われわれの情動、認知、運動、意欲など高次神経活動の所産、すなわち精神活動は皮質連合野を主体とする脳全体の活動の結果として発現するものと考える。この神経活動の主体は大脳皮質、大脳基底核、視床、小脳から成る複数のニューラル・サーキット(神経回路網)が、システム全体として並列的に、連続して円滑に作動し続けられなければならない。そして時に自動性を帯びる。この神経機構の形態的基盤について考察した。十八番の民謡や器楽・声楽、いつもの小話や演説などは、これらの系がかかわる(仕)業であろう。もしも150歳のフロイトが現在の精神・神経科学の成果を共有し得たならば、どのように「精神現象」について語り、研究計画を練り直すであろうか?右往するか、それとも左転するか、それとも合一を謀るか?
連合野、小脳、基底核、運動と思考、情報の並列処理
動物は生活環境からの刺激を知覚し、認知し、認識する。これは大脳活動の受動的側面で、感覚野および感覚連合野(=後連合野)の働きに依存する。社会生活を営む人間の場合は、言葉を用いて人と交わり、環境社会に対して能動的に働きかける。この機能を制御し、統括する脳内領域は前頭前野(=前連合野)である。この神経活動の機能的モデルを小脳組織は如何にして作るのであろうか?
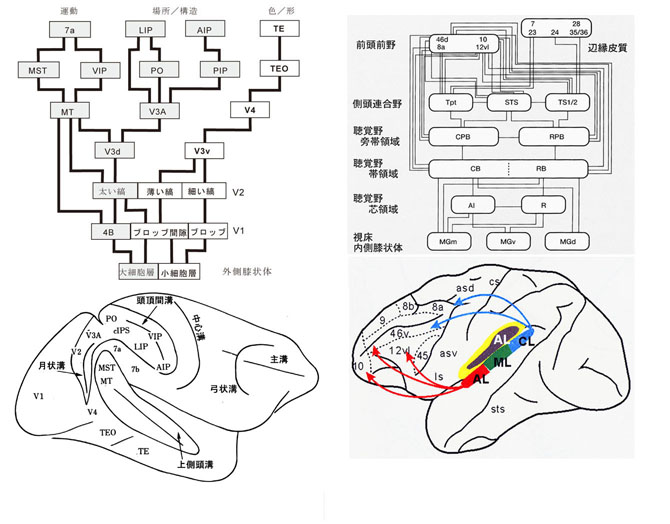
体内および体外からの刺激は、中枢神経系内を脊髄→脳幹→視床→終脳へと向かい、最終的には大脳皮質の知覚野(sensory area)で知覚・認知・認識される。知覚野は感覚野とも言われるが、嗅覚、味覚、体性感覚、聴覚、視覚に関する種々の領域に分かれる。視床の特殊核からの入力を直接受け、要素的な感覚刺激の分析に関与する第一知覚野のほかに、より高次の機能分析に関わる知覚性連合野も含まれる。これらの視床・皮質投射線維は、嗅覚路は前頭葉眼窩面皮質に、味覚路は前中心(脳)回腹側部および島皮質近傍に、体性感覚路は頭頂葉内の後中心回に、聴覚路は上側頭回内側部皮質に、視覚路は後頭葉内側面鳥距溝内皮質に各々終止する。第一知覚野に至るまで、相互間の干渉・融合はなく各々独立した神経路を構成している。皮質連合野において初めて、異なる感覚様態間の相互作用が起こり、知覚機能の融合がみられ、認知機能の統合がなされる。風景を見て音楽を想起し、音楽を聴いて色彩を帯びた形象が「脳裏」に浮かび上がるのは、大脳皮質連合野で起こる機能の発現/神経活動である。その形態的基盤であるニューロン群の機能の特殊分化、興奮伝達系および異種感覚間の連合・統合などの問題についてここで論じるには大き過ぎる。視覚系と聴覚系については図1で説明する。体性感覚系については酒田26) に詳しい。嗅覚系、味覚系および自律神経系や辺縁系への言及は割愛する。様々の感覚系間の融合・統合の問題も含めて拙書「脳と精神―生命の響き―」18) を併読いただければ幸いである。
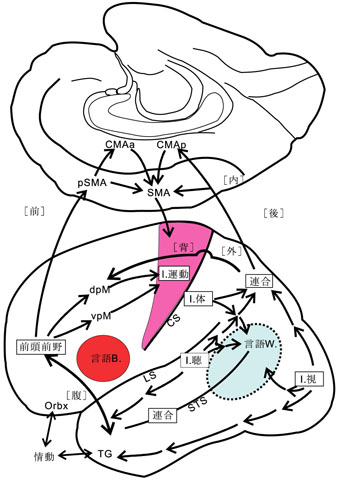
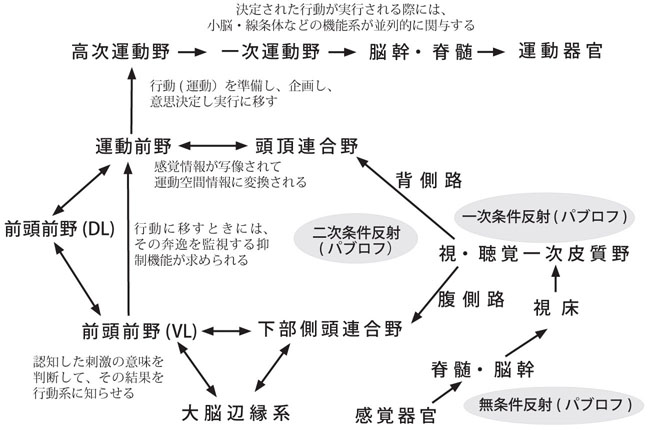
「受動性」機能(知覚・認知など)は頭頂・側頭・後頭葉皮質内の連合野、すなわち一括して後連合野と呼ばれる領域における高次神経活動であるが、便宜的に分けた、脳のもう一つの重要な機能として、前(頭)連合野あるいは前頭前野が関与する「能動性」機能(状況判断・行動・意欲など)、すなわち動物が環境に働きかける高次機能がある。換言すれば、後連合野での知覚・認知機能に、「後部言語野(ウェルニッケ野)」と辺縁系の機能(情動の要素・情報は、実は、側頭葉の他に前頭葉にも入力される)が加わって、その結果が前連合野に伝達される18)(図2)。この後連合野からの興奮は、「前頭前野」、「高次運動野」で状況に即して判断され、処理されて、その情報が「一次運動野」に伝達される。この「高次運動野」は運動の順序を決めて準備し、企画し、命令する運動機能に関連する領域である。能動的な運動はこうして実行される。前頭葉における言語(思考)、意欲、記憶などとの関連をも含めて、感覚・運動の情報処理全般について図3に示す18)。
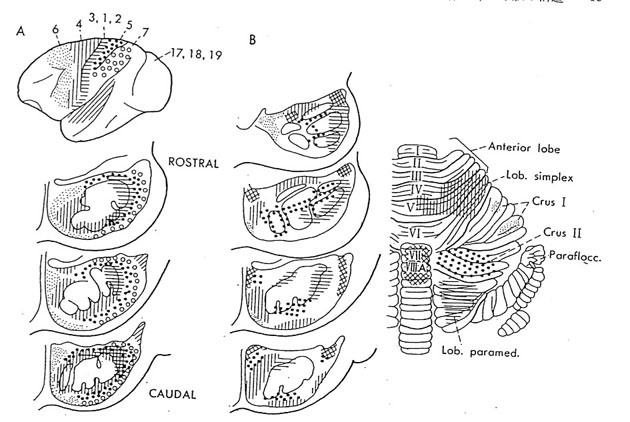
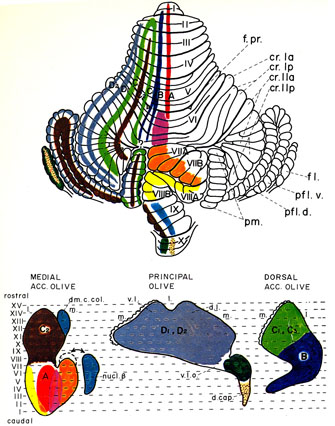
大脳皮質連合野は、動物が高等化に伴って発達する。領域的にも拡大し、皮質全体に占める割合も増大する。大脳皮質の感覚野、高次運動野、連合野を含む広範囲の領域から、橋核および下オリーブ核を経由して、各々苔状線維および登上線維が小脳皮質の広い領域に投射が行なわれている。その苔状線維投射4,5,17) および登上線維投射3,9,10,16) の局在関係(図4、図5)も調べられている。視床核を介する小脳皮質から大脳皮質への投射にも局在性が認められる。この小脳・大脳関連ループに関しては、小脳半球外側部は大脳皮質の運動野外側部、運動前野および前頭前野との間に、他方、小脳中間部は大脳皮質の全運動野(とくにその中間部)との間に相互連絡が存在することがSasaki 28) によって明らかにされた。
小脳は運動の調節・制御に関与するのみならず、認知・思考を含む言語機能、ひいては広く「精神」機能の制御活動にも関わっていることが注目されている。『小脳は mental skill に貢献しているか?』と題する論文を20年前に発表したLeiner ら21) は、ヒト歯状核外側部の障害症例において、行動を計画し、それを観念として実行するような予測能力が著しく低下していることを観察した。最近は、MRI, PET, rBF(局所脳血流)などを使用して言語やイメージを含む認知機構に、小脳半球、とくに後葉の外側部や歯状核が関与していることが明らかにされている22) 。
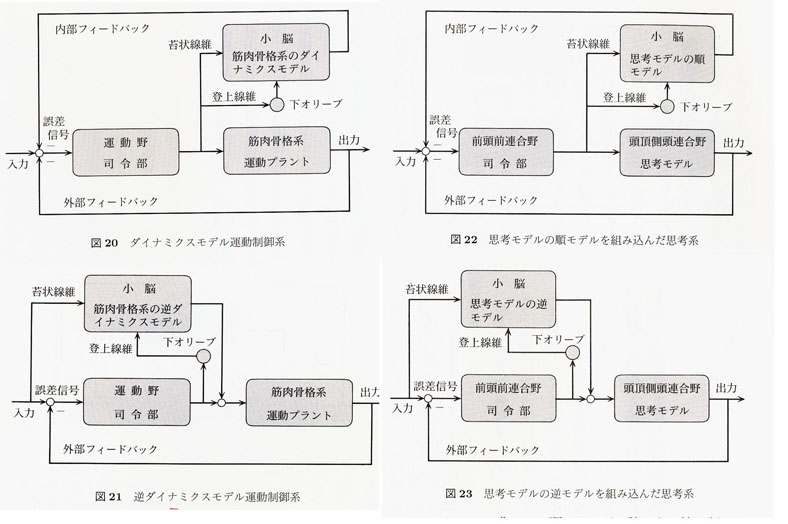
Ito13,14,15) は、大脳のフィードバック制御の働き、すなわち、「前向き制御」に転換する「予測制御」(工学用語を借用して)の働きを、小脳はもっていることを明らかにし、思考と運動の類似性に注目した。運動の場合には運動前野、補足運動野そして一次運動野を働かせて身体を動かすが、対して思考の場合には言語連合野を働かせて観念や概念を作り、思考過程を前向きに自動化する働きがあると推定した。伊藤は「思考は脳内にあるモデル化された概念を動かすことである」という意味の心理学用語を用いて「思考モデル」と呼び、小脳において提示された運動制御系を大脳のモデル思考制御系に適用(対応)させて提案した。すなわち、大脳皮質内でブローカ野を含む前連合野が、認知思考の要素の貯蔵庫であるウェルニッケ野を含む後連合野内に存在する思考モデルに繰り返し働きかけた結果として、ヒトが大脳皮質内活動として思考、すなわちさまざまに考えることをくり返すうちに、小脳と大脳皮質との間を両方向性に密接に結ぶ結合2,28) を使って小脳内にそのシミュレートされた思考モデルが形成されてしまえば、何度か、既に経験された思考に関して、改めて大脳皮質内活動をすることなく自動的に思考が進むことになるとした。「思考モデル」は以下の順序で進行する。すなわち、①前頭前野が後連合野内の思考モデルに働きかける。②この思考モデルの動特性をシミュレートするモデルを小脳内に作る。③前頭前野はこの小脳内思考モデルに働く。この過程を繰り返し続けることにより、半ば自動的に思考することが可能になり、思考モデルの逆モデルが小脳内にできれば、無意識に思考過程が進行するようになる13, 14, 19, 20)(図6)。
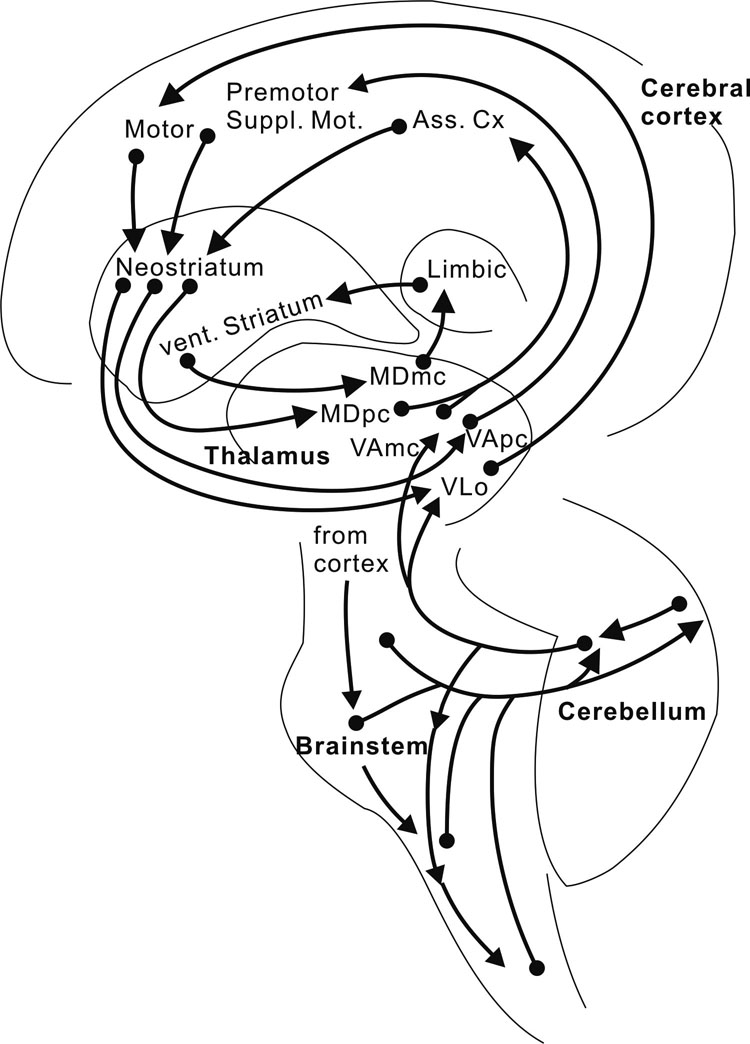
Alexanderら1) は、錐体外路系の一つである「大脳基底核-視床-大脳皮質」を結ぶ神経線維連絡は閉鎖回路を形成しており、この情報処理に関わるシステム回路は、形態、機能ごとに並列的チャンネルを作っており(parallel channeling)、個別的、並列的な情報処理(parallel processing)をする場であるという考えを呈示した。便宜上、運動系、連合系および辺縁系の3つの系に分けて考えると、運動行為、思考形成、情動発現など、一連の過程が円滑に進行するためには、「小脳・大脳関連ループ」に加えて、この「大脳皮質・基底核・視床パラレル・ループ」(図7)18)の関与が必要となる。
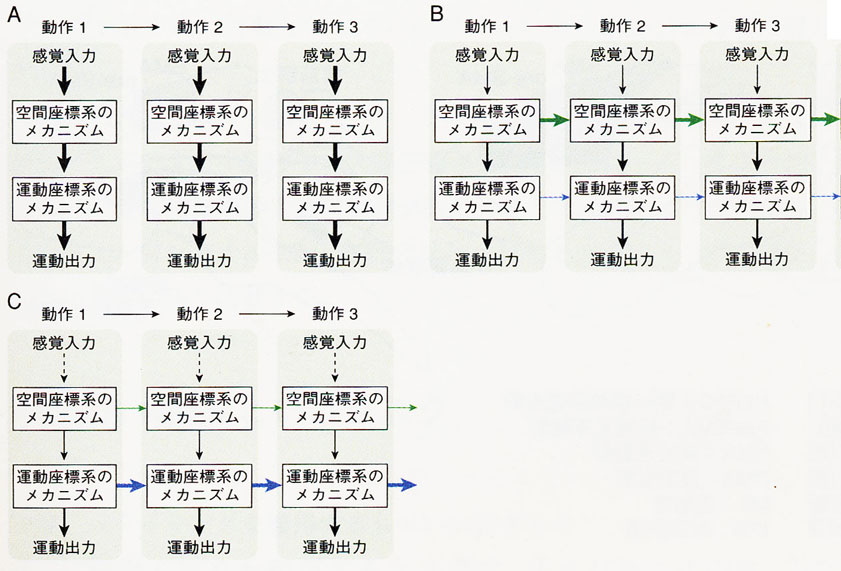
あらゆる種類の能動的行為の進行過程(working processes)において、複数の動作が連続して、同時に並行して作動し、それらの出力が連動してなければならない。近年、彦坂グループ12, 23, 24) は、この大脳皮質-大脳基底核連関に注目して、その手続き的(procedural) 運動記憶とその実行系の神経機構を明らかにすべく、サルを用いてニューロン活動を調べ(細胞外記録)た結果、以下の事象を明らかにした。すなわち、① (視覚空間座標でコードされる)学習の初期、つまり新しい課題を獲得しようとする段階では、前頭前野/前補足運動野(pre-SMA)/頂頭葉内側部などの連合皮質-基底核前部ループ(視覚ループ)が関与し、次に ②(運動座標でコードされる)学習がさらに進んで蓄えられた記憶を保持し、読み出し、スキルになった手続きを実行する段階になると、補足運動野(SMA)などの運動関連皮質-大脳基底核中央部ループ(運動ループ)が関与するようになる12)(図8)。
以上、簡単にまとめると、「前頭前野・大脳基底核前部・視床(MD, VA)核・小脳半球外側部」を包含する「大脳・小脳“連合”ループ」が形態基盤として存在し、この回路が“自動性”を獲得するほどに回転すると、大脳皮質連合野活動が小脳内にも“写像”されて、いわばミニ・コピーとして、小脳に潜在的に内在していた「高次精神機能」が発揮されるというストーリーとなる。この仮説は今後の研究によって検証されよう。ここで、少し厳しく考察してみる。① ヒトで調べた研究による、小脳への入力線維が多数である反面、出力線維は凡そその1/40に過ぎないという事実11a)、および ② 第一次世界大戦のとき小脳組織が銃弾で限局的に傷害された兵士の多くの症例で、概念形成能力は保持されていたという報告12a) は看過できまい。本来、metric & automatic & smooth な小脳の運動処理能力は、個体発生的にみても、系統発生的にみても、前庭機能や脊髄機能の円滑な反射機構に関連して、延髄の一部から形成されたその古い部分(原小脳Archicerebellmと古小脳Paleocerebellum)を包括した神経回路網が基盤になっているものである。高等哺乳動物の段階になり、大脳新皮質、さらに皮質連合野が発達すると、それらの新しい領域で処理されることになった認知や判断の情報を、一次運動野に直接的に伝えるというシステムを採らずに、その大部分を連合野→脳幹→小脳→視床→皮質運動野という神経回路を利用して、それを円滑(smooth)に、かつ能動的に働かせるようになる。小脳を「円滑装置」と呼ぶか、大脳の「モデル器官」と考えるか、これは研究者の視点の問題であろう。
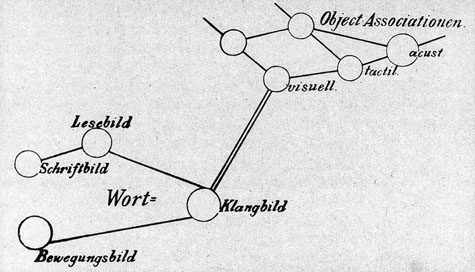
想い起こすに、今から100年以上も前に実行不可能として、Sigmund Freud (1856-1939) が公表を憚った“Associationen”を念頭に置いた「研究計画(草稿)」がある。夢の分析と解釈-無意識の世界に足を踏み込む前にフロイトは、心理学を“ニューロン”という概念を採り入れて、自然科学的に研究しようと意図して、友人のフリースへの手紙という形で計画(Entwurf /Project)8) を練った。この草稿は彼の死後、1950年になって、娘アンナと精神分析家クリスによって編集されて公になった。当時の脳研究のレベルがその段階に達していなかったので、高次機能に関係する脳内構造、とくに前-後連合野、高次運動野(=運動連合野)、言語野(=最高次に発達した連合野)などの用語を使用して相互関連やシステム機構について考察することはできなかった。しかしわれわれは、図9,図10の中に、フロイトが19世紀末(~20世紀初頭)における神経科学の到達点に立って、ヒトの精神現象をいかにシステムのレベルで解明しようと努めたかを読み取ることができる。
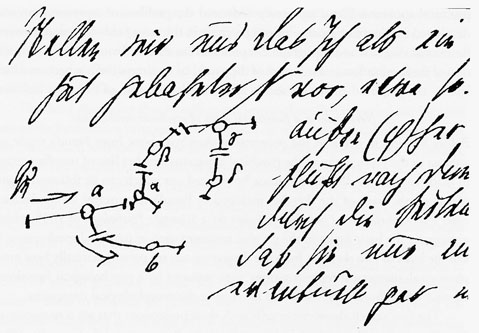
1) Alexander, G.E., DeLong, M.R., and Strick, P.L.
Parallel organization of functionally segregated circuits linking basal ganglia and cortex, Annu. Rev. Neurosci. 9(1986) 357-381.
2) Allen, G.I., and Tsukahara, N.
Cerebrocerebellar communication system, Physiol. Rev. 54 (1974) 957-1006.
3) Brodal, A., and Kawamura, K.
Olivocerebellar projection: a review, Adv. Anat. Embryol. Cell Biol. 64 (1980) 1-140.
4) Brodal, P.
The corticopontine projection in the rhesus monkey. Origin and principles of organization, Brain 101 (1978) 251-283.
5) Brodal, P.
The pontocerebellar projection in the rhesus monkey: An experimental study with retrograde transport of horseradish peroxidase, Neuroscience 4 (1979) 193-208.
6) Desimone, R., Schein S.J., Moran, J., and Ungerleider, L.G.
Contour, color and shape analysis beyond the striate cortex, Vision Res. 25 (1985) 441-452.
7) Flechsig, P.
Meine myelogenetische hirnlehre mit biographischer Einleitung, Springer Verl. Berlin, 1927. 但し、髄鞘発生研究の最初の原著論文は1898年に発表されている。
8) Freud, S.
Entwurf Einer Psychologie / Project for a Scientific Psychology, 「φψω」Theorie (1895); Sigmund Freud, Brief an Willhelm Fliess (1887-1904間の手紙で1950年に出版)。cf. Freud and the Neurosciences, From Brain Research to the Unconscious, G.Guttmann and I.Scholz-Strasser (Eds.) Verlag der Österreichischen Akademie der Wissenschaften, Vienna,1998.
9) Groenewegen, H.J., and Voogd, J.
The parasagittal zonation within the olivocerebellar projection. I. Climbing fiber distribution in the vermis of cat cerebellum, J. comp.Neural. (1977) 417-488.
10) Groenewegen, H.J., Voogd, J.and Freedman, S.L.
The parasagittal zonal organization within the olivocerebellar projection. II. Climbing fiber distribution in the intermediate and hemispheric parts of cat cerebellum, J. comp. Neurol. 183 (1979) 551-602.
11) 端川勉
聴覚野の機能解剖学、Clinical Neurology 24 (2006) 504-507.
11a) Heidary, H. and Tomasch, J.
Neuron numbers and perikaryon areas in the human cerebellar nuclei. Acta Anat (Basel) 74 (1969) 290-296.
12) Hikosaka, O., Nakahara, H., Rand M.K., Sakai, K., Lu, X., Nakamura, K., Miyachi, S., and Doya, K.
Parallel neural networks for learning sequential procedures, Trends Neurosci. 10 (1999) 464-471.
12a) Holmes, G.
The symptoms of acute cerebellar injuries due to gunshot injuries. Brain 40 (1917) 461-535.
13) Ito, M.
Movement and thought: Identical control mechanisms by the cerebellum, Trend Neurosci. 16 (1993) 448-450.
14) 伊藤正男
脳の不思議、 1998、岩波書店
15) Ito, M.
Neural control of cognition and language, In: Image, Language and Brain, Maranz, A., Miyashita, Y., and O’Neil, W. Eds., Cambridge, MA: MIT Press, 2001, p.149-162.
16) Kawamura K., and Hashikawa T.
Olivocerebellar projections in the cat studied by means of anterograde axonal transport of labeled amino acids as tracers, Neuroscience 4 (1979) 1615-1633.
17) Kawamura, K., and Hashikawa, T.
Projections from the pontine nuclei proper and reticular tegmental nucleus onto the cerebellar cortex in the cat. An autoradiographic study, J. Comp. Neurol. 201 (1981) 395-413.
18) 川村光毅
脳と精神―生命の響き―、2006 慶應義塾大学出版会
19) Kawato, M., and Gomi, H.
A computational model of four regions of the cerebellum based on feedback-error learning, boil. Cybern. 68 (1992a) 95-103.
20) Kawato, M., and Gomi, H.
The cerebellum and VOR/OKR learning models, Trends Neurosci. 15 (1992b) 445-453.
21) Leiner, H.C., Leiner, A. L., and Dow, R. S.
Does the cerebellum contribute to mental skills ? Behav. Neurosci. 100 (1986) 443-454.
22) Mathiak, K., Hertrich, I., Grodd, W., and Ackermann, H.
Cerebellum and speech perception: a functional magnetic resonance imaging study, J. Cognitive Nerosci. 14 (2002) 902-912.
23) Miyachi,S., Hikosaka, O., Miyashita, K., Karadi, Z., and Rand, M.K.
Differential roles of monkey striatum in learning of sequential hand movement, Exp. Brain Res. 115 (1997) 1-5.
24) 中村加枝
大脳皮質-基底核関連と情報統合-手続き記憶と閉回路システムによる‘Executive function’、 Brain Medical 13 (2001) 343-352.
25) Romanski, L.M., Bates, J.F., and Goldman-Rakic P.S.
Auditory belt and parabelt projections to the prefrontal cortex in the rhesus monkey, J. comp. Neurol. 403 (1999) 141-157.
26) 酒田英夫
頭頂葉、2006 医学書院、
27) Sakata, H., Taira, M., Kusunoki, M., Murata, A., and Tanaka, Y.
The parietal association cortex in depth perception and visual control of handaction, Trends Neurosci. 20 (1997) 350-357.
28) Sasaki, K.
Cerebro-cerebellar interconnections in cats and monkeys, In: Cerebrocerebellar Interactions, ed. by Massion, J., and Sasaki, K. pp. 105-124, Elsevier, Amsterdam, 1979.
分子精神医学 7(2007)27-36 (先端医学社)より 許可を得て転載