
前頭葉の解剖学(Anatomy of the frontal lobe with particular reference to corticocortical connections of the prefrontal area)
--前頭前野の皮質間結合を中心に--
川村光毅
| I.はじめに | |
| II.前頭葉への求心路(図2A) | |
| Ⅲ.前頭葉からの遠心路(図2B) | |
| IV.前頭葉とSTs域との間の相互連絡 | |
| V.ヒトの前頭葉への探訪 | |
| 文献 |
あらためてヒトの前頭葉とはどの範囲かと考えてみると、大脳の中心溝より前方の外套pallium(大脳皮質と大脳髄質に区分される)の部分と定められる。脳回でいえば、大脳半球の外側面にある上前頭回、中前頭回、下前頭回と中心前回、内側面にある内側前頭回、帯状回前部と中心傍小葉前部、それに脳底面にある眼窩回と直回が脳表面に区別される。脳回は脳溝間に生じた隆起につけられた個人差の大きいマクロ的名称である。皮質の細胞構築の差異を基準にして皮質領野をアラビア数字番号記載方式で区分したBrodmann6)の脳地図を借りれば、領野4,6,8,9,10,11,12,24,25,32,33,44、45、46などが含まれることになる。機能的には、一次運動野、補足運動野、前頭眼野、前頭前野、嗅覚野および、辺縁系に属する中古皮質などに分けられる。前頭葉といわれる領域はヒトのみならず、サルやネコなど他の動物にも当然のことながら存在する。そこには、種々の動物に共通の機能領野がみられる反面、動物種により質的にも異なる領域もある。言葉を換えていえば、共通点と相違点と、または、基本的に同じ諭理または基準で語られる部分と、同一基準では論じられない部分が同時に存在する。例をあげると、運動領とか嗅覚野とか前頭眼野などはどちらかというと前者に属し、前頭前野(前連合野)とくに高次中枢神経活動の(複合的)所産といわれる意欲とか情操とか思考などいわゆる高等な精神機能に関係する領域は後者に属する。前頭葉のうちで系統発生的にみて最も新しく、ヒトで最高の発達を示す領野が前頭前野で、細胞構築学的には前頭顆粒皮質(frontal granular cortex)に相当する領域である。ついでにいえば、皮質連合野は後運合野と前連合野に分けられるが、簡潔に言い切ってしまえば、それぞれ、外界からの種々の感覚入カを最終的に分析し旦つ統合処理をして判断する領野である後連合野と、その判断ないし了解にもとづいて外界に対して能動的に働きかける役目をもつ前連合野の領域が、発達段階の差こそあれ、少なくとも哺乳類のレベルでみるとどの動物にも存在する。もう1つ明瞭な点は、動物が高等になるにつれて連合野は領域的に広くなり、とくに前連合野で明らかである。
前頭前野は投射結合系からみると視床の背側内側核(MD)と相互に結合している。皮質間結合の面からみると、同所性および異所性(後連合野へ脳梁線維を送る)の交連線維結合のほかに、連合線維(同側半球間)結合には隣接する脳回を結ぶ短い弓状線維と、側頭極とを結ぶ鈎状束、後連合野中央部とを結ぶ上縦束、それに帯状回内を弧状に走る帯状束などの長い神経路の存在が起始も終止も確定しないまま教科書に記載されている。しかし、ヒトについてはこの肉眼解剖学的所見以上の確かな証拠に基づいた知識を現在われわれは持ち合わせていない。このギャップをネコやサルを用いた実験から得られた知見で埋めることができるだろうか。可とするには少なくとも2つの条件が成立することが必要であろう。すなわち、1)ヒトとサルの大脳皮質の領域区分に関して、かなりの部分で類似性が論じられること、2)サルおよびチンパンジーの皮質間結合を比較した時、その間に少なくとも類似性ないし連続性がみられ、共通する基本点がいくつか抽出できること。さらに述べれば、以上の2点が肯定的に論じられた上で、動物進化の観点から予想される前頭前野の発達にともなう大脳皮質構造および皮質間結合系の質的変化の存在をヒトとサルとの間の相違点として考察されねばならない。現在われわれは、このレペルの論究を進めるに足る段階に近づいているといえよう。
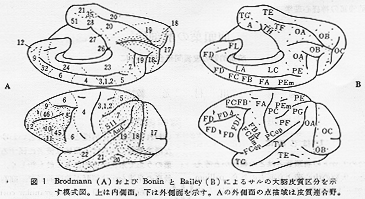
動物間にみられる大脳皮質の領域区分や皮質間の線維結合の類似点と相違点を含めた比較の問題に関しては別に論じているので14)、それを参照していただきたい。ところで、チンパンジーの脳は脳溝の走行をみる限り外見的にはサルよりもヒトの脳によく似ている。しかしながら、この動物の皮質間結合に関する知見は1940年代にストリヒニン法を用いて行なわれたBaileyら1~3)の仕事以降には得られていない。また、ヒトの病的脳や損傷脳を用いて連続切片を作成して線維結合を調べるという研究も進んでいない。このような現状において、サルの所見結果からヒトの皮質間結合を推論することが当面の課題となる。以下の項でHRP法を用いて調べた最近のわれわれの所見4,17)を含めて現時点で明らかにされているサルの前頭前野およびその近傍にみられる連合線維結合の知見について述べる。図1はサルの大脳皮質区分を説明の便宜のために示したものである。
サルの前頭前野はBonin and Bailey5)のFD野、あるいは、Brodmann6)の領野8~12を含み、機能的には遅延反応や弁別学習、行動のプログラミング、短期記憶などに関連する領域といわれている。また、Walker20)はBrodmann6)が領域9、10と名づけた主溝(sulcus principalis)の背側および腹側に位置するかなり広い部分を領域46、それよりもさらに後腹方で弓状溝(sulcus arcuatus)の前方域を領野45としている。ヒトの領野45およびその後方部に発達している領野44は下前頭回後部にあり運動性言語機能に関連する領域(Broca)として知られているが、一般にサルには運動性言語中枢は存在しないと考えられているので、Walkerの領野番号をヒトとの類似で考えることは早計であろう。チンパンジーの言語機能関連域に関する研究結果が望まれる所以である。
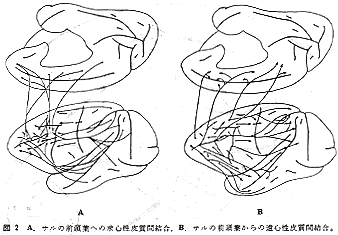
前頭前野への求心路についてであるが、HRP法を用いたわれわれの仕事17)によれば、隣接する領野6(FC,FB)からのほか、PG,OA,OB,TA,TEなど広範囲の後連合野から連合線維を受ける。しかしながら、運動領やいわゆる第1感覚野(SI,AI,VI域)から直接線維を受けない。すなわち、頭頂連合野に属するPG(領野7)からは弓状溝とその付近の領野6,8に多く投射するが、一部主溝の背側部(領野9)および腹側部(領野10)にも線維を送る。また、後頭連合野(OA,OB)からは弓状溝の前方域にある前頭眼野(領野8)に投射があるが、それより前方部の領域9,10にはほとんど投射が認められない。一方、側頭連合野からの投射についてみると、上側頭回(領野22,TA)から主溝腹側部(領野10)への強い投射があり注目されるが、そのほかにも、上側頭回から主溝背側部(領野9)へ、中および下側頭回(領野21,20,TE)から主溝腹側都へ投射が認められる。このように前頭前野への皮質間求心性投射には、かなりorganizeされた系が存在している。この点は後にも述べる(“前頭葉とSTs域との間の相互連絡”を参照)。一般に前頭前野への皮質求心路の集中性は主溝腹側部に強いという特徴がみられる。一方、前頭葉極(領野12)は前頭前野内の領野9、10からの線維を受けるが、側頭極(TG)からの線維もかなり存在する。最後に帯状回からの投射であるが、主溝およびその背側域への強い投射が帯状回全域から認められる。領野8は主として帯状回の後部(領野23,LC)からの線維を受ける。側頭極から強い投射を受ける主溝の腹側域への帯状回からの投射が比較的に弱いものであることは、主溝を境にその背、腹域間に皮質間結合上明らかな差違が存在することを示すものであり興味深い。
簡潔にまとめてみると、前頭前野からの遠心路は、1)頭頂連合野(主として領野7,PG)、2)側頭葉前方域(主としてTG)3)中古皮質(帯状回と帯状溝)および4)後連合野内の2つの主要溝周囲皮質(後出)へと大別される。PG野への起始細胞はFD野のうちでも弓状溝近傍域(領野6,8)に多い。主溝隣接域を含めて、その背側部分はTA野と、腹側部分はTG,TE野との相互結合関係が認められる。特徴的なことは、これらの連合線維群がサルの主要溝である頭頂間溝(intraparietal sulcus)、上側頭溝(superior temporal sulcus, STs)および帯状溝(cingulate sulcus)をかこむ壁面皮質部分に数多く終止することである。なおこのほか、前頭前野はその広範囲から領野6(FB,FC)に弓状線維を送るが、体性感覚運動野(領野1,2,3,4)には送らない。また、一次運動野(領野4)から後連合野に直接投射するものはない。これに反して、帯状回への投射はかなり強力で、前頭前野の広い領域--少なくとも主溝およびその背側に位置する皮質全域--から起こり帯状回への全域(領野23と24,LCとLA)に終止する。そのうち、帯状溝の底部および腹側壁皮質部分に終わるものが多い。
ここで、前連合野(前頭前野)と後連合野(頭頂、側頭および後頭の3つの連合野の総称)との間の連合線維結合に焦点をしぼって考察してみたい。
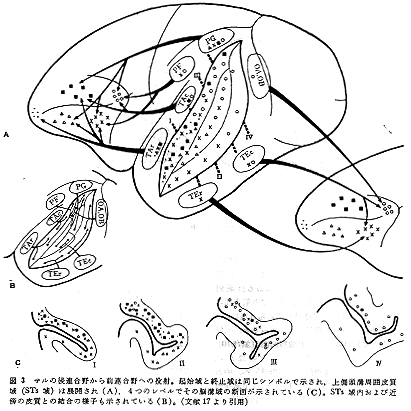
Chavis and Pandya8)はサルの後連合野の外側面皮質に限局した小さな傷を与えて、軸索変性法を用いて前頭葉への皮質間投射についての研究を行なった。そして、彼らが規定した視覚、聴覚、体性感覚系のそれぞれ第1、第2、第3の感覚連合野(sensory association areas: VA 1,2,3;AA 1,2,3;SA 1,2,3)から起こり前頭前野に終わる連合線維の領域に一定のパターンが認められることを示した(彼等の論文8)のFigs.6と7をみよ)。それによると第1連合野(VA 1,AA 1,SA 1)からの連合線維の終止域は弓状溝の前、後付近の領域で部位的差は認められるものの、おのおのが互いにかなり重複する。第2連合野(VA 2,AA 2,SA 2)からの終止域は、主溝腹側部内での重複が顕著である。これに対して、第3連合野(VA 3, AA 3, SA 3)からの終止域はFD野内の一定部位にそれぞれ限局し互いに重複しない。このような特徴ある投射パターンのもつ意味を解釈することは現在困難であるにしても注目に価するように思われる。
厳しい見方をすれば、後連合野を上記のように第1、第2、第3の感覚亜領域に分けるという彼らの着想は良いとしても、その仕方には形態学的立場からみても同意しかねる要素があり多分に便宜的な所があるように思われる。ところで、Jones and Powell 11)が明らかにしたように、後連合野内の上側頭溝(STs)周辺域、とくにその後部の周囲皮質域は、体性感覚性、視覚性、聴覚性など種々の感覚刺激がいくつかのニューロンを介して連合線維によって伝達(播)され一定のパターンを示しながら集中して到達する部位である10,11)。この領域(STs域)には多感覚性ニューロンが多く存在する。すなわち、Bruceら7)はSTs域の脳溝底部および背側壁部分で単一ニューロンの活動を調べた結果、ニューロンの半数以上が多感覚性(polysensory)であり、その性質は、受容野が大きく、動きや形(form)に敏感に反応し、パターン認識よりも空間定位などの機能に関連すると報告した。大脳皮質内の位置的関係、線維結合の状況、機能的な考察などを行ない、進化論的視点に立ってサルのSTs域およびその近傍域に相当する部位をヒトの脳において求めるとすれば、失語、失行、失認の症候出現に関連する部位と考えられている角回(gyrus angularis,領野39)および縁上回(gyrus supramarginalis, 領野40)付近の領域とみなすことができる。しかし、このように結論的な記述のみをすると誤解が生じるので、読者は別にやや詳しく見解をまとめた拙論12~15)を読んでその是非について判断していただきたい。
HRP法を用いて後連合野から前頭葉(特に前頭前野)への皮質間投射について詳しく調べてみると17)、連合線維の起始細胞は脳回面(gyral surface)に存在するばかりでなく、脳溝をかこむ皮質部分(sulcal walls and fundus)にも多数存在し、とくにSTs域に多いことが判明した。そして、この投射系の起始域と終止域の関係を調べてみると、そこにはかなりの程度の部位局在の傾向が認められる。すなわち、STs域の尾側半からは領野8へ、吻側半背側部からは領野9へ、吻側半腹側部からは領野10へと比較的に強い投射がみられる。さらに注目したい点は、後連合野内の種種の部位で前連合野に直接投射する領域は、STs域を介して、つまり、そこで一旦ニューロンを変えて二次的に投射する領域と一致することである。その関係を図3に示してある。この点は、研究がさらに進んだ段階で誤りの少ない考察をしなげればならないと考えている。ともあれ、後連合野から前連合野への投射系に部位局在が存在することの意昧づけは、現段階では詳細なデータが集積されていないためむずかしいが、今後重要な問題として議論されるようになると思われる。
このように、後連合野から前頭前野への投射は広範囲で、且つかなり強い部位局在が認められるが16,17)、逆方向の前頭前野から後連合野への投射をみると、その起始域は主溝および弓状溝の周囲皮質や主溝背側部位(領野9)および前頭眼野(領野8)の一部などに比較的限局しており、且つ、その投射も比較的に弱いものものであることが最近判明した4)。
前頭葉の細胞構築学、組織学、運動(およびその関運)野、嗅覚野、間脳領域および辺縁系との関連、意識や言語の問題、発生学的な問題など重要な問題についてここでは概観することさえできなかったが、それらについては他書9,19,21)および、本特集号の他の論文が大いに参考になるであろう。
これらの諸問題のうち、筆者にとっては、ヒトの症例に接する機会を得させていただくよう努力しながら、前頭葉の機能とその傷害の問題を考察していくことが興味ある課題である。
前頭葉の働きを大胆に表現すれば、“低次’’から”高次”までを含む発展段階に相応した”運動関連系”の範疇に属するものの総体であると言ってもよいであろう。つまり、病的機転としては、運動麻痺も言語障害も自発性減退もすべて能動性機能の低下または障害として把えられないだろうか。
基礎的理解として、前頭葉の構造上および機能的な部位局在が、これまた、質的に程度の差こそあれ、明らかに存在することは容認されよう。Pavlov18)の大脳皮質に関する動的モザイク説が今やCT (computed tomography), rCBF (regional cerebral blood flow), PET (positron emission tomography), NMR (nuclear magnetic resonance)などの近代的武器を使うことによってヒトの脳を対象にして日常臨床の場で検証される時代入っているような気がする。解釈の困難な場合には、それが重要なものである時にはとくに、そのまま問題を大切に残しておくようにし、結論を急いで脳を冒涜しないように心がけたい。
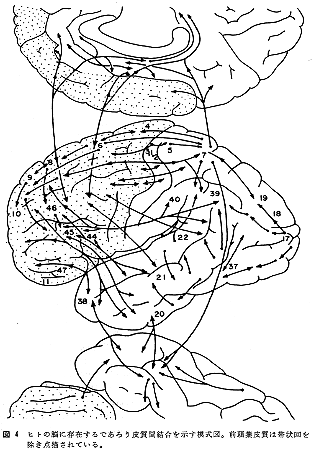
図4にヒトの脳に存在すると思われる連合神経路の摸式図を掲げたが、これは現時点での研究成果の上に立って注意深く推量したものである(以前に発表したもの14)をかなり修正した)。連合線維が離断されたときに現われる機能障害(ないし症状)は、それらの起始部位、障害部位、終止部位のすくなくとも3つの脳部位に起因すると考えられる機能障害がミックスされ修飾された形で症候(群)として観察されるものであろう。”局在”にしても、理論上でなく実際上の問題として、形態的局在から、機能的局在、さらにはいわゆる症候的局在に至るまでの異なる次元での問題を統括して考察できるレベルにまで脳の研究が進展することを期待するものである。ヒトを含めて高等霊長類の脳の生化学的研究などの論文が学術誌に掲載されるようになった。それにつけても、ヒトの正常脳と病的脳の研究成果が、大脳皮質の分野であと何年先に集大成されるであろうか。節酒節煙の控を守り前頭葉をいたわりながら研究生活を長くつづけたいものである。
1) Bailey P, Barenne JG, Garol HW, et al: Sensory cortex of chimpanzee. J Neurophysiol 3; 469-485, 1940.
2) Bailey P, Bonin Gv, Garol HW, et al: Functional organization of temporal lobe of monkey (Macaca mulatta) and chimpanzee (Pan satyrus). J Neurophysiol 6; 121-128, 1973 a.
3) Bailey P, Bonin Gv, Garol HW, et al: Long association fibers in cerebral hemispheres of monkey and chimpanzee. J Neurophysiol 6; 129-134, 1943 b.
4) 伴 亨、川村光毅:サルの上側頭溝周囲皮質への求心性結合に関するHRP法による研究.解剖学雑誌 59; 350, 1984.
5) Bonin Gv, Bailey P: Neocortex of Macaca mulatta, Univ. Illinois Press, Urbana, 1947.
6) Brodmann K: Vergleichende Localisationslehre der Grosshirnrinde, Barth, Lepzig, 1909.
7) Bruce C, Desimone R, Gross CG: Visual properties of neurons in a polysensory area in superior temporal sulcus of the macaque. J Neurophysiol 46; 369-387, 1981.
8) Chavis DA, Pandya DN: Further observations on corticofrontal connections in the rhesus monkey. Brain Res 117; 369-386, 1976.
9) 岩井栄一:脳学習のメカニズム、朝倉書店、1984.
10) Jones EG: The anatomy of extrageniculostriate visual mechanisms. In: The Neurosciences, Third study program, edited by Schmitt FO, Worden FG, MIT Press, Cambridge, Mass, pp. 215-227, 1974.
11) Jones EG, Powell TPS: An anatomical study of converging sensory pathways within the cerebral cortex of the monkey. Brain 93; 793-820, 1970.
12) Kawamura K: Corticocortical fiber connections of the cat cerebrum. III. The occipital region. Brain Res 51; 41-60, 1973.
13) 川村光毅:大脳皮質の連合線維について.形態学的所見をもとにした皮質機能の考察.岩手医学雑誌 26; 629-639, 1974.
14) 川村光毅:”連合野”の線維連絡(I)皮質間結合 - サルとネコの皮質間結合の比較と”連合野”の発達についての試論.神経進歩 21; 1085-1101, 1977.
15) 川村光毅:角回を中心とする皮質間線維連絡.失語研究 2; 13-19, 1982.
16) 川村光毅、伴 亨、鈴木 薫:頭頂葉の構造.Clinical Neuroscience 3; 18-21, 1985.
17) Kawamura K, Naito J: Corticocortical projections to the prefrontal cortex in the rhesus monkey investigated with horseradish peroxidase techniques. Neurosci Res 1; 89-103, 1984.
18) Pavlov IP: Conditioned reflex, an investigation of the physiological activies of the cerebral cortex. Lecture XIII: The Cortex as a Mosaic of Functions (GV Anrep, transl.), Oxford Univ. Press, 232 pp, 1927.
19) The Frontal Lobes (特集): Trends Neurosci 7/ Nov; 403-454, 1984.
20) Walker AE: A cytoarchitectural study of the prefrontal area of the macaque monkey. J Comp Neur 73; 59-86, 1940.
21) 随意運動のメカニズム(特集):神経進歩 28; 1-178, 1984.