
脳の形態と機能―精神医学に関連して
川村光毅
|
|
|
|||||||
ヒトの脳と脊髄は3種の結合組織性の膜、すなわち硬膜、くも膜、軟膜におおわれている。硬膜は厚い線維性の膜で、中に血管や神経を含んでいる。くも膜は薄い膜で、突起を出して軟膜とつながっている。軟膜は脳表面を直接おおう血管の多い薄い膜である。くも膜と軟膜の間のくも膜下腔は脳脊髄液をいれている。脳脊髄液は、一層の脈絡上皮層と軟膜が接している脳室(側脳室、第三脳室、第四脳室)内の特定の部位から突出した脈絡叢という組織で作られる。このように、脳は液体にひたされた柔らかい器官で三重の膜により保護されている大切な器官である。また、脳硬膜は内外の2層から成り、脳の被膜であると同時に頭蓋骨内面の骨膜である。特定の部位で、2層の間に硬膜静脈洞を作る。脳脊髄液は、くも膜下腔から大脳の天井で静脈洞へ突き出しているくも膜顆粒を通して静脈中へ吸収される。脳の血液はここへ集められて内頸静脈へ流出する。なお、硬膜には弾性線維も含まれており、脳脊髄液の分泌と吸収のバランスで一定の圧力によりテンションを保っている。
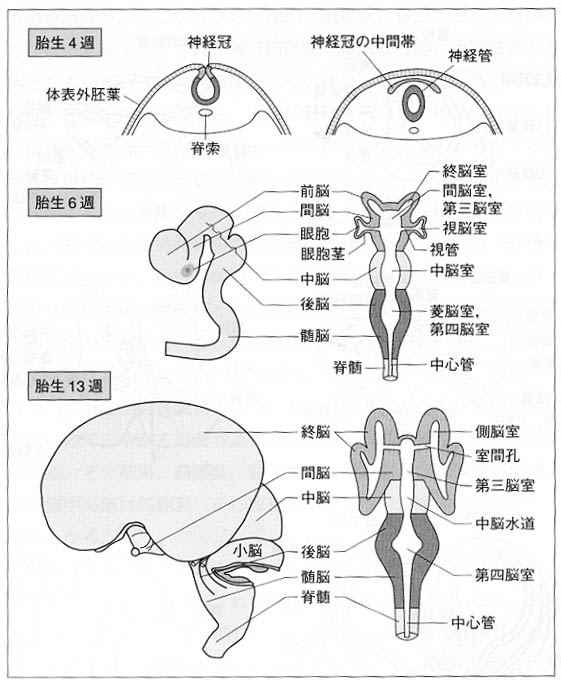
図1:ヒトの中枢神経系の発生を示す模式図。脳は均一に発達するものではないことに留意したい。
最初に、神経系の初期発達について述べる(図1)。受精卵が細胞分裂し、増殖・成長し、内、中、外の3胚葉が形成される(胎生3週)。次いで、外胚葉の表皮が陥入し、4週のはじめに神経板の左右の外側縁が癒着し、神経管が形成される。神経板の外縁部は神経冠(神経堤ともいう)となる。神経冠の細胞は一つは体表外胚葉の直下を、他は胚体内を移動する。こうして最終的に神経管系統からは中枢神経系が、また神経冠系統からは最終到着部位に応じて脊髄神経節、自律神経節後ニューロン、シュワン細胞Schwann
cell、色素細胞が分化してくる。このほか、外胚葉からは、嗅上皮、レンズ、内耳の原基を作る部分が肥厚してプラコードと呼ばれる組織が生じる。
神経管の腹側には、脊索が脊髄管全長と脳管の後半にわたって位置する。脊索が位置しない脳管の前半から終脳と間脳が分化し、脊索が位置する脳管の後半から中脳と菱脳が分化する。脳管は三つの膨らみをもち、前方から前脳胞、中脳胞、菱脳胞と呼ばれる(4-6週)。これらの"膨隆"
ないし"肥厚"は脳室層(神経上皮層)における細胞の増殖・分化および次いで形成される外套層内の細胞移動にほかならず、外套層が腹方と背方へ膨らんだ部位を、それぞれ基板と翼板という(両生類以上に認められる)。前脳胞の前端部の左右から終脳胞が膨れて形成される。残りの部分が間脳胞でその左右から眼胞が突出して網膜が分化する。また、菱脳胞の背側前端部から小脳が分化する。この小脳と橋になる部分を後脳胞、この後方に続いて延髄になる部分を髄脳胞と呼ぶ。この形成過程でヒトの神経管は、脳の部分で二つの大きな屈曲を示す。それらは、脊索の前端から前へ前脳胞が中脳域に続く部位(頭屈)と両側に耳胞、背側に小脳原基が生ずる後脳と髄脳の境界域(橋屈)である。ほかに、菱脳と脊髄との境に屈曲(頸屈)がみられる。
神経系の発達を系統発生的にみると、細胞の突起が互いに連絡する神経網(腔腸動物)、次いで細胞が集まって神経節(環形、軟体、節足動物)を作り、分節化し体軸が定まると梯子状に左右の神経節が連なり、中枢化がすすみ脳神経節が発達する。最後に、脊索動物、脊椎動物の段階で背中の中央を走る神経管が現れる。
ヒトの精神の異常を対象とする精神医学における科学的治療法を問題とする場合、脳の発達をもとにした構成に関する基礎的知識をもつことは大切なことである。とくにヒトで高度の発達を遂げた前脳(のちに終脳と間脳に分化する)の部分は重要である。
前脳が注目される第一の理由は、言語中枢がヒトでこの領域に存在するということ。第二に、ヒトの精神すなわち高次神経活動が認知機構と情動機構に大別されるとして、その主役は前脳にあることである。ヒトの脳はネコやサルの脳と基本的に同じ構成をもっているが、動物の脳と異なる大きな点は、ヒトでは言語を使用できる脳髄をもっていることである。これは、パブロフの条件反射の第二次信号系(信号を信号化するというもっとも抽象的な符号の系)がヒトの段階になってはじめて発達したことを意味する。
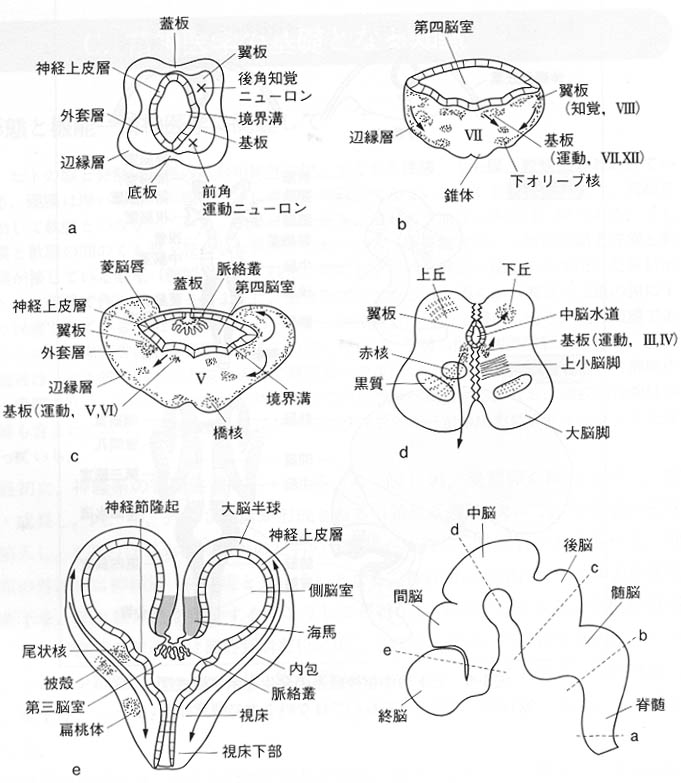
図2:ヒトの中枢神経系の発達が部位別に異なり、それがどのように構成されているのかを学ぶように。基板は運動性細胞を含み、翼板は知覚性細胞を含む。
終脳は前脳胞の前端部の背外側壁が外方、次いで背外方に袋状に膨出することによって発生を始める。胎生2か月の中ごろから半球胞の腹側壁(脳室面神経上皮)においてさかんな細胞増殖が起こる。ここで発生した幼若線条体ニューロンは、まず脳室下層を形成し、そこからさらに深部へ放射状に移動する。この部分は側脳室に向かって丘状に隆起する。これを神経節隆起(基底核隆起、大脳核丘ともいう。のちに線条体となる)という。神経節隆起はその後急速に大きくなり、半球胞の腹側ないし腹外側壁のほぼ全体を形成する。大脳核原基外側端部の神経上皮からは背外側方へ接線方向に移動して新皮質の介在ニューロンに分化するものが発生する。またその内側端部からは扁桃体核や旧皮質に移動し分化するニューロンが発生する。また、一部間脳域に移動するニューロンも存在する。ところで、前脳胞の膨出は前方にも向かい、成長した大脳では本来の神経管の前端部は視交叉上部の薄い膜(終板)に相当し、大脳の前端部ではない。終板の後方では左右の脳室が室間孔で交通し第三脳室に続く。この部分は脈絡組織が脈絡叢として側脳室と第三脳室へ侵入する部位にもあたる。後方への膨出部の内側壁の一部は脈絡組織となり脈絡裂という。
半球胞は発生の進行につれて増大していくが、この際、外套(神経節隆起以外の部分で、大脳の皮質と髄質を作る)および側脳室の拡大が神経節隆起の増大よりもずっと速いので、やがて外套および側脳室が、神経節隆起を前上方と後下方から包むようになる。その結果、前頭葉、頭頂葉、後頭葉および側頭葉が形成される。半球外側面の腹側中央部に外側窩(その底部を島という)と呼ぶ凹みが生じ、それが陥没して外側溝となる。
また、外套の発育の過程でもっとも早く出現する脳溝は鳥距溝・頭頂後頭溝・帯状溝などであり、中心溝がこれに続く。胎生7か月にはいると、さらに中心前溝・中心後溝などの多くの溝が出現し、溝と溝の間の部分は大脳回となる。胎生7か月末になると大脳半球外表面の基本形が完成する。
大脳皮質が形成されるにつれて、大脳皮質から出て視床(第VI層ニューロンから)および脳幹・脊髄(第V層ニューロン)へ向かう下行線維が増加する。これらの線維は外套と神経節隆起の移行部から神経節隆起内に進入し、室間孔の後縁のところで間脳の前端部に進入する。視床経由で大脳皮質に達する上行線維も、この経路を逆行する。この神経節隆起(線条体に分化)を貫いて走る線維群を内包といい、神経節隆起はこれによって側脳室に隆起している背内側部と、その腹外側部に分断される。前者を尾状核、後者を被殻という。内包の発達が悪い下等哺乳類では両者の区別は不明確で尾状核/被殻と呼ばれる。なお、海馬は脈絡裂のすぐ上方の外套壁が肥厚して形成され、側脳室内へ突出する。海馬は本来前脳胞が背方へ膨出した時、その背内側壁に形成され、海馬と乳頭体を結ぶ線維は最短距離である前脳胞前壁を通って走っていたが、その後の大脳新皮質の発達により後方へ、次いで下方へ、さらには再び前下方へと押しやられ、乳頭体への線維束(脳弓)は脳梁の下を大きく弧を描いて走ることとなった。これは大脳新皮質の発達を示す好個の例といえる。また、嗅球は前脳胞の前下端から膨出した構造で、下等動物では嗅球内に側脳室が存在する(嗅球脳室)。
基板と翼板は神経管の側壁の肥厚であるが、これに対して、神経管の蓋板と底板(床板ともいう)では神経組織の増生は起こらない。しかし両側の神経組織の発達に伴い、それらの連合を行ったり左右交叉する線維は神経管の蓋板か底板の領域を通るのでその部位では肥厚する(例をあげると、視交叉、前交連、後交連、結合腕、錐体交叉、毛帯交叉、滑車神経、手綱交連など)。脳梁は大きくなりすぎて前脳胞の蓋板内には収まりきらず大きく後方へ突出する。
このように、終脳は周辺を取り巻く外套と基底部から脳室内へ膨出する神経節隆起からなるが、ともに系統発生を反映してそれぞれ原始archi-、旧paleo-、新neo-に分けられる。外套についていえば脳室内側壁の背側には原始皮質である歯状回と海馬が生じ、神経節隆起の腹外側には旧皮質である梨状葉皮質が生じる(胎生7週)。これらには本来はそれぞれ内側・外側嗅束がはいり、嗅覚の中枢として中隔や手綱や視床下部に線維を送っているものである。哺乳類では脳室背側の新皮質が大きく発達して高次神経機能を担うようになり、これら古い皮質は本来の嗅覚の中枢が発展して、情動に関係する領域となる。大脳基底核では原始線条体は扁桃核へ旧線条体は淡蒼球へ新線条体は尾状核/被殻へと発展して錐体外路の主要中枢となる。ちなみに鳥類は皮質の発達ではなく、新線条体より上位へ上位線条体hyperstriatumという新しい高次中枢を発達させた動物である。
新皮質の中に、第1感覚野(体性感覚-、視覚-、聴覚-、味覚野など)、運動野に続いいて連合野が形成される。ついで、ヒトで集団的労働の社会が形成されるに従って、連合野内に言語野が発達する。
ヒトを含む哺乳類の大脳皮質は領域により、特有の細胞配列パターンを示し、基本的に6層の構造をもち、深層から表層へと順に形成される。表層からの名称は I
分子層、 II 外顆粒層、 III 外錐体層、 IV 内顆粒層、 V
内錐体層、 VI
多形細胞層 である。このタイプの大脳皮質の形成は動物が高等になり、進化のすすんだ段階で獲得した様式で以下の①~⑤の順にすすむ特殊化した組織形成過程を示している。すなわち、①脳室面神経上皮細胞が増殖・分化する。②表層に向かって放射線状に移動する。③プレプレートpreplateを最初に形成する。④次に発生・移動したニューロンはプレプレートの間に割りこんで、深層から表層への順に皮質板を形成する。⑤遅れてグリア(神経膠細胞)が発生する。このほか、接線方向に移動する細胞群の存在も知られている。これらは介在ニューロン(GABAニューロン)に分化して各層に広く散在性に分布する。
胎生4週末の前脳胞ではその外側壁の前腹側端部から外方に向かって眼胞が膨れる。最初は眼胞の内腔(のちの色素上皮と視細胞外節との間)は神経管腔と交通している。5週にはいると、その基部付近の外側壁が前・外・背方に向かって膨れ1対の半球胞を形成する。左右の半球胞を連ねる前脳胞の前背側端部を終脳正中部という。終脳正中部と半球胞とが終脳を形成し、残りの大部分を占める領域は、間脳胞と呼ばれるようになる。このように間脳は主として間脳胞の左右両側壁の肥厚によって形成される。
肥厚・増大し、モザイク状に分化していく間脳の第三脳室壁の内面には一時期、前後に走る三つの溝(間脳背側溝、間脳中間溝、間脳腹側溝)が認められ、これによって間脳は視床上部、背側視床、腹側視床、視床下部の4部に分けられる。
視床ははじめ翼板の前方域、次いで胎生2か月末には翼板全域の細胞が増殖・肥厚して生じ、大きな灰白質の塊となる。視床の核域は明瞭な2群に分かれる。すなわち背側核群と腹側核群である。翼板の後部からは視床後部が生ずる。これは内側膝状体(聴覚神経路の中継核)と外側膝状体(視覚神経路の中継核)という2個の細胞集団に分化する。なお翼板の背側部および蓋板の一部から視床上部が生ずる。視床上部は、ヒト脳では発達が弱く視床の内側の後上端部に位置し、手綱および手綱三角と松果体が区別される。松果体は内分泌(神経分泌)器官で、腺部と感覚部があり哺乳動物の松果体は腺部が発達したもので日周リズムや性成熟に関わるホルモン(メラトニン)を分泌する。爬虫類以下の動物では感覚部に網膜に似た構造を有し光感受性がある。これらの動物ではメラトニンは体色変化を司る。
視床下部は間脳の外側壁腹側部(いわゆる基板と底板の領域であるが、一部翼板も含む)の肥厚によって形成され、いくつかの独立した核に分化する。これらの領域は食欲、睡眠、性欲、体温および情動行動のような内臓機能調節中枢として働く。視床下部の前端をなす視陥凹のすぐ後下方に続く底板は、胎生5週ごろから肥厚し始め、視交叉板となる。網膜からの神経軸索がここに到達し、視交叉を形成する。視交叉板の後方に続く間脳底は円錐形となって腹側に突出する。これが漏斗でその先端部は下垂体後葉となる。間脳底の後端部は正中線上に1個の半球形の高まりを生ずる。これはやがて左右の2個に分かれ乳頭体となる。漏斗の基部は肥厚して灰白隆起(正中隆起)となる。この部位は毛細血管に富み、ここには視床下部諸核からの各種放出ホルモンが運ばれて終わる(後出)。
中脳胞(菱脳胞との境は胎生4週で明瞭となる)の背外側壁をなす翼板は背外方に増大し、四丘板を形成する。その後、四丘板は4個の半球状のたかまりに分割され、上丘と下丘となる。
上丘では神経上皮層で生じた神経芽細胞は表面に向かって放射状に移動し、表面に平行な三つの層を形成する。これらの細胞層の間に上丘に出入りする神経線維の層ができる。下丘では神経芽細胞は下丘の内部を埋め、全体として単一の下丘核を形成する。上丘の核は視覚刺激の連合および反射中枢として働き、下丘の核は聴覚刺激の中継所として働く。四丘体の形成のほかに、基板から動眼神経核、滑車神経核、エーディンガー・ウェストファルEdinger-Westphal 核, ダークシェーヴィチDarkschewitch
核が生じる。また、翼板と基板(主に後者)の細胞が移動して赤核および黒質が形成される。
中脳後部の被蓋では上小脳脚の大量の線維が交叉する(結合腕交叉)。中脳の腹側では大脳から下行する線維が太い束になって存在し大脳脚といわれる。左右の大脳脚の間は深い谷間になっていて脚間窩と呼ばれる。
菱脳の頭側部半の後脳では、翼板の背側部で蓋板との移行部を占める菱脳唇から小脳が形成され、翼板の腹内側部と基板から橋が形成される。橋核ニューロンや小脳の顆粒細胞は軟膜下を最終定着地に向かって移動する。後脳の菱脳唇は多数の神経細胞を生ずるが、これらは、辺縁帯の中を腹内方に移動し、基板の辺縁層の中に大きい神経核を作る。胎生3か月ごろ、頭側部から生じた神経細胞は後脳の腹側部に橋核を、尾側部から生じたものは髄脳の腹側部に下オリーブ核を形成する。下オリーブ核ニューロンは軟膜下または実質内を移動する。橋被蓋は、後脳の翼板と基板から形成され、菱形窩の頭側半をなし、ここに三叉、外転、顔面および内耳神経の諸核が生ずる。外套層の大部分は上行、下行、横走の諸線維が走行し、線維間に散在する翼板および基板由来の神経細胞とともに網様体を形成する。
網様体は上部頸髄において灰白質と白質の間に、線維がおりなす網の目の間に大小のニューロンが散在する構造として生じ、上方へ延髄―橋―中脳―視床まで連続する。この構造は神経束や神経核が分化する以前の原始的中枢の形態を示すものと考えられる。被蓋正中線をはさんで縦に長い線維の束が走るが、これは内側縦束と呼ばれ、中脳被蓋に発し脊髄まで続く。これは中脳以下で行われる姿勢反射をはじめさまざまな運動性反射の回路を形成する線維の集合で、系統発生的に古く、ヒトの発生では最初に髄鞘をとる。蓋板は上部には小脳が発達するが、下部では薄いまま残存し第四脳室脈絡叢を形成するほか、外側と正中に孔(外側孔と正中孔)が開きここから脳脊髄液がくも膜下腔へ流れでる。
4か月になると、橋核には大脳皮質から下行し、橋縦束の一部を構成する神経線維が終わる。ちなみに、橋縦束は大脳皮質から出て橋および延髄の脳神経運動核と脳幹網様体ならびに脊髄前角細胞にいたる錐体路線維と、直接橋核に終わる皮質橋路線維とからなっている。
小脳は後脳の翼板の背外側が背内方に突出する菱脳唇から発生する。胎生2か月後半に急速に増大し、左右の菱脳唇が正中部で合して小脳板となる。正中部からは小脳虫部が、外側部からは左右の小脳半球が形成される。
小脳原基の背側面には、虫部から半球に向かって走る溝が出現する。胎生5か月末には小脳虫部の主な区分が完成する。小脳発生の最初に現れる溝は系統発生的にもっとも旧い小脳部分である小節と、それに次ぐ虫部垂
の間の溝(後外側裂)で、次いで第1裂が生じる。これらの構造は上方から新しい領域が生じてくるにつれて後方へと押しやられる。
小脳原基においても第四脳室に接する内側から表面に向かって神経上皮層・外套層・辺縁層の3層が分化する。この神経上皮層から小脳核の神経細胞、小脳皮質のプルキンエPurkinje細胞、ゴルジGolgi細胞が発生する。小脳原基が3層に分化するとまもなく、放射状グリアに沿って表層に向かって移動した大型の幼若神経細胞が小脳板の背側面(表面)に平行に1列に並ぶ。これがプルキンエ細胞の幼若型である。次いで小脳板の尾側端部の神経上皮層でさかんな細胞分裂が起こり、ここで生じた未分化細胞は辺縁層の軟膜下の表層部を前内方に移動して、小脳原基の全表面をおおう未分化細胞層を形成する。これを胎生外顆粒層という。この層の細胞は移動中にも分裂し、神経細胞を作りだす。この神経細胞は辺縁層およびプルキンエ細胞層を貫いて、プルキンエ細胞の層の下に達し、ここに新たに内顆粒層を作る。胎生顆粒層からは、このほかに辺縁層の中に散在する篭細胞や小皮質細胞が生ずる。このように小脳を構成するニューロンの発生も複雑でかなり特殊化されたものである。かくして、小脳の全表面は、表面から分子層・プルキンエ細胞層・(内)顆粒層の3層から成る小脳皮質でおおわれる。
小脳求心線維は下オリーブ核からの登上線維と橋核などからの苔状線維があり、おのおのプルキンエ細胞、顆粒細胞に終わる。小脳遠心線維はほとんどが小脳核から起こり、小脳核はプルキンエ細胞からの入力を受ける。
脊髄の発生は神経管の発生そのもので、もっとも原始的な基本的な型が保たれている。神経管の左右の壁は活発な細胞分裂によって肥厚し、内側から外側に向かって同心円状に神経上皮層、外套層、辺縁層の3層に分化する。左右の壁のうち基板は翼板よりも早期に発育し、その外套層は神経細胞の集積によって腹外方に拡大し、全体として前角を形成する。背側半はやや遅れて発育し、その外套層は背外方に拡大して後角となる。基板の最背側部から生じた神経細胞は、前角の背外側部に集まって側角を形成する。後角の神経細胞は、脊髄神経節からはいってくる後根線維に接続して、これによってもたらされる求心性刺激を、他の脊髄レベルおよび脳の各部に伝達する。
後根線維のうちで高等な触覚を伝える長後根線維は、脊髄にはいると上行して延髄の後索核(薄束核と楔状束核)にいたる。後索核ニューロンの軸索の束は毛帯交叉をしたのち、内側毛帯と呼ばれ、対側の視床後腹側核にいたる。神経管の蓋板と底板は神経上皮層と辺縁帯のみが分化するが、翼板と基板の発達に伴い左右を連絡する線維や交叉する線維は蓋板あるいは底板を通るほかないので、それらの線維の増加とともに厚みを増す。脊髄視床路の起始細胞は後角ニューロンであるが、その軸索は脊髄中心管の腹側域にある底板を通過して対側に向かう。
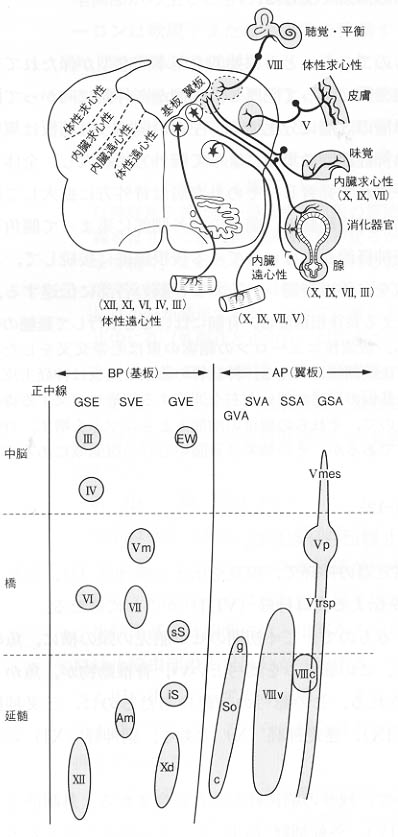
図3:脳管(発達した神経管の脳の部分)、とくに、延髄の基本構造を示す断面図の模式図。脳神経の機能的要素には、遠心性(運動性)と求心性(感覚性)、一般性と特殊性があり、胎生期にそれぞれの性質をもった柱状の細胞集団、すなわち、細胞柱が区別される。GSE:一般体性遠心性,
SVE:特殊内臓遠心性, GVE:一般内臓遠心性, GVA:一般内臓求心性, SVA:特殊内臓求心性,
SSA:特殊体性求心性, Ⅲ:動眼神経核, Ⅳ:滑車神経核, Ⅴm:三叉神経運動核,
Ⅴmes:三叉神経中脳路核, Ⅴp:三叉神経主知覚核, Ⅴtrsp:三叉神経脊髄路核,
Ⅵ:外転神経核, Ⅷc:蝸牛神経核, Ⅷv:前庭神経核, ⅩⅡ:舌下神経核,
Ⅹd:迷走神経背側核, Am:疑核, c:循環呼吸部分(孤束核),EW:エディンガー・ウェストファル核,
g:味覚部分(孤束核), iS:下唾液核, So:孤束核, sS:上唾液核
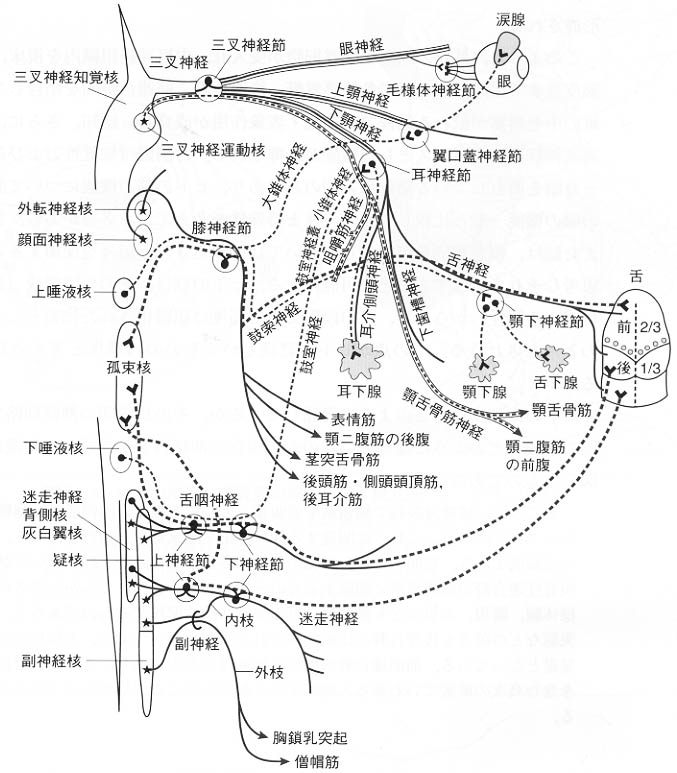
図4:脳神経の構成を示す模式図。一見複雑にみえるが、重点を考えて簡略化してある。
脳神経は、由来や働きによって、3群に分けられる。
第1群は、頭にだけある特殊な感覚器の神経で、嗅覚を伝える嗅神経(Ⅰ)、視覚を伝える視神経(Ⅱ)、聴覚と平衡覚を伝える内耳神経(Ⅷ)がこれにあたる。
第2群は、魚の鰓弓の神経にあたるもので、ごく初期のヒト胎児の頸の横に、魚の鰓に似た切れ込みと膨らみがみえる。この膨らみを鰓弓といい、脊椎動物が、魚から四足動物に進化した名残りと考えられる。この鰓弓の神経にあたるのは、三叉神経(Ⅴ)、顔面神経(Ⅶ)、舌咽神経(Ⅸ)、迷走神経(Ⅹ)である。副神経(ⅩⅠ)の一部も鰓弓神経に含まれる。
第3群は、脊髄神経にあたるもので、残りの脳神経がこれに含まれる。外眼筋を支配する動眼神経(Ⅲ)、滑車神経(Ⅳ)、外転神経(Ⅵ)と、舌の筋を支配する舌下神経(ⅩⅡ)である。
なお、本来の脳の最前端である終板基底部から1対の神経が鼻腔へいっており昔はこれを第1脳神経とした。ヒトでは発生の過程で退行するがこれを有している動物は多い。
脳神経核の本来的位置は体性運動性と体性知覚性のものはそれぞれ境界溝から遠い基板と翼板に位置し、神経は神経管の腹側と背側から出入りする。内臓性運動性と内臓性知覚性のものは境界溝に近く位置し、その神経は側面から出入りする。鰓弓は咽頭壁に形成される呼吸器であるからその核と神経は内臓性の位置をとる。
脳内の脳神経の発生学的構成と完成された脳幹内におけるそれらの位置(部位)については図3に、また、脳外における脳神経の走行と名称については図4に、おのおの模式図として示されている。全体をまとめる時に参考にして欲しい。
われわれの身体の内外に生起するさまざまの変化は、特殊に発達した有毛感覚細胞によって受け取られる。皮膚、舌、鼻、目、耳などの感覚器官内にこれらの細胞はみられる。「五感」といわれるこれらの触、味、嗅、視、聴の感覚をひきおこす物質の性質は異なるが、興奮はすべて中枢神経系(脳と脊髄)にはいり、それぞれの認知機構を有し、その結果、有機体の能動系(筋や腺)に伝達されるという共通点をもっている。また、体性感覚についていえば、表在感覚と深部感覚があり、深部感覚は内臓感覚同様、性質と位置の認識は曖昧であるが情動の基盤として重要な意味をもっている。
①
体性感覚:皮膚・粘膜 → 脊髄後角 → 視床後腹側核→皮質体性感覚野
延髄後索核
② 味覚:味蕾→延髄孤束核→視床後腹内側核→皮質味覚野
③
嗅覚:鼻粘膜嗅細胞→扁桃核→視床背内側核(大細胞群)→皮質嗅覚野
④
視覚:網膜視細胞→双極細胞→網膜神経節細胞→外側膝状体→皮質視覚野
⑤
聴覚:内耳有毛細胞→蝸牛神経核→外側毛帯核→下丘→内側膝状体→皮質聴覚野
⑥
他に明瞭な意識にはのぼらない内臓感覚や固有感覚(筋紡錘、腱紡錘、関節包な ど)がある。
①~⑤の上行性感覚伝導路の最終領域である皮質感覚野は第一感覚野といわれ、連合線維すなわち皮質・皮質間線維により、第二、第三感覚野(すなわち後連合野)に刺激が伝達される。
皮質連合野は前連合野(前頭前野)と後連合野(頭頂、側頭、後頭の各連合野)に分けられる。言語を使用する動物、ヒト、において、後連合野の内に感覚性言語野(ウェルニッケ
Wernicke 野)が、前連合野内に運動性言語野(ブローカ Broca
野)が形成される。
このように、外界の刺激を感覚細胞が受け入れ、中枢神経組織内を視床、さらに、大脳皮質まで伝達し、そこで各種感覚野、後連合野、前連合野間を結合している連合線維の中を興奮が伝わる過程で、知覚・表象作用が成立し、認知、さらに、認識という高次神経活動をベースとした機能が発揮される。言語野(知覚性および運動性)をもった脳を所有している動物は人間のみであり、ヒトの脳の機能について語る際、動物の脳の機能一般と比較して、同一性と特殊性を考えておく必要がある。言語機能を備えた脳は、概念形成を可能にし、ひいては抽象化された信号を使用することにより、思考しそれを伝達することを可能にする。ヒトの脳は、その化学構成(分子的化合を形成する属性)からみて、この段階にまで高度に組織化された物質として発展したものとみなされうる。この点がヒトの意識というものの特殊性と考えられる所以である。
ヒトの大脳皮質がどのように発達してくるか、その皮質間の神経回路である連合線維の構成はどのようになっているのか。現在の神経科学が到達した知識に基づいて作図したものである(図5)。
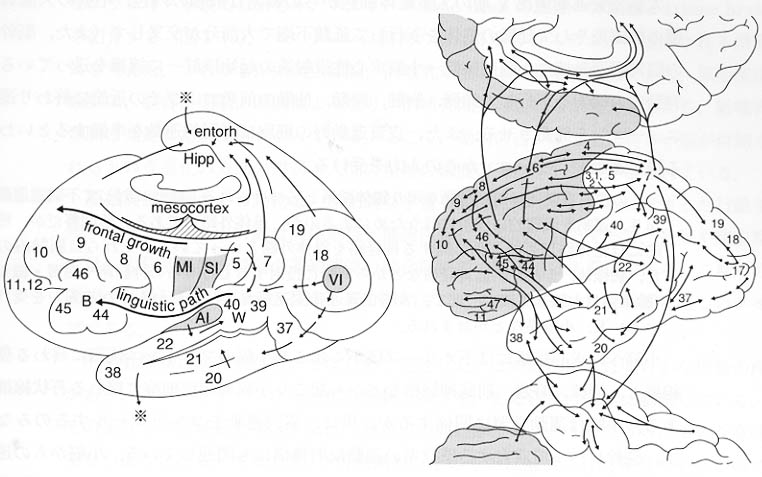
図5:ヒトの皮質・皮質間の構成を示す模式図(右:川村、1988、
より)と皮質の段階的発達を示す想像図(左:川村、1977、より)。アラビア数字は皮質区分を示すBrodmannの番号。B:
Broca 言語野、 W: Wernicke 言語野、AI, SI, VI
はそれぞれ第一聴覚野、第一体性知覚野、第一視覚野を表す。*で連続している。
ところで、精神分裂病で側頭葉や前頭葉、とくにそれらの内側部の細胞構築等の障害例が示されているが、これらに関連する領野は第一感覚野や運動野ではなく、連合野に相当する領域である。想像するに、幻聴や幻視は多少とも言語野と結びついた聴覚性連合野や視覚性連合野の機能異常に関係するものと思われる。また、てんかん患者にみられる夢幻様体験、既視,未視なども側頭連合野の異常活動に関係するものであろう。失語、失行、失認などの障害も後連合野の比較的限られた領野の障害であり、大脳病理学が局所診断の基礎となっている。前頭連合野の機能は能動性の発現であり、その異常は自閉、保続などを含む高次の障害でいわゆる人間味を失う起因となる責任病巣がここにあるように思われる。
筋系に興奮を伝達し筋収縮をひきおこすのは脊髄前角細胞であるが、筋収縮が拮抗筋や共同筋との相互関係の中で調和のとれた収縮を行い、もっとも効果的かつ合目的的な関節運動を行うためにさまざまな調整機能が存在する。
1個の運動ニューロンは多数の(ときには100個をこす)筋細胞を支配しており、これを神経筋単位という。また骨格筋には速筋と遅筋があり、前者が主に無酸素的に瞬発的収縮を行うのに対し、後者はクレブスKrebs回路の好酸素的解糖系に由来するエネルギーによって収縮する。ヒトの筋にはすべて速筋と遅筋が混在しており筋運動の要求に応じて使い分けされている。
運動性反射経路は、脊髄レベル、延髄・橋レベル、中脳レベル、より高次レベルでの反射回路を通って脊髄前角へ到達する。大脳皮質運動領の錐体細胞からの運動ニューロンへの入力は意識的運動を誘発するが、そこへは錐体外路系をはじめ無数の反射経路からの信号が、ある場合にはそれを促通し、ある場合には抑制することによって神経筋単位の活動を調整している。
大脳皮質運動領第V層の大型錐体細胞からの軸索は前脳の内包、中脳の大脳脚、橋の橋縦束そして延髄の錐体を下行して延髄下端で大部分が交叉して(また、脳幹内の脳神経運動核や小脳前核群―小脳求心性投射系の起始核群―に線維を送っている)、脊髄内の側索を下行して頸髄、胸髄、腰髄、仙髄の前角およびその近傍に終わり運動ニューロンを興奮させる。また、皮質運動野の細胞は意図的運動を準備するといわれる補足運動野ニューロンからの入力を受ける。
この皮質脊髄路は随意運動を司り錐体路系と名づけられる。これに対して不随意運動を司り、運動を協調的に円滑に行うためにあるのが、錐体外路系である。珍問答だが、形態的には錐体路以外の運動に関する神経路を錐体外路といっている。それらは脳幹内の上丘、網様体、赤核、前庭神経核などから脊髄に投射するもろもろの神経路や黒質・線条体(線条体は大脳基底核の一部で、大脳皮質運動関連野および黒質から主たる入力を受けている)系、小脳系などが含まれる。
小脳への求心線維には下オリーブ核から起こり小脳のプルキンエ細胞に終わる登上線維と、橋核、脊髄、前庭神経核などから起こり小脳の顆粒細胞に終わる苔状線維がある。小脳は運動学習に関係するが、単に、協同運動をコントロールするのみならず、脳幹レベルで成立する感覚系の運動反射機構にも関連している。小脳からの遠心線維は脳幹内の前庭神経核、下オリーブ核、赤核、視床運動核などにいたり、視床経由の神経路を介して大脳運動野に達してそこに興奮を起こさせる。このように、小脳と大脳との間には相互関連ループが存在している。
このほか、反射機構も重要で、運動性反射は関節運動だけでなく関連痛とも関係して筋の固縮などをひきおこす。また、自律系反射は内臓・血管運動、分泌、内分泌に重要な変化をもたらす。
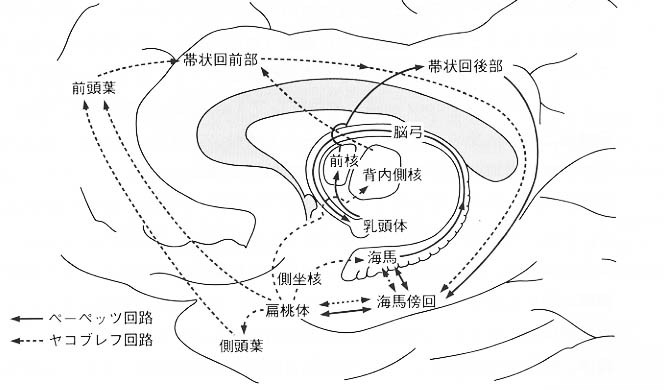
図6:情動機構を説明するうえでのキーワードとなる構造物とそれらを連結する神経回路(Papez
回路、Yakovlev 回路)を示す模式図。
扁桃体は、快・不快などの感覚刺激の生物学的価値評価および条件づけの獲得や情動体験によるエピソード記憶の固定過程(短期記憶から長期記憶への移行)に関与している。また、大脳辺縁系が関与する情動および/または記憶の回路として有名なものに、ぺーペッツ回路とヤコブレフYakovlev回路がある(図6)。現在、辺縁系は脳の情動系を構成しているとする情動の辺縁系説が有力であるが、海馬や扁桃体や嗅脳溝周囲皮質を含む側頭葉辺縁領域は情動とともに少なくとも記憶のような認識機構にも関与していることが示されている。ちなみに意欲の研究は遅れているが、最近、行動生理学的研究の結果からも帯状回がこれに関与していると考えられるようになった。神経解剖学的には、扁桃体には脳幹レベルから味覚や内臓知覚、血液循環に関する諸核(孤束核、結合腕傍核など)から直接の投射があり、また、聴覚入力は視床レベルから(内側膝状体の関連近傍核から)の入力もみられる。さらに、扁桃体は前頭葉や側頭葉との間に相互結合関係をもっている。また、扁桃体中心核からの出力経路の一つに、視床下部や中隔核へ向かう神経束(分界条)がある。
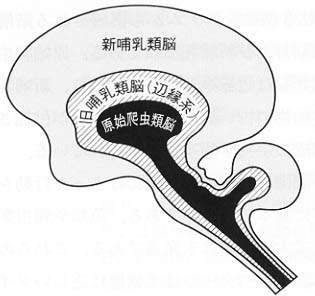
図7:マクリーンの三位一体脳説といわれる一見神秘的色彩を帯びた説で、進化する脳の階層性を示す模式図。情動、認知、思考など心理学でも使用される用語を脳の構造と機能と結びつけて理解するうえに役立つであろう古典的なモデル。
ここで、系統発生学的に情動機構をどのように眺めるかという観点から、古典的なヒエラルキー学説ではあるが、マクリーンMacLeanの三位一体脳説a
triune brainを紹介する(図7)。マクリーンは動物の脳に3型のシステムから構成される階層性を考えた。いわゆる原始爬虫類脳、旧哺乳類脳および新哺乳類脳である。原始爬虫類脳は、脳幹、間脳、基底核よりなり、旧哺乳類脳は辺縁系が強力に加わり、新哺乳類脳は新皮質をもっている。ここで、動物の古い脳の上に新しい脳が付加されるという進化の方向の道筋と人間の精神の構成の生物学的基盤が示されている。
原始爬虫類脳をもつ動物は、原始的な学習や記憶に基づいた、型にはまった行動を現わす。そして、この行動は個体維持と種族保存に基本的なものである。魚類や爬虫類では、大脳基底核が最高の運動機能の統合部位で大脳皮質は未発達である。これらの動物の行動反応は辺縁系と視床下部で開始され、そのパターンは柔軟性に乏しいタイプである。
旧哺乳類脳は哺乳類においてはじめて発達した。この脳部位には明らかな情動の座があり、ステレオタイプな原始爬虫類脳の働きを、ある程度、柔軟に制御している。その条件下で辺縁系と視床下部は行動発現の開始部位の一つとしてさらに重要な意味をもつようになる。すなわち、「情動過程」そのものに基づく行動発現と結びついている。
新哺乳類脳は高等哺乳類においてみられ、新皮質は、外界環境因子を分析し、高度の精神活動を行う。霊長類になると、大脳皮質、小脳、大脳基底核が著しく発達し、ここに行動発現に対する「認知過程」が関与してくる。新皮質と辺縁系との相互連絡は動物が高等になるにつれて発達し、とくに側頭連合野・前頭前野と辺縁系との間の線維連絡は密になり、情動行動はさらに洗練されて複雑になり、質的にも発達したものとなる。ヒトの脳内には言語野が発達する。抽象概念を用いた思考が可能となり、情動状態を自省し、洞察できるようになる。ここでは辺縁系に対する前頭前野の支配と、「思考過程」の「情動状態」への関与が特徴的で、解剖学的にも連合野と視床下部・扁桃体との直接的または間接的(嗅内野を介して)結びつきが強くなっている。
大脳辺縁系とくに扁桃体の機能異常は精神疾患のうちの情動障害と深く結びついていると考えられる。精神分裂病や躁うつ病の患者にみられる両価性や自閉症状や抑うつ気分などは広義の辺縁系領域の機能逸脱と考えられよう。ここで情動に関して強調しておかねばならないことは、、大脳辺縁系、視床下部、下垂体を問題にする以上、脳を単に神経回路網の集合からなる"硬い"刺激伝達系としてみるのではなく、"植物性"機能の面(内部環境および外部環境)、すなわち、自律神経系、内分泌系も含めたホルモンや伝達物質など液性伝達系としてみる立場も忘れてはならない。カテコールアミン、インドールアミン、ペプチド、糖質グルココルチコイド、ほか、多くのホルモンの作用についての知識が必要である(後述)。
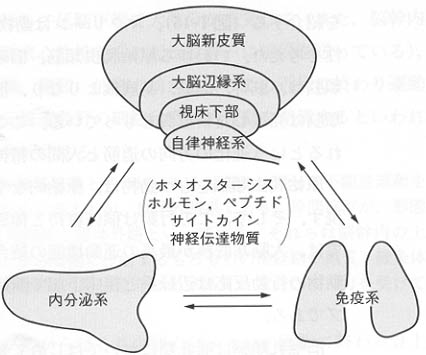
図8:神経系、内分泌系、免疫系の鼎談
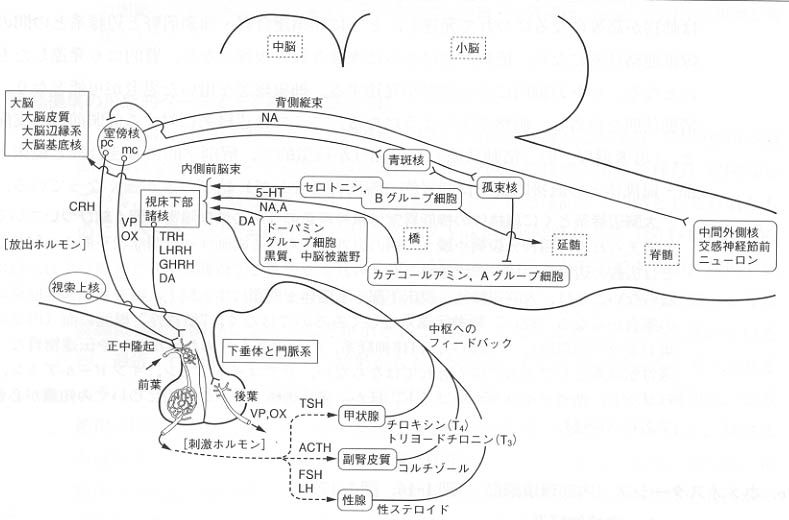
図9:脳を硬い複雑な神経回路網から構成される単なる集合体としてみるのではなく、自律神経系、内分泌系をふくめた液性調節系も含めてみるべきことを示す模式図。
自律神経系は体内からの情報を受け取り、内分泌臓器や免疫臓器を含む生体内諸臓器の調節を行っている。自律神経系の系統発生的起原は古く、魚類ですでに脳・脊髄とは別に交感神経幹に相当する中枢を形成しているものがある。自律神経には、交感神経と副交感神経がある。両生類では交感・副交感神経はすでに形態的には完成されているが、支配器官に対する作用はさまざまで、新しい環境との相互関係の中でいかに生命を維持し、種属を保存するかという目的にそって動物種独自の機能特性を獲得したと考えることができる。
交感神経は身体を活動的な状態にする働きがある。心拍数を増やし、血圧を高め、消化管の運動を抑制する。これに対して、副交感神経は身体活動を準備するように働く。心拍数は減り、血圧は下がり、消化管の運動は盛んになる。すなわち、交感性反応は身体の広い範囲に影響を及ぼすタイプであり、他方、副交感性反応はローカルに作用してエネルギーを貯めるタイプと考えられる。
一般に交感神経のニューロンの結合は一つの節前ニューロンが多数の節後ニューロンと結合しているために一部の興奮が広汎な作用を及ぼすことができるが、これに反して、副交感神経では一つの節前ニューロンがわずかの節後ニューロンと接続するため一般にその作用が限局する傾向にある。
副交感神経の節前線維は一定の脳神経および仙骨(脊髄)神経を通して現われ脳仙髄系
を構成する。他方交感神経の節前線維は胸神経と上部腰神経から現われ胸腰髄系
を作る。
副交感系の節後ニューロンの細胞体は末梢部に位置しており、支配する臓器の近くにはっきりした集団を作り神経節を形成するかまたは、ときには臓器の壁の中にうずもれている。
交感神経系の節後ニューロンの細胞体は一般に交感神経幹の神経節の中に、あるいは末梢にある神経叢中、支配する臓器よりも脊髄近傍に位置する神経節中に位置している。
すべての節前線維、副交感性節後線維あるいは体性遠心性線維の神経刺激の伝導は、終止部におけるアセチルコリンの放出と関連しているが、一方、交感性節後線維の場合には、その物質はノルアドレナリンまたはアドレナリンである。その故に、コリン作動性、アドレナリン作動性と呼ばれる。
ここで、自律神経の最高中枢である視床下部と脳幹や脊髄の自律神経関連諸核間の関連について一言しておく。バゾプレッシン
、オキシトシンを産生する室傍核や視索上核内の細胞は下垂体後葉に軸索を送る(ニューロンがホルモンを分泌する)が、そのほか、ある種の細胞(甲状腺刺激ホルモン放出ホルモンや副腎皮質ホルモン放出ホルモン含有細胞)は脳幹の孤束核、迷走神経背側核、青斑核や脊髄の中間帯との間に結合関係が存在することが知られている。また、視床下部は大脳辺縁系や前頭前野からの入力も受けている。
以上の自律神経系の機構をサポートする形で、ノルアドレナリン(橋、延髄より)、セロトニン(中脳以下脳幹より)、ドーパミン(中脳より)を伝達物質としてもつ神経投射系があり、それらは、前脳の広い範囲に分布している。
内分泌系の作用として、内外の環境変化に対応して、自律神経系と協調しつつ、生体の恒常状態を維持する(ホメオスターシス)ということがある。内分泌系の調節は、インスリンと血糖のように、内分泌腺と標的細胞の間のフィードバックだけで決定されるものもあるが、多くの場合には、視床下部、下垂体を介した複雑な調節が行われている。すなわち、①視床下部からの放出(調節)ホルモンが、下垂体前葉ホルモンの分泌を促進あるいは抑制する。②下垂体前葉ホルモンがそれぞれの標的器官に作用してホルモンの分泌を促す。③末梢ホルモンは標的細胞に作用してその機能を変化させるとともに、視床下部あるいは下垂体前葉にフィードバックをかけてホルモン分泌を抑制する。また、下垂体前葉ホルモンによる視床下部へのフィードバックもある。また脳は単にホルモンの調節器官であるのみならず、ホルモンの標的器官でもあって、たとえば女性性周期が正常に発現するためには、脳の発育過程において特定時期の特定部位にホルモンが作用しなければならない。さらにまた脳内のペプチドホルモンは、ホルモン作用をもつのみならず神経伝達にも重要な関わりがある。
また、さまざまなストレスが視床下部―下垂体―標的器官系に影響を及ぼすことはセリエSelye以来注目されているが、大脳皮質が発達したヒトにおいては外因的ストレスだけでなく内因的ストレスも正常な生命機能を阻害する大きな要因と考えられる。ストレスも内的・外的環境変化の一種のフィードバック機構を作動させるものととらえることができる。また、ストレスが個体に及ぼす影響についても個体がどのようなホメオスタティックな状態にあるかによって異なり、したがって個体のストレスに対する反応もそれによって異なってくることも知られている。
このように内分泌系は生体のホメオスターシスを維持し、さまざまな情報を処理しつつ複雑な統合作用を営み、ストレスへの応答にも重要な役割を演じている。そして、視床下部・下垂体を中心とする神経内分泌経路が、精神と身体の重要な接点となっている。このような意味で、精神神経内分泌学は精神医学の中で重要な位置を占めている。
上にみたように、ホルモンには内分泌腺から血中へ分泌されるもののほかに、視床下部・門脈・下垂体系で産生されるものがある。ここで、脳を液性伝達系から眺めると、視床下部ー下垂体ー末梢標的器官という側面が浮かびあがる。これには、以下の3系がある。
1) 視床下部ー下垂体ー副腎皮質 hypothalamo-pituitary-adrenal 系(HPA)
2) 視床下部ー下垂体ー甲状腺 hypothalamo-pituitary-thyroid 系 (HPT)
3) 視床下部ー下垂体ー性腺 hypothalamo-pituitary-gonadal 系 (HPG)
これに関係するホルモンとして、以下のものがある。すなわち、
① 視床下部のホルモンには、
成長ホルモン放出ホルモン(GHRH)、ソマトスタチン、プロラクチン[放出]抑制ホルモン(PIH)、甲状腺刺激ホルモン放出ホルモン(TRH)、副腎皮質ホルモン放出ホルモン(CRH)、 黄体形成ホルモン放出ホルモン(LHRH)、バゾプレッシン、オキシトシン
② 脳下垂体のホルモンには、
成長ホルモン(GH)、プロラクチン(乳腺刺激ホルモン)、副腎皮質刺激ホルモン(ACTH)、エンドルフィン、甲状腺刺激ホルモン(TSH)、卵胞刺激ホルモン(FSH)、 黄体形成ホルモン(LH)、メラニン細胞刺激ホルモン(MSH)、バゾプレッシン、オキシトシン
免疫系は自律神経系による神経性調節と内分泌系による液性調節を受けているが、さらに免疫担当細胞から分泌される種々のサイトカインにより調節されている。免疫系の機能は、多様化した細胞群と体液性物質との相互作用によって成立する生体防御機構である。すなわち、非自己による外界からの侵害(広義のストレス)に対して自己を防衛し内部環境を一定に保とうとする機能をもっている。免疫系には液性免疫と細胞性免疫があり、内分泌系や自律神経系の免疫系への関与としては、液性免疫の場合には免疫グロブリン(抗体)産生の調節に働き、細胞性免疫に関してはマクロファージによる情報提示やT細胞の分化成熟・環境の整備に働いている。心身にストレスが過剰に加わると、ホメオスターシスが崩れ、心身症や神経症やうつ状態に陥る。これは、ストレスに対する体の防御反応が、体にとって逆に悪い影響を与える方向にすすんでしまった結果である。脳がストレス状態を感じると(おそらくその最初は大脳辺縁系で)、次に、視床下部-下垂体-副腎皮質系(HPA)が働いて副腎皮質から糖質コルチコイドが分泌される。この状態が長く続くと内分泌系、自律神経系と並んで、もう一つのコントロール系である免疫系にも重大な影響を与える。
糖質コルチコイドはリンパ球の働きを弱めるなど免疫系の機能を抑制する。反対に、精神的なストレッサーが視床下部ー自律神経系に働くと副腎髄質からアドレナリンが分泌され、免疫力が強まる方向に流れる。以上は、脳から免疫系への影響であるが、免疫系から脳への影響についても、脳は免疫学的寛容であるという従来の見解に反して、異物獲得の情報などを末梢の免疫担当細胞(好中球やリンパ球など)が生理活性物質、インターロイキン(サイトカインの一つ)を介して伝えている。サイトカインは免疫担当細胞が放出する蛋白質であり、脳内サイトカインが神経伝達物質の代謝に影響を与え、その結果、動物の情動・行動面に変化を起こさせることもわかってきた。サイトカインはIL-1,IL-2,IL-6,TNF-αなど数多く存在するが、それらの脳内での作用部位、およびそこからホルモンやペプチドの分泌にいたる情報の伝達経路に関しては、現在明らかにされつつある。このように、免疫系は自律神経系および内分泌系と密接に関連しており、精神医学関連の疾病としては、ストレス負荷およびうつ状態における免疫機能の低下との関連が注目されている。
通常の、単なる記載神経解剖学ではなく、中枢神経系の成り立ちを基軸にして、脳を身体の全体の中でとらえるという立場が大切と考えて書いた。それは、とりもなおさず、精神の異常に悩む人たちと日常的につきあい、そして、患者さんたちに勇気を与えてあげられる職業をもった治療者の皆さんにとって、多少むずかしくても病気の原因を考えながら、ダイナミックに勉強して頂きたかったからにほかならない。内容が圧縮されて記載されているために理解が不十分な点は、実地を経験しながら関連の教科書も開いて、知識を自分のものにして欲しい。
1. 新生理学体系(監修:勝木ら)第12巻:高次脳機能の生理学(鈴木、酒田
編). (1988): 医学書院.
2. 生物学的精神医学(小島、大熊 編)第4巻:
認知機能からみた精神分裂病. (1993) : 学会出版センター.
3. 脳の科学―特集:情動・意欲の神経機構とその病態―、20/7 (1998)
: 星和書店
4. 脳と神経―分子生物神経科学入門― (金子、川村、植村 編)(1999)
: 共立出版株式会社
精神医学テキスト(上島、立山編、南光堂 2000年)より許可を得て加筆掲載