonal transport of horseradish peroxidase.IV.The projection to the anterior
lobe.J.comp.Neurol., 172:85-108, 1977a.
13) Brodal,A.and Walberg,F.:The olivocerebellar projection in the cat studied with the
method of retrograde axnal transport of horseradish peroxidase.VI.The projection onto
longitudinal zones of the paramedian lobule.J.comp.Neurol., 176:281-294,1977b.
14) Chambers,W.W.and Sprague,J.M.:Functional localization in the cerebellum.I.Oranization
in longitudinal corticonuclear zones and their contribution to the control of posture,both
extrapyramidal and pyramidal.J.comp.Neurol., 103:105-130,1955a.
15) Chambers, W.W.and Sprague,J.M.:Functional localization in the
cerebellum.II.Somatotopic organization in cortex and nuclei.A.M.A. Arch.
Neur.Psychiat.,74:653-680,1955b.
16) Chan-Palay,V.,Palay,S.L.,Brown,J.T.and Van Itallie, C.:Sagittal organization of
olivocerebellar and reticulocerebellar projections:autoradigraphic studies with
35S-methionine.Exp.Brain Res.,30:561-576,1977.
17) Chu,N.-S. and Bloom,F.E.:The catecholamine-containing neurons in the cat dorsolateral
pontine tegmentum:Distribution of the cell bodies and some axonal projections.Brain
Res.,66:1-21,1974.
18) Corvaja,N.and Pompeiano,O.:Identification of cerebellar corticovestibular neurons
retrogradely labeled with horseradish peroxidase.Neuroscience, 4:507-515,1979.
19) Dietrichs,E.and Walberg,F.:The cerebellar corticonualear and nucleocortical
projections in the cat as studied with anterograde and retrograde transport of horseradish
peroxidase.I.The paramedian lobule.Anat.Embryol.,158:13-39,1979.
20) Ekerot,C.-F. and Larson,B.:Three sagittal zones in the cerebellar anterior lobe
innervated by a common group of climbing fibers.
Proc.Int.Union.Physiol.Sci.Paris,Vol.XIII:208,1977.
21) Ekerot,C.-F. and Larson,B.:The dorsal spino-olivocerebellar system in the
cat.I.Funtional organization and termination in the anterior lobe.Exp.Brain
Res.,36:201-217,1979a.
22) Ekerot,C.-F. and Larson,B.:The dorsal spino-olivocerebellar system in the
cat.II.Somatotopical oranization. Exp.Brain Res.,36:219-232,1979b.
23) Escober,A.,Sampedro,E.D. and Dow,R.S.: Quantitative data on the inferior olivary
nucleus in man,cat and vampire bat.J.comp.Neurol.,132:397-404, 1968.
24) Falck,B.,Hillarp,N.A., Thieme,G.and Thorpe,A.:Fluorescence of catecholamines and
related compound condensed with formaldehyde.J.Histochem.Cytochem., 10:348-354,1962.
25) Fox,C.A., Andrade,A.and Schwyn,R.C.: Climbing fiber branching in the granular
layer.In:R. Llinas (ed.) Neurobiology of Cerebellar Evolution and Development,
p.603-611,Chicago:Am. Med. Assoc.,1969.
26) Groenewegen,H.J. and Voogd, J.:The parasagittal zonation within the olivocerebellar
projection.I. Climbing fiber distribution in the vermis of cat cerebellum.J. comp.Neural.,
174:417-488,1977.
27) Groenewgen,H.J., Voogd,J.and Freedman,S.L.:The parasagittal zonal organization within
the olivocerebellar projection.II. Climbing fiber distribution in the intermediate and
hemispheric parts of cat cerebellum.J.comp.Neurol., 183:551-602,1979.
28) Hillman,D.E.:Light and electron microscopical study of the relationships between the
cerebellum and the vestibular organ of The frog.Exp.Brain Res.,9:1-15,1969.
29) Hokfelt,T.and Fuxe,K.:Cerebellar monoamine nerve terminals, a new type of afferent
fibers to the cortex cerebelli.Exp.Brain Res., 9:63-72,1969.
30) Jansen,J.and Brodal,A.:Experimental studies on the intrinsic fibers of the
cerebellum.II.The corticonuclear projection.J.comp.Neurol.,73:267-321, 1940.
31) Jansen,J.and Brodal,A.:Experimental studies on the intrinsic fibers of the
cerebellum.III.The corticonuclear projection in the rabbit and the monkey.Auh.Norske
Vid.Akad.I.Mat.-Nat.Kl., No.3:1-50,1942.
32) Kawamura,K.and Hashikawa,T.:Olivocerebellar projections in the cat studied by means
of anterograde axonal transport of labeled amino acids as
tracers.Neuroscience,4:1615-1633,1979.
33) 川村光毅、端川 勉:HRP法の再検討とニューロン標識法による神経回路網の研究。脳の統御機能5「活性物質と神経回路網」大塚・伊藤編。p.85-107,医歯薬出版、1980.
34) Korneliussen, H.K.:On the ontogenetic development of the cerebellum(nuclei,
fissures,and cortex)of the rat,with special refence to regional variations in
corticogenesis.J.Hirnforsch., 10:379-412,1968.
35) Korneliussen.H.K.:Cerebellar organization in the light of cerebellar nuclear
morphology and cerebellar corticogenesis.In:R. Llinas(ed.)Neurobiology of Cerebellar
Evolution and Development,p.515-523, Education and Research Foundation,Chicago,Ill., 1969.
36) Kotchabhakdi,N., Walberg,F.and Brodal,A.:The olivocerebellar projection in the cat
studied with the method of retrograde axonal transport of horseradish peroxidase.VII.The
projection to lobulus simplex,crus I and II.J.comp.Neurol., 182:293-314,1978.
37) Larsell,O.:The cerebellum.A review and
interpretation.Arch.Neurol.Psychiat.(Chicago),38:580-607,1937.
38) Larsell,O.:The comparative anatomy and histology of the cerebellum from monotremes
through apes,269 pp., ed.J.Jansen.The University of Minnesota Press,Minneapolis,1970.
39) Llinas,R., Precht,W.and Kitai,S.T.:Climbing fibre activation of Purkinje cell
following primary vestibular afferent stimulation in the frog.Brain Res., 6:371-375,1967.
40) Marani, E., Voogd,J.and Boekee,A.:Acetylcholinesterase staining in subdivisions of the
cat's inferior olive.J.comp.Neurol., 174:209-226,1977.
41) Mlonyeni,M.:The number of Purkinje cells and inferior olivary neurones in the
cat.J.comp.Neurol., 147:1-10,1973.
42) Moatamed,F.:Cell frequencies in the human inferior olivary nuclear
complex.J.comp.Neurol.,128:109-116,1966.
43) Oscarsson,O.:The sagittal organization of the cerebellar anterior lobe as revealed by
the projection patterns of the climbing fiber system. In:R.Llinas(ed.)Neurobiology of
Cerebellar Evolution and Development, p 525-537,Education&Research
Foundation,Chicago,Illinois,1969.
44) Oscarsson,O.:Functional organization of spinocerebellar paths.In A.Iggo(ed.)Handbook
of Sensory Physiology.Vol.II.Somatosensory system,p.339-380,Springer,Berlin,1973.
45) Oscarsson,O.:Functional units of the cerebellum-sagittal zones and microzones.Trends
in Neurosci:,2:143-145, 1979.
46) Oscarsson,O.and Sjolund,B.:The ventral spino-olivocerebellar system in the
cat.I.Identification of five paths and their termination in the cerebellar anterior
lobe.Exp.Brain Res., 28:469-486,1977a.
47) Oscarsson,O.and Sjolund,B.:The ventral spinoolivocerebellar system in the
cat.II.Termination zones in the cerebellar posterior lobe.Exp.Brain Res.,
28:487-503,1977b.
48) Palay,S.L. and Chan-Palay,V.:Cerebellar Cortex,Cytology and Organization.348 pp.,
Springer-Verlag,Berlin-Heidelberg-New York,1974.
49) Ramon-Moliner,E.:An attempt at classifying nerve cells on the basis of their dendritic
patterns.J.comp.Neurol., 119:221-227,1962.
50) Ramon-Moliner,E.:La differentiation morphologique des neurones.Arch.Ital.Biol.,
105:149-188,1967.
51) Scheibel,M.E. and Scheibel,A.B.:Observations on the intracortical relations of the
climbing fibers of the cerebellum.A Golgi study.J.comp.Neurol., 101:733-764,1954.
52) Scheibel,M.E. and Scheibel,A.B.:The inferior olive.A Golgi study.J.comp.Neurol.,
102:77-132,1955.
53) Scheibel,M.E. Scheibel,A.B., Walberg,F.and Brodal,A.:Areal distribution of axonal
and dendritic patterns in inferior olive.J.comp.Neurol., 106:21-49,1956.
54) Strata,P.:The dual input to the cerebellar cortex.In:M. Santini(ed.)Golgi Centennial
Symposium Proc., pp.273-280.Raven Press,New York,1975.
55) Voogd,J.:The Cerebellum of the Cat.Structure and Fibre Connexions.Thesis.Assen,Van
Gorcum,215 pp.,1964.
56) Voogd,J.:The importance of fiber connections in the comparative anatomy of the
mammalian cerebellum. In:R.Linas(ed.)Neurobiology of Cerebellar Evolution and
Development,p.493-514,Education&Research Foundation,Chicago,Illinois,1969.
57) Voogd,J.and Bigare,F.:The topographical distribution of olivary and corticonuclar
fibers in the cerebellum.A review.In:J. Courville(ed.)The Inferior Olivary Nucleus, Raven
Press,New York,1980.(in press)
58) Walberg,F.and Brodal,A.:The longitudinal zonal pattern in the paramedian lobule of the
cat's cerebellum.An analysis based on a correlation of recent HRP data with results of
studies with other methods.J.comp.Neurol., 187:581-588,1979.
59) Wiklund,L., Bjorklund,A.and Sjolund,B.:The indolaminergic innervation of the inferior
olive.I.Convergence with the direct spinal afferents in the areas projecting to the
cerebellar anterior lobe.Brain Res., 131:1-21,1977.
生体の科学31(2):139-148,1980、医学書院, より許可を得て掲載