
"連合野"の線維結合(I)皮質間結合
--サルとネコの皮質間結合の比較と"連合野"の発達についての試論--
Corticocortical fiber connections of the "association"areas. Comparison of
the fiber connections of cats with those of monkeys with comments on the development of
the "association"cortex
川村光毅
皮質連合領という言葉は現在,厳密な意味ではいわば共通の定義なしに用いられている。神経科学には種々の側面があり,各分野で得られた研究成果をもとにある程度の総合的な規定もできよう。学問は,その時代がもつ制約内で,"意味論"に流れることのない言葉の定義を要求する。これに答えることは研究者の務めでもあり,同時に,実際的な意味も否定できない。この特集号で他の執筆者たちにより発生学,皮質下結合,生理学,行動学などそれぞれの立場から考察が加えられるので,私は多少あいまいであっても,ひとまず"皮質連合野"を「大脳皮質のうちで,"第一次"感覚野と運動野を除いた新皮質の領域」と考えておくことにする。
ヒトの連合線維についてわれわれは,隣接する脳回を結ぶ短い弓状線維と,前頭葉とか後頭葉とかの異なる葉間を結ぶ長い連合線維の神経路の存在を肉眼解剖学の所見として得ているにすぎない。後者は上縦束,鈎状束などの名称が示すように,起始も終止も未確定の神経線維群である。したがってヒトについての確実な知見を欠く現段階では,動物によって得られた実験結果からその基本点を見出し,さらに動物一般の連合線維についての性質をつかんだ上でヒトの大脳皮質の働きを推論する道が残されている。
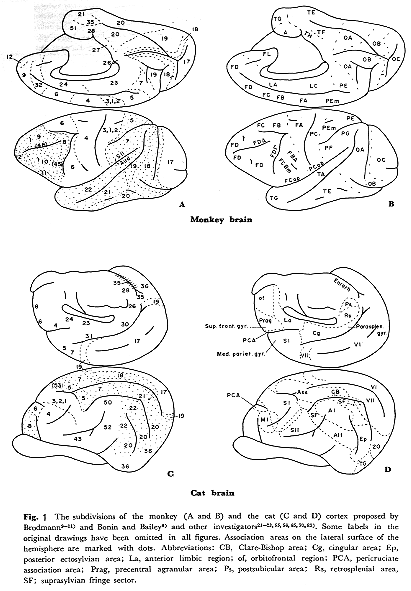
ネコとサルの皮質間結合については,最近十年間,かなり詳細に調べられてきた。私自身も,ネコの大脳皮質外側面の各機能領野から起こる連合線維について,Nauta法51,52)とFink-Heimer法16)を用いて明らかにしてきた34~46)。現在われわれはネコとサルでhorseradish
peroxidase(HRP)法を用いてさらに研究を進めているが,サルのFink-Heimer法による自家所見は持っていない。なお本稿では,批難を承知のうえ,研究者により今まで発表された知見を私なりに"整理"してサルとネコの皮質連合野の皮質間結合(求心路と遠心路)を示す模式図を作成することにした。この模式図作成は大胆な試みで,数人の研究者が得た所見結果を"標準"脳図という一つの図にまとめて書き入れたために,総体的理解という点ではそれなりの便宜的価値もあろうが,個々の所見結果を忠実に提示するという目的からは多少ずれるので私自身も最上の方法とは思わない。個々の所見の詳細については原著にあたっていただくことでお許し願いたい。ネコの標準脳図については私の作成したもの38)を用いたが,サルについてはその脳溝の変異を系統的に調べて作図したものがないので,Bonin
and Bailey 8)のものを多少改変して用いることにする。
個体による脳溝の位置,走行,形成程度に差異("変異")があるので,最も忠実な方法として個体ごとに変異の状態を記載し,脳回や脳溝との位置関係から大脳皮質部位の表現をすることが望まれるが,この方法ではいたずらに表現形式のみが複雑化するおそれがある。その上,私は,本論文の記述にあたり個体差を捨象する方法をとることにしたので,現在広く使用されているBrodmann
9~11)とBonin and Bailey 8)の脳区分をここで記述の便を助けるために採用する。なお,Brodmannが用いたサルはCercopithecus
(Macaca mulattaと同じオナガザル科で脳溝の走行も類似)で,かつネコについての詳細は発表されていない(脳溝走行が似ているものにクマのものがある)。Brodmannが皮質区分にアラビア数字の領野番号記載方式を用いたのに対して,Bonin
and Baileyはローマ字を用いてサル(Macaca mulatta)で調べているが,近縁関連皮質の類推に便宜を与えている。私は図1にこれらを単に提示することにとどめたい。なお最近の研究者による形態的および機能的区分名称を,読者の便宜を考え,とくにネコの図で一部加えた21~23,55,56,65,70,83)。各区分の比較および研究者間の分類区分についての比較を示すことは必要ではあるが,私見としてもまとめていないので省略することにしたい。本稿では便宜的にたとえばPE(5)野のように記載することもあるが,これはサルとネコの所見の比較の便を念頭においたためである。
一般に,皮質連合領は前連合領(前頭前野)と後連合領(頭頂連合野,後頭前野,側頭連合野)とに大別される。この名称は便宜的であるという点から考えて,サルにおいては,まず問題点は少ない。しかし,ネコにおいてはかなりの問題がある。第一にネコの9,10,11,12野(FD野)に相当する皮質部分の有無とその位置決定,第二に20,21,22野間の境界を含めてサルとの類同ないし相同の問題,第三に,両動物共通の問題として後頭前野が第2視覚領野(18野と19野,またはOBとOA)に相当することがら,同一基準82,83)で他の第2感覚領野であるSII,AII域を連合野内に含めることの妥当性の問題,第四に,第三の問題と密接に関係するがこれら第2感覚野一般の共通性を論じておかなければならない点,第五に内側面および底面の皮質部分の連合線維の研究が遅れている点,第六に,大脳皮質の全域中かなりの広い部分を占めている脳溝内皮質部分の知見の不足など多くの問題に直面する。それゆえ,今後の神経科学の研究により上記の問題点が明らかにされ,信頼のおけるデータが現われた時点であらためて考えるとの楽観的態度をとらないかぎり,筆は進まない。このような状況下で私は,一般的区分けの方法に従って,動物(ここではサルとネコ)の皮質連合領を,1.前頭連合野(前頭前野),2.頭頂連合野,3.側頭連合野,および,4.後頭連合野(後頭前野)に分け,まず,それぞれの求心性連合線維(入力)と遠心性連合線維(出力)についてみていくことにする。
各連合野の占める領域は広く,それぞれ溝造上も機能の面でも細分化された特質をもっていることが明らかにされつつある(本特集号の他の執筆者により詳しく論じられるであろう)。したがって,それに応じた形の記載が要求される。私は起始と終止の部位が明らかな神経路については,図の上で努めてそれを表現したい。しかしながら,たとえば「サル外側面の月状溝周囲の皮質部分(18野,OB野)から弓状溝前方の小部分(8α,8S野,FD野)へ弱い投射の存在がNauta-Gygax法52)でKuypers
49)らにより明らかにされている」と書くかわりに「前頭前野はわずかながら後頭前野からも連合線維を受ける49)」と記載したいと思う。それは意図的に表理内容を低下させているのではなく,主な理由はここで私が扱う対象が広すぎるので文章の簡略化が要求されるためである。
サル(図2F):細胞構築学的には前頭顆粒皮質(frontal granular cortex)に相当し,Bonin
and BaileyのFD野,Brodma㎜の8~12野を含む(図1)。機能的には遅延反応(delayed
response)28)や弁別学習,行動のプログラミング50),短期記億などに関連する領域といわれている。またWalker
79)はBonin and Baileyが9および10野と名づげた主溝(sulcus principalis)の背側と腹側に位置するかなり広い部分を46野,その後方で弓状溝(sulcus
arcuatus)の前方域を45野としている。ヒトの45,(46)野は下前頭葉後部にあたりBroca中枢と呼ばれ運動性言語機能に関連する部分として知られている。この前頭葉域は視床のMD核と局在的相互結合をもつ領域で,連合線維を隣接する6野(FC,FB)からのほか,PG,OA,0B,TA,TEなど広範囲の後連合野からうける。しかしながら,運動領やいわゆる第1感覚野(SI,AI,VI域)から直接線維は受けない。すなわち,頭頂連合野(PG)からは弓状溝とその付近の6,8野に多く投射するが49,59),一部主溝の背側部にも線維を送る33)。後頭連合野(主としてOA,OB)からは弓状溝の前方域(主として8野)および主溝腹側部の9,10野に,一方側頭連合野のTE(20,21野)からは主溝腹側部へ,TA(22野)からは主溝背側部へ終わる49,59)。このように一部終止域の重複はみられるがかなりのorganizeされた系の存在を予測させる。特徴として,一般に前頭前野への皮質求心路の集中性は主溝腹側部に強い。一方,前頭葉極は,遠心路が多いのに対して(後述)他の葉からの求心路が非常に少ない。このことから,この領域への入力は前頭連合野内でいったんシナプスを代えたものが多いと考えられる。われわれのHRP所見も以上の所見と考察を一部裏づけている(未発表)。この前頭前野への求心路についての皮質-間脳間の結合を含めた精細な研究は前頭機能解明の上から今後とくに望まれる。
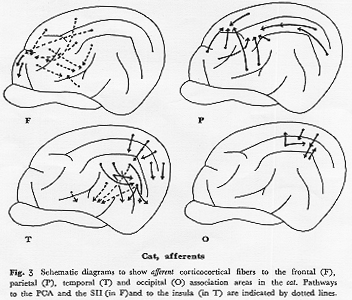
ネコ(図3F):図1に明らかなごとく,前頭連合野の発達は非常に悪くBrodmannの8野がわずかに舟状回とそれに隣接する内側面にのみみられる。この動物における9,10野などの発達の有無,およびその位置についての見解を筆者はもたない。今後前シルビウス溝や十字溝の壁面皮質および眼窩面皮質についての研究成果を期待したい。この十字溝周囲皮質,とくにその腹側面皮質はThompson,
Johnson and Hoopes 70)が誘発電位法による研究からpericruciate association area
(PCA)と命名した領域で,細胞構築学上では6,4野が主体 22)であるが,頭頂連合野との関係が密接である40)ことから強いて類似を求めればサルの弓状溝前方域に相当すると思われる。視覚野からはわずかに前外側回の19野からのみPCA域への投射が存在する可能性がある40)。一方,ネコの弁別学習,時間認知などの研究66)は少ない。その理由の一つは,舟状回,前S溝への求心路に関する確かな形態学的データの不足にある。また,第2体性感覚野(SII)を,後頭前野にOA(19)野,OB(18)野を当てたと同じ基準で前頭前野に含めるとすれば,この領域には側頭葉からの入力が入る。以上述べたように,広義に解釈すればネコの前頭連合野はサルと同様,他の皮質領野から連合線維をうけるが(破線,図3F),その発達はサルに比して格段に劣る。
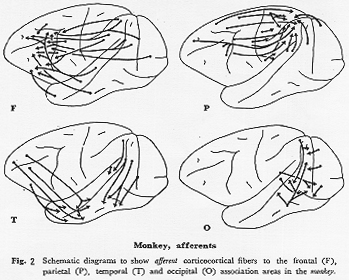
サル(図2P):主として視床核LP,LDと結合を有する領域でBonin and
BaileyのPE,PEm,PG,PF野,Brodmannの5,7野が占める。サルではヒトの上頭頂小葉に相当する部分のみが発達しており,下頭頂小葉に相当する40野(縁上回),39野(角回)は未発達である。サルが高等化する過程でこの領域に対応する部分が発達してくるとすれば7(PG)野の腹側,19(OA)野の吻側,21(TE)野,22(TA)野の背側にある上側頭溝周囲皮質部分におそらく由来する。この点に関しては読者はJones
and Powell 33),Kawamura 44)の論文および連合線維と皮質機能についてまとめた川村の総説
42)を一読されたい。
前頭連合野の機能が時間的要素の情報処理に関与するのに対してこの頭頂連合野は空間的要素の情報処理と密接に関係している。すなわち体性知覚性の身体各部の定位が主として5(PE)野に,視覚性空間の定位が7(PG)野に関係すると思われる。聴覚性のfieldについては,実験的条件の困難性のためかよく調べられていないが,おそらくPGの腹側部でシルビウス溝後背側端付近の皮質が関与するのではあるまいか?
求心性連合線維結合についてみるとサルの頭頂連合野は他の皮質領野からかなりの量の線維を受ける。すなわち,PE(5)野の前方域は体性知覚野(PC)から,後方域はFB(6)野から多く線維を受けており32,
33, 59),後者はその他に頭頂間溝に接した部分で視覚性の入力も受けている33)。それゆえ,頭頂間溝背側部分の機能を論ずる場合その前方域と後方域とで異なることが充分予想されるので,部位を正確に明示することが望まれる。またその最後方部分はPG(7)野となる8)が,PE(5)野との境界設定も変異があることが予想されるので,個々の動物についてそのつど決定したのち所見結果を論ずべきである。
一方,頭頂間溝腹側部分(PG)は,皮質間結合の様式において背側部分とは大分異なる。すなわち,この領域にはPC(3,1,2)野からの入力はほとんどなく,FB(6)野を含めて前頭前野(FD)からの成分が多い33)。また後頭前野(OA,OB)49)や第1聴覚野
83)やTE野の限局した背側部がら58)それぞれ視覚性および聴覚性の入力をうける。われわれの研究室で最近得られたHRPによる研究結果でもこれらを裏づけている(宋発表)。またこの領野が中,下側頭葉(TE,20,21野)からの求心線維をわずかしかうけないことは注目される。
ネコ(図3P):註釈なしでネコの皮質連合領といえば通常5,7野 22)を含むこの頭頂連合野を意味する。細胞構築学的にこの領域はGurewitsch
and Chatschaturian 21),Hassler and Muhs-Clements 32), Sanides and Hoffman 67)の三つの著者グループにより研究されており,研究者間で範囲や名称が少しずつ異なるが,その比較考察は私の他の論文40)を参照されたい。また,電気生理学的には誘発波の潜時が長く,波形や出現が不安定である反応を特徴とするpolysensory
responsesが得られる領域で,Amassian 2)の仕事をはじめとして,近年ではAMSA,PMSA,ALAおよびPCAの領域に分けたThompsonら70)の仕事がある。また最近,PMSAの神経細胞のユニット活動電位を調べた研究で多感覚性を示すニューロンが大部分を占めることが明らかにされ64),行動学的研究80)もなされるようになった。
求心路としては,体性知覚運動領31,46),聴覚領39),視覚領41)から線維をうける。この連合野の腹側を境する中S上溝をかこむ皮質領域(middle
suprasylvian sulcus area, MSs域)の線維結合と機能的な考察は別にすることにする。
全体としてみると5野は体性知覚野から,7野は視覚領野からの求心線維がそれぞれ主力を占めており,5野と7野の中間域付近は聴覚野からも線維をうけ,そのほかに前外側回前端部に位置する"Amassian"の領域(53野21))からの求心路が終わる。またThompsonら70)のALA,AMSA,PMSAは連合線維結合の立場からみるとそれぞれ体性知覚運動野,体性知覚と聴覚野,視覚野と聴覚野に関連が深いと思われる。このようにネコの頭頂連合野においても,サルでみたと同様の構成パターンが基本的に存在しており,機能的にほほ三つの部分に分けられるように思う。なおこの連合領野内の相互の結合関係についてはKawamura
40)のFig.8を参照されたい。
サル(図2T):本連合野の主要な部分はTA(22)野とTE(21,20)野でおよそ上,中,下側頭回に対応する。Brodmannによるヒトの脳図にはTEとOAの境界部に37野が命名されているが,サルにおいてもそのrudimentaryな対応が認められよう(TEOまたはPH
27,61))。一方島域(Reil)であるIB,IA野もsylvius溝内に埋れた連合野として含め得るし,このほかallocortexへの漸次移行野として内側面皮質のTFとTHおよび側頭極のTG野を区別し得るが,本論文ではこれら領野への連合線維の投射は後に述べることにする。このように側頭葉はほとんど"連合野"のみから成ることは特徴的である。
側頭連合野への求心性線維のパターンをみると二,三の特徴が抽出される。第一にかなり多くの長い連合線維が存在すること。第二にこれらのすべてが他の連合野内に起始細胞を有しており,FB,FA野や"第一次"感覚野からの入力はほとんどないこと。第三に局在関係が比較的明瞭な点である。すなわち,側頭葉の前半部は前頭前野(FD)から33,57,59)またその後半部は,聴覚領58),PG(7)野59)および視覚連合野(OA,OB)から33,49)入力をうける。これらのうちでも,FD野の主溝より背側部分からは主としてTA野に,主溝の腹側部分からは主としてTE,TG野に投射する傾向がある。また,OA,OB野からはTE野に,一方聴覚領やPG野からはTA野にと明らかに区別されて投射する。TE野(inferotemporal
cortex)の機能を破壊実験後の視覚弁別学習の影響という点から調べたIwai
and Mishkin 27)やBlake, Jarvis and Mishkin 7)の仕事は前方部と後方部の機能上の差異を明示している。一方TA野の仕事は現在遅れているが,聴覚弁別学習の仕事の困難性を克服した実験データが望まれる。なおこの連合野は視床結合の上で視床枕と密接な関係を有する。
ネコ(図3T):ネコの側頭連合野という術語はまだ定着されていない。その責任はおそらくネコの側にあらずして研究者の側にある。すなわち,この領域の解剖・生理学的研究は乏しい。したがってサルとの対比も困難である。第2視覚領(VII,18,19野)を後頭連合野(後頭前野)としたので,同様に第2聴覚領(AII)やEpさらにはSF域(suprasylvian
fringe sector)を連合野に入れて考えてもよいという理屈が成り立つ。しかし私はこれらの"paraauditory"cortexをサルの項で特記しなかった。イヌやネコの段階では聴覚領皮質はほとんどが脳回表面にあるが,サル(赤毛)になるとsylvius溝内皮質に限局して存在するようになる。その最大の原因は側頭連合領,とくにTE,TG野の発達にもとづくものであろう。一方TA野,とくにその後部は聴覚領に非常に近い領域で,皮質間結合および位置的関係からネコのSF域との類似を考えたくなる。このように考えてくるとネコ側頭連合野の範囲はmainの聴覚領(AI)をとりまくAII,Ep,SFおよび20野とsylvius脳回の腹側部を含む広い領野となる。これは一見妥当のようにみえるが,このように皮質レベルのみで論じていくとサルの連合領や連合線維群のパターンとの比較を論じる上で必ずしも適当ではなくなる。そこで皮質視床間結合の上から視床枕への投射を重視する立場からネコの本連合野を"paraauditory"areaを含めておよそ20,21,22野の範囲とすればサルのそれとほぽ一致し,比較しやすくなる。
ここでもう一言述べておくが,Heath and Jones 23)は線維結合の研究から細胞構築学上の19野55)の幅を腹側に広くとった。これにともない21野も中および後S上溝内の皮質にまで広がる。事実19野と21野の境界をNissl標本上で引くことはむずかしい。Heath
and Jones 23)の18,19,21野を合した部分は第2視覚野82,83)にほぼ一致する。サルの上側頭溝が21(TE)野と22(TA)野を境するように,ネコのsuprasylvius溝がS上回部分(21野)とS外回部分(22野)とを境するとしても,この領域は変異の多い部分である38)ことから漸次移行の形態をとるものであろう。私はこの変異の多い(ということはしばしば発生学的に"新しい"部分であることを推察させる)S外回の後背側の部分を皮質間結合の所見結果から,その後背部分が視覚性(連合)領域,前腹部分が聴覚性(連合)領域であることを提唱したが39),これはそれぞれをClare-Bishop域およびSF域の延長部分と考える立揚23)に一致している。
異論が予想されるところなので皮質の部位的考察をやや詳しくしたが,このようにネコの側頭連合野を考えると,20,21野は視覚領から41),22野は聴覚領から15,39)の線維をうけることが明らかになる。この関係はサルの側頭連合野と同様のパターンを示しつつ視覚および聴覚性の求心性連合線維をうけるという点で一致する。しかしながら,ネコの側頭連合野は前頭前野から直接線維を受けないという点でサルと異なる。ここに,ネコの前S回(43野21)および島域83))に対しても第2体性知覚領からわずかながら連合投射が存在する46)ことを付記しておく(破線,図3T)。
サル(図2O):典型的正常層形成顆粒皮質を示すOC(17)野をかこみOB(18)野,OA(19)野を合した後頭前野が発達している。この構成はヒトやネコでも基本的に同じである。Bonin
and Bailer 8)はBrodmannのいうように18野と19野の間は明瞭に境されないと述べており,脳図でみる限り,前者のOA野は後者の19野よりも外側面で範囲が広い。しかし個体差についての記載がないのでこの両研究者間の差異をそのまま受け取ってよいかどうか疑問である。本連合野は内側面に広い領域を占めるが,内側面皮質に関する知見は乏しく,今後の研究課題である。なお19野は前腹方で頭頂・側頭連合野に隣接し,機能的には立体視や空間的動きの識別に関与するといわれている25,26,84,85)。
後頭前野は,第1視覚領33,49)からのほかに,第2聴覚領ないし第2体性知覚領
59),側頭連合野(TA,TE野)の後部49,58,59)さらにはPG(7)野59)などかなり広範囲の他の連合野からも線維をうける。ただし,前頭連合野(FD)やSI,MI,AIからの求心線維はない。
本連合野でも,17野からのものを除いて上述の連合線維をすべて広範囲にうけているのではなく,体性知覚性の入力はその背側部に,聴覚性とTA,TEからのものは腹側部にというように局在性が比較的明瞭であるようにみえる。
ネコ(図3O):ネコの場合も18,19野の領域設定は研究者21,23,25,55,82,83)により多少異なるが,大体において一致している。細胞構築学的研究をもとにしたOtsuka
and Hassler 55)の脳図に従うのが一般的であろうが,ここでは線維結合の研究結果から提唱されたHeath
and Jonesのmap(Fig. 23)に従うことにする。
本連合野への求心路の発達は,他の連合野へのそれらにくらべ著しく劣る。すなわち第1視覚領以外からは,7野後部と前述の"視聴覚混成野"39,42)からの入力をうけるのみである。最近Woodyら81)条件付けをしたネコを用いて聴覚刺激に対するこの領野に存在するニューロンの活動性を調べているが,この領域のうちS上溝に近い部分は,側頭・後頭・頭頂の各連合野の接触する部分であり,単にauditory
association cortexと註釈なしで用いる81)のは適切でないように思う。この皮質領野(とくに後上S溝寄りの部分)を細区分して皮質内ニューロンの性質を調べてみるならば,サルのinferotemporal
cortexの最後部が有する27)ような視覚弁別学習との関連を示唆するデータが得られるように思う68)。
ここで視点を変えて,各連合野から起こる遠心路について簡潔な記述を試みる。
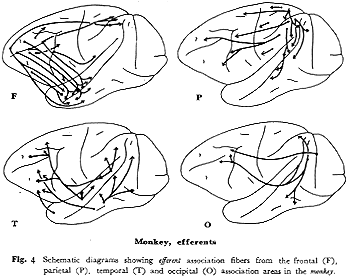
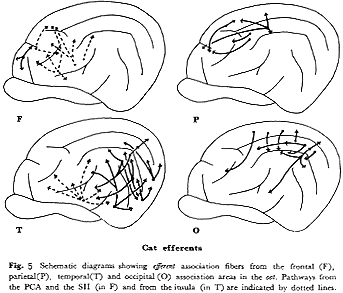
サル(図4F):本連合野からの遠心路は1)頭頂連合野(主としてPG野),2)側頭野前部,3)中古皮質(帯状回)へと大別される33,57,59,60)。PG野への起始細胞はFD野のうちでも弓状溝前域(6,8野付近)に多い。主溝隣接域を含めて,その背側部分はTA野と,腹側部分はTG,TE野との相互結合関係が認められる。さらに特徴的なことは,これらの連合線維群がサルの主要溝である頭頂間溝(intraparietal
sulcus),上側頭溝(superior temporal sulcus)および帯状溝(cingulate sulcus)をかこむ皮質部分に数多く終止する。なおこのほか,前頭前野はその広範囲から6野(FB,FC)に弓状線維を送るが,体性知覚運動野(1,2,3,4野)には送らない
33,59)。
ネコ(図5F):8野から6野への線維投射がある46)ほかは連合線維の発達は悪い。図5にPCA域とSII域から頭頂連合野の最前部(5野)にいたる神経路を破線で示したが,たとえこれらの域を前頭連合野に含めたとしても(1089頁参照),サルにみられるような頭頂葉やさらには側頭葉との豊富な結合はない。
サル(図4P):PE(5)野からは6野とPG(7)野への遠心性線維がほとんどである33)が,PG野からは4,6野のほかに,前頭前野(FD野)のかなりの範囲とくに主溝の背部と弓状溝前方野域に線維を送る33,59)。このほか上側頭溝の両壁,主にその背半部とそれに接する19,18野に投射する33,59)。しかし側頭連合野の前方域には直接連合線維を送らない。このようPE野とPG野とでは遠心性連合線維の投射パターンに明らかな差異を有する。
ネコ(図5P):サルにくらべて明らかに発達が劣る。すなわち,5野からはPCAとSI域に,7野からはPCAと18,19野およびClare-Bishop域に連合投射があるのみで側頭葉領域との結合はない40)。なお本領域からは,内側面の中古皮質(帯状回)に投射がある14,23,40)。
サル(図4T):TE(20,21)野の前方部から前頭前野(FD野)の腹側部に,後方部から後頭前野の前腹部に投射する33,59)。一方,TA(22)野の前方部からは6(FCBm)野およびFD野の主として主溝腹側部に
33,49),後方部からはPG野に線維を送る59)。TA野とTE野との間の結合がきわめて弱い59)ことは注目に価いする。第1体性知覚領や第1視覚領への直接の投射はない。
ネコ(図5T):(20,21)野からは後頭連合野(18,19野)およびAII,Ep領へ,また22野からはAI,SF領へ線維を送る。しかしながら頭頂連合野への投射はきわめてわずかである。一方,前sylvius回(島域)からSII全域に強い投射とSmIの顔面域に弱い投射がみられるが(破線,図5T),サルの場合と異なり前頭前野へは線維を送らない15,39)。
サル(図4O):体性知覚運動野や聴覚野に連合線維を送らず,特徴ある投射パターンを呈する。すなわち,17(OC)野へは少量の線維を送るが,PG野,TE野の中央部より後半部および弓状溝前方域とくに主溝腹側部にかなりの線維を送る49)。
ネコ(図5O):頭頂連合野(5,7野)への強力な投射があるに比して,側頭葉(主にSF野と島域)への投射は弱い。前頭葉へは前外回の前部(19野)からSI野にわずかの投射があるのみで前頭前野(8野)との相互結合はない41,46)。この点,サルの場合と異なる。
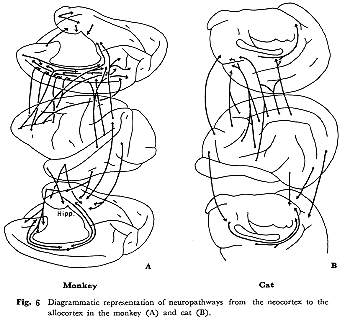
"古い"皮質についての区分の考察は他書にゆずる47)。ここでは脳梁溝と帯状溝との間を占める帯状回皮質を中古皮質(mesocortex),その他を古皮質(allocortexまたはpaleocortexとarchicortex)と呼ぶことにする。新皮質から"古い"皮質への連合線維投射の特徴として,サル,ネコともにそのほとんどすべてが連合野から起こる。まずサル(図6A)では,前頭前野(FD野)のおそらく全域からかなりの量の線維が起こり帯状回の広い範囲に終わる57,59,60)。また,前頭葉極(9野)から脳梁膨大付近の帯状回にいたる長い連合線維もみられる
33)。PG野やTA野の(おそらく)後半域から58)も同様の投射がある。しかしながら,PE(5)野からの投射はみられない。一方側頭連合野の前方域から海馬(広義の海馬形成)付近の古皮質へ連合線維を送る33,59,68)。さらにFink-Heimer法16)で詳しく調べたVan
Hoesenと彼の共同研究者ら75~78)により,最近,腹側側頭皮質,前頭葉眼窩面皮質,帯状回から内嗅領(entorhinal,28野)や周嗅領(perirhinal,35野)に,さらに,内嗅,周嗅両皮質から海馬領野の一定領域に投射があるという確実な証拠が得られた。
ネコ(図6B)の"古い"皮質への投射パターンも基本においてサルのそれに類似している。すなわち,側頭連合野から内嗅領およびその付近の皮質領域へ14,39,41),前頭前野から前辺縁領野(anterior
limbic area, La 56))を介して支脚前野(presubiculum)へ線維を送り,さらに帯状回からはpresubiculumと内嗅野に投射がみられる14)。サルと著しく異なる点は,ネコの帯状回は連合線維をほとんど頭頂連合野からのみうけており,そのほかLa域がわずかに前頭前野から線維をうけるが,側頭連合野からの直接投射は認められない14,40)。
このように,ネコの段階では主たる海馬への入力は頭頂連合野→帯状回→内嗅野・周嗅野→海馬という経路をとり,それに一部側頭連合野が海馬付近へ直接影響を与えるという形態学的構成をみるが,サルの段階になると,前頭葉と側頭葉の発達にともない,前頭連合野(主として背側部)から帯状回へ,それに前頭連合野腹側部,側頭連合野前方部,さらには頭頂連合野から直接,内嗅野・周嗅野へという神経路が著しく発達してくることがわかる。"原始"感覚,情動,記憶さらには自律機能に対する調節ないし統御機能の複雑化(高度化)という視点に立った行動生理学的研究が望まれる。
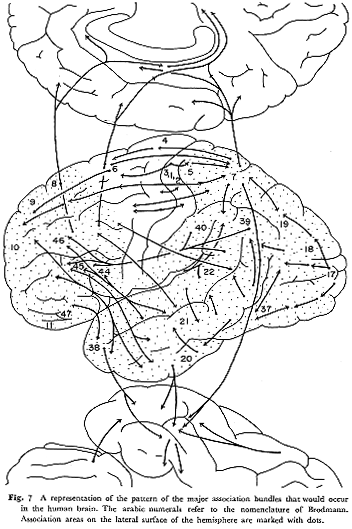
ここで,皮質間結合の主たる特徴について二,三整理しておく。まずサル32,33,49,58,60)およびネコ15,31,39,41,46)において,体性知覚領,聴覚領,視覚領などいわゆる第1感覚野内では同一機能領域内部の相互連絡が,異なる機能領域間の結合にくらべてはるかに強いのに対して,連合野では異なる"連合領野"間の連絡が発達している。すなわち,後連合野(頭頂連合野,側頭連合野,後頭連合野)間の結合をみると,ヒトの下縦束に相当すると思われる神経路がサル,ネコともに一定の発達をとけている。一方,前連合野に関する求心路と遠心路はネコでは未発達であるが,サルの段階になると,ヒトの上縦束,鈎状束,帯状束に相当する長連合神経路はすべて認められる。次に中古皮質への投射はネコでは頭頂連合野からのものに限られるが,サルではさらに前頭前野からの強い投射が加わる。しかし前辺縁領域(La)や内嗅領への連合線維の投射パターンは類似している。最後に種々の感覚情報を伝える連合線維の方向性という点に着目すると,大脳半球内の一定皮質部位に集中する傾向が認められる。すなわち,サルでは上側頭溝をかこむ皮質領域
33),ネコで中S上溝をかこむ皮質領域 41)に各種感覚野からの連合神経路がかなり重複する範囲をもって集中して終わる。この皮質領野についての機能的考察は次項にゆずる。この連合線維の起始ニューロンであるが,サルの"第1次"感覚領野にはみられず,そのほとんどが各種連合野内にあるのに比して,ネコでは,連合野からの成分が比較的少ないようにみえる。一方,前連合野への集中性は後連合野へのそれほどには明瞭に限局していないが,サルにおいてのみ存在する(後述)。
以上,サル(霊長類)とネコ(食肉類)の連合神経路の構成について比較した限りでは,基本においてかなりの共通点がみられる反面,動物の進化の立場から当然予想されるごとく,主として前頭葉の発達にともなう大脳皮質構造の質的変化と考えられる相違点が両者間に存在することが理解される。
はじめに述べたように,ヒトの連合線維に関する確かなデータをわれわれはもっていない。それゆえ,ここに,ネコとサルの脳で明らかにされた共通点と相違点の上に立ち,ストリヒニン法により得られたチンパンジー脳の所見結果3,4,5)をも勘案し,それに多少の推理力も働かせて,ヒトの脳に存在すると思われる連合神経路を推量してみた。サルとヒトを直接結びつけてすぐ比較類推してしまうのは正しくないが,ヒトがサルから進化したことが確かである以上,誤ちを犯す危険は大きいがサルの研究者の態度としてヒト(人間)のことを考えたいからである。少なくとも現段階ではこれを模式図(図7)として提示することが許され,また一定の意義もあろうかと考える。
これまでの論述から,サルの前連合野と後連合野との間の皮質結合は,基本パターンにおいてヒトのそれに比肩し得るほどに発達していると考えられる。しかし,ネコの脳では前連合野の発達は8野の段階でとまり,9~I2野はrudimentaryである。これに対応して両連合野間の結合も未発達である。より社会的動物と思われるイヌになるとネコにくらべて前頭葉もいくぶん発達してくる44)のでこの動物の前頭前野への求心路の研究はなされる価値があると思う。すでにみたような,高次の感覚受容野である後連合野から前連合野へ向かう連合線維は,量的にも集中度の上からいっても逆方向性のものにくらべて優位であり,線維結合(皮質下結合についての考察は本号の他の論文にゆずる)の上からも,皮質構築学上からも,また化石人類の頭蓋骨を調べた考古学的研究24)からも,ひいては,感覚の複合知覚にもとづく感性的認識から符合ないしシンボルの理解という高次作用にもとづく抽象的(理論的)思惟の形成への発展という認識論の立場からいっても、進化論的にいって後連合野において一定レベルの構造上の質的発展段階を経たのちに前連合野が発達してきたものと推論される。この変化を起こす"trigger"は何であろうが?ここで動物の行動様式の発展について考えてみる。外部環境との接触が複雑化してくるなかで,高等な動物は生物学的に高度な発展(達)をなしとげる(直立歩行,前肢による道具使用,発声器官の変化よる分節音の発声可能など)。自然界に対して受動的行動様式を主としてとる段階から能動的行動様式をとることが可能となる過程を経つつサル,さらにはヒトに進化するという観点からみれば,外界からの種々の感覚刺激の入力経路の様式をみることは興昧のある重要な問題である。
ここで大脳辺縁系の近傍に受容野があるとされている嗅覚系と味覚系を除き,体性知覚,視覚,聴覚の大脳皮質内の連合線維による刺激伝達の系について考察してみる。
これらの神経路はそれぞれ体性知覚領(PC;3,1,2)→PE(5)→PG(7)→後連合野;視覚領(OC;17)→OB(18)→OA(19)→後連合野;聴覚領(TC;AI)→AII+TA(22)→後連合野という図式で示されるように段階的に進行する(stepwise
progression 29,33))。サルにおいては,この後連合野内の特定の領域,すなわち上側頭溝周囲皮質(STs域)に上記の3感覚領からの入力が一定のパターンを示して集中している33)。一方ネコでもこれら3感覚領からの中S上溝周囲皮質(MSs域)への集中性の事実が明らかにされた41~43)。さらに,私はサルのSTs域とネコのMSs域がともに5,7野の腹側,19,21野の前方部,22野の後背部に位置するという皮質内の関係をも考慮して両動物間の後連合野内にみられるこの特殊な部位の相同性(homologue)を推論した41)。ヒトの場合39野(角回)と40野(縁上回)とがこの部位に位置することから感覚性言語中枢域(Wernicke中枢)に発達し分化する可能性がこれらの溝をかこむ皮質部分に内蔵されていると考えられる33,41,42)。
この段階の完成が進むなかで,感性的知覚から簡単な弁別学習の経過をへて符合認知への基盤が形成されるのであろうが,さらに高次の符合(言語)を媒介として抽象的思考への段階に進むためには,より発達したより強力な前頭連合野(前頭前野)への入力(形態学的表現でいえば刺激の伝達経路)の存在が必要条件となる。この条件がみたされた時に不断の社会的接触と労働を通じて脳が全体的に発展していく(運動野,感覚野,後連合野の活動で皮質ニューロンは相互に影響を与えつつ機能的に質的発展をとげる)。この過程のなかで前頭前野内に能動的性質をもった言語に関連する領野が新しく形成されてくると考えられる。この段階になってはじめて概念を形成し判断を下し,推理を試みるという思考能力を備えた前頭葉皮質が高度に発達してくる。そのためには形態学的に,少なくとも2種の神経路を介しての前頭葉"刺激"促進要因が考えられよう。
第一の経路は広範囲の大脳皮質領域から視床の諸核や,上丘17,48)へ,さらにニューロンを代えて間脳の後外側核一枕核群(LP-pulvinar
complex)および背内側核(MD)6,18,54)へ,さらに,視床皮質投射1,12,19,20,30,53,71,72~74)を介して,皮質連合野へ影響を与える。このうち,上丘からMD核への投射はネコ18),サル6)において量的に少ないが,たとえば後核群や髄板内核群を介するMD核への入力の構成パターンが将来高等な動物で明らかにされるならば,われわれの前頭前野の機能についての理解はより一層深まると思われる。視床核と皮質連合野との関係は,ある意味では,近年皮質間結合以上に注目されつつあり,私は本号の他の執筆者により書かれる論文に期待している。
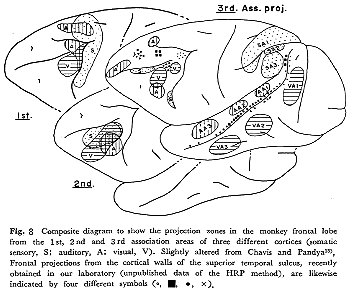
第二の経路は連合線維を介する皮質間レベルの結合で,前頭前野への求心路についてわれわれが得ている1976年までの知見は,すでに考察され図2に示されている。現在われわれはサルの前頭前野の種々の領域にHRPを注入し,PG野,TE野,STs域などの一定部位の皮質内に起始細胞を同定することにより,連合神経路の研究を進め一定の結果を得ている(未発表)。また最近Chavis
and Pandya 13)はFink-Heimer法16)を用いて,サルの後連合野の外側面皮質に限局した傷を与え,前頭前野への求心路について詳しく研究し,従来の所見結果の分析をさらに深めた。彼らは三つの感覚系の段階的刺激進行過程(stepwise
outward progression)の理論の上に立ち,多少の飛躍はあるが,視覚,聴覚,体性知覚系のそれぞれ第1,第2,第3の感覚連合野(sensory
association areas:VA1,2,3;AA1,2,3;SA1,2,3)から起こり前頭前野に終わる連合線維の領域に一定のパターンが存在することを示した。それによると第1連合野(VA1,AA1,SA1)からの連合線維の終止域は弓状溝の前,後の領域で部位的差は認められるが,おのおのがかなり重複する。第2連合野(VA2,AA2,SA2)からの終止域は,主溝腹側部内で,顕著に重複する。これに対して,第3連合野(VA3,AA3,SA3)からの終止域はFD野内の一定部位にそれぞれ限局し互いに重複しない。このような特徴ある投射パターンのもつ意味を解釈することは現在困難であるにしても注目に価する。またSTs域の広い範囲から前頭前野への投射が存左するが(われわれの未発表HRPデータ),その一部を加え,Chavis
and Pandya 13)のFigs.6 and 7を改変して図8に示した。
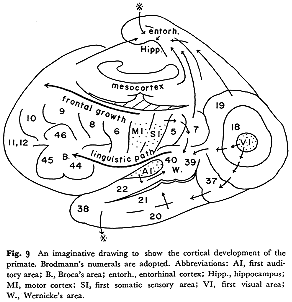
大脳皮質の"階層性"とその発達過程について論ずるには,少なくとも実験発生学的研究や生理学的研究の成果をも加味して全体的立場に立たねばならない。自然界に起こるある現象を一つ下の階層の言葉で表現することをその階層の法則に還元するというが,この視点に立って生物学における物質の最高の発展段階の集合体である大脳皮質の領域で正面から"階層性"の問題に取り組むことは筆者の現在の能力をはるかに越えている。将来の課題として持ち続けていきたいと思う。ここに,皮質連合野の段階的発展(達)を示唆する抽象図を図9として掲げ擱筆する。
図の作製,原稿の清書に御協力下さった当教室の菅原主三,北村陽子,山崎義孝,内藤順平の諸氏に,また,拙稿をご一読下さった山内昭雄,車田正男の両教授に感謝いたします。
文献
1) Akert, K.: Comparative anatomy of frontal cortex and thalamocortical connections. The
frontal granular cortex and behavior (ed. by J.M. Warren and K. Akert). 372-396,
McGraw-Hill, New York, 1964.
2) Amassian, V.E.: Studies on organization of a somesthetic association area, including a
single unit analysis. J. Neurophysiol., 17: 39-58, 1954.
3) Bailey, P., Barenne, J.G., Garol, H.W. and McCulloch, W.S.: Sensory cortex of
chimpanzee. J. Neurophysiol., 3: 469-485, 1940.
4) Bailey, P., Bonin, G.v., Garol, H.W. and McCulloch, W.S.: Long association fibers in
cerebral hemispheres of monkey and chimpanzee. J. Neurophysiol., 6: 129-134, 1943a.
5) Bailey, P., Bonin. G.v., Garol, H.W. and McCulloch, W.S.: Functional organization of
temporal lobe of monkey (Macaca mulatta) and chimpanzee (Pan satyrus). J. Neurophysiol.,
6: 121-128, 1943b.
6) Benevento, L.A. and Fallon, J.H.: The ascending projections of the superior colliculus
in the rhesus monkey (Macaca mulatta). J. comp. Neur., 160: 339-362, 1975.
7) Blake, L., Jarvis, C.D. and Mishkin, M. : Pattern discrimination thresholds after
partial inferior temporal or lateral striate lesions in monkeys. Brain Res., 120: 209-220,
1977.
8) Bonin. G.v, and Bailey, P. : Neocortex of Macaca mulatta, Univ. Illinois Press, Urbana,
1947.
9) Brodmann, K.: Beitrage zur histologischen Lokalisation der Grosshirnrinde. V. Ueber den
allgemeinen Bauplan des Cortex pallii bei den Mammaliern und zwei homologe Rindenfelder im
besonderen. Zugleich ein Beitrag zur Furchenlehre. J. f. Psychol. u. Neurol., 6 : 275-400,
1906.
10) Brodmann, K.: Beitrage zur histologischen Localisation der Grosshirnrinde. VI. Die
Cortexgliederung des Menschen. J. f. Psychol. u. Neurol., 10: 231-246, 1908.
11) Brodmann, K. : Vergleichende Localisations-lehre der Grosshirnrinde. Barth, Lepzig,
1909.
12) Burton, H. and Jones, E.G. : The posterior thalamic region and its cortical projection
in new world and old world monkeys. J. comp. Neur., 168: 249-302, 1976.
13) Chavis, D.A. and Pandya, D.N.: Further observations on corticofrontal connections in
the rhesus monkey. Brain Res., 117: 369-386,
1976.
14) Cragg, B.G.: Afferent connexions of the allocortex. J. Anat. (Lond.), 99: 339-357,
1965.
15) Diamond, I.T., Jones, E.G. and Powell, T. P.S.: The association connections of the
auditory cortex of the cat. Brain Res., 11 : 560-579, 1968.
16) Fink, R.P. and Heimer, L.: Two methods for selective silver impregnation of
degenerating axons and their synaptic endings in the central nervous system. Brain Res., 4
: 369-374, 1967.
l7) Garey, L.J., Jones. E.G. and Powell, T.P.S.: Interrelationships of striate and
extrastriate cortex with the primary relay sites of the visual pathway. J. Neurosurg.
Psychiat., 31 :135-157, 1968.
18) Graham, J.: An autoradiographic study of the efferent connections of the superior
colliculus in the cat. J. comp. Neur., 173: 629-654, 1977.
l9) Graybiel, A.M.: Some ascending connections of the pulvinar and nucleus lateralis
posterior of the thalamus in the cat. Brain Res., 44: 99-125, 1972.
20) Graybiel. A.M.: The thalamo-cortical projection of the so-called posterior nuclear
group : A study with anterograde degeneration methods in the cat. Brain Res., 49 :
229-244, 1973.
21) Gurewitsch, M. and Chatschaturian, A.: Zur Cytoarchitektonik der Groβhirnrinde der
Feliden. Z. Anat. Entwickl.-Gesch., 87: 100-138, 1928.
22) Hassler, R. and Muhs-Clement, K. : Architektonischer Aufbau des sensomotorischen und
parietalen Cortex der Katze. J. Hirnforsch., 6: 377-422, 1964.
23) Heath, C.J. and Jones, E.G. : The anatomical organization of the suprasylvian gyrus of
the cat. Ergebn. Anat. Entwickl.-Gesch., 45: 64 pp., 1971.
24) Holloway, R.S.: The casts of fossil hominid brains. Sci. Amer., 231: 106-115., 1974.
25) Hubel, D.H. and Wiesel, T.N.: Receptive fields and functional architecture in two
nonstriate visual areas (18 and 19) of the cat. J. Neurophysiol., 28 : 229-289, 1965.
26) Hubel, D.H. and Wiesel. T.N. : Stereoscopic vision in macaque monkey : Cells sensitive
to binocular depth in area 18 of the macaque monkey cortex. Nature (Lond.), 225: 41-42,
1970.
27) lwai, E. and Mishkin, M.: Further evidence on the locus of the visual area in the
temporal lobe of the monkey. Exp. Neurol., 25: 585-594, 1969.
28) Jacobsen, C.F.: Function of the frontal association area in primates. Arch. Neurol.
Psychiat., 33: 558-569, 1935.
29) Jones, E.G. : The anatomy of extrageniculostriate visual mechanisms. In: The
Neurosciences, third study program (eds. Schmitt, F. O. and Worden, F.G.), p. 215-227, MIT
Press, Cambridge, Mass., 1974.
30) Jones, E.G. and Burton, H.: Areal differences in the laminar distribution of thalamic
afferents in cortical fields of the insular, parietal and temporal regions of primates. J.
comp. Neur., 168: 197-248, 1976.
31) Jones, E.G. and Powell, T.P.S. : The ipsilateral cortical connexions of the somatic
sensory areas in the cat. Brain Res., 9: 71-94, 1968.
32) Jones, E.G. and Powell, T.P.S.: Connexions of the somatic sensory cortex of the rhesus
monkey. I. Ipsilateral cortical connexions. Brain, 92: 477-502, 1969.
33) Jones, E.G. and Powell, T.P.S.: An anatomical study of converging sensory pathways
within the cerebral cortex of the monkey. Brain, 93: 793-820, 1970.
34) Kawamura, K.: Association and commissural fibers from the sensorimotor and adjacent
cortices in the cat. Brain and Nerve, 18: 1003-l012, 1966.
35) Kawamura, K.: Association and commissural fibers from the second somatosensory (SII)
cortical area in the cat. Acta anat. nippon., 44: Suppl. 2, 4 p, 1969.
36) Kawamura, K.: Association and commissural fibers from the auditory and adjacent
cortices in the cat. Acta anat. nippon., 45: 57-58, 1970a.
37) Kawamura, K.: Association and commissural fibers in the cat cerebrum. IX Int. Congr.
Anatomists, Abstracts of Papers, 63 : 1970b.
38) Kawamura, K.: Variation of the cerebral sulci in the cat. Acta anat., 80: 204-221,
1971.
39) Kawamura, K.: Corticocortical fiber connections of the cat cerebrum. I. The temporal
region. Brain Res., 51 : l-21, 1973a.
40) Kawamura, K.: Corticocortical fiber connections of the cat cerebrum. II. The parietal
region. Brain Res., 51 : 23-40, 1973b.
41) Kawamura, K.: Corticocortical fiber connections of the cat cerebrum. III. The
occipital region. Brain Res., 51 : 41-60, 1973c.
42) Kawamura, K.: Association fibers of the cerebral cortex. A functional consideration of
the cortex based upon morphological findings (Review). J. Iwate Med. Ass., 26: 629-639,
1974.
43) Kawamura, K. and Naito. J.: Corticocortical afferents to the cortex of the middle
suprasylvian sulcus area in the cat. In: Afferent and Intrinsic Organization of Laminated
Structures in the Brain, ed. by O. Creutzfeldt., Exp. Brain Res., Suppl. 1, Springer,
Berlin, p. 323-328, 1976.
44) Kawamura, K. and Naito, J.: Variations of the dog cerebral sulci, compared in
particular with those of the cat. J. Hirnforsch., in press.
45) Kawamura, K. and Otani, K.: Association and commissural fibers from the association,
visual and adjacent cortices in the cat. Acta anat. nippon., 41 : Suppl. 6, 8 p, 1966.
46) Kawamura, K. and Otani, K. : Corticocortical fiber connections in the cat cerebrum :
The frontal region. J. comp. Neur., 1 39 : 423-448, 1970.
47) 小池上春芳:大脳辺縁系.第2版、中外医学社、東京、384頁、1968.
48)紺野敏昭、川村光毅、内藤順平:ネコ皮質視蓋路の起始細胞とその終止部位について。Fink-Heimer法およびHRP法による実験解剖学的研究.解剖誌.,
51: 261, 1976.
49) Kuypers, H. G. J. M., Szwarcbart, M. K., Mishkin, M. and Rosvold, H.E.:
Occipito-temporal corticocortical connections in the rhesus monkey. Exp. Neurol., 1 1 :
245-262, 1965.
50) Luria, A.R. : Frontal lobe syndromes. In Handbook Clin. Neurol., 2, Localization on
clinical neurology (ed. Vinken, P.J. and Bruyn, G.W.), p. 725-757, North-Holland,
Amsterdam, 1969.
51) Nauta, W. J.H. : Silver impregnation of degenerating axons. In W.F. Windle (Ed.), New
Research Techniques of Neuroanatomy. Thomas, Springfield, Ill., p. 17-26, 1957.
52) Nauta, W. J.H. and Gygax, P.A.: Silver impregnation of degenerating axons in the
central nervous system : A modified technic. Stain Technol., 29: 91-93, 1954.
53) Niimi, K., Kadota. M. and Matsushita, Y.: Cortical projections of the pulvinar nuclear
group of the thalamus in the cat. Brain Behav. Evol., 9 : 422-457, 1974.
54) Niimi, K., Miki, M. and Kawamura, S.: Ascending projections of the superior colliculus
in the cat. Okajimas Folia Anat. Jpn., 47: 269-287, 1970.
55) Otsuka, R. and Hassler, R.: Uber Aufbau und Gliederung der corticalen Sehsphare bei
der Katze. Arch. Psychiatr., 203: 212-234. 1962.
56) Papez, J. W.: Comparative Neurology. A Manual and Text for the Study of the Nervous
System of Vertebrates. Noble Offset Printers, New York, 1929.
57) Pandya, D.N., Dye, P. and Butters, N.: Efferent corticocortical projections of the
prefrontal cortex in the rhesus monkey. Brain Res., 31 : 35-46, 1971.
58) Pandya, D.N., Hallett, M. and Mukherjee, S.K. : Intra- and interhemispheric
connections of the neocortical auditory system in the rhesus monkey. Brain Res., 14:
49-65, 1969.
59) Pandya, D.N. and Kuypers, H.G.J.M.: Cortico-cortical connections in the rhesus monkey.
Brain Res., 13: 13-36, 1969.
60) Pandya, D.N. and Vignolo, L.A.: Intra- and interhemispheric projections of the
precentral, premotor and arcuate areas in the rhesus monkey. Brain Res., 26: 217-233,
1971.
61) Petr, R., Holden, L.B. and Jirout. J.: The efferent intercortical connections of the
superficial cortex of temporal lobe (Macaca mulatta). J. Neuropathol. Exp. Neurol., 8 :
100-103, 1949.
62) Robertson, R.T.: Thalamic projections to visually responsive regions of parietal
cortex. Brain Res. Bulletin, 1 : 459-469, 1976.
63) Robertson, R.T.: Thalamic projections to parietal cortex. Brain Behav. Evol., 14:
161-184, 1977,
64) Robertson, R.T., Mayers, K.S., Teyler, T. J., Rettinger, L.A., Birch, H., Davis, J.L.,
Phillips, D.S. and Thompson, R.F.: Unit activity in posterior association cortex of cat.
J. Neurophysiol., 38 : 780-794, 1975.
65) Rose, J.E. and Woolsey, C.N.: Structure and relations of limbic cortex and anterior
thalamic nuclei in rabbit and cat. J. comp. Neur., 89 : 279-340, 1948.
66) Rosenkilde, C .E. and Divac, I . : Discrimination of time intervals in cats. Acta
Neurobiol. Exp., 36: 311-317, 1976.
67) Sanides, F. and Hoffmann, J.: Cyto- and myeloarchitecture of the visual cortex of the
cat and of the surrounding integration cortices. J. Hirnforsch., 11 : 79-104, 1969.
68) Seltzer. B. and Pandya, D.N.: Some cortical projections to the parahippocampal area in
the rhesus monkey. Exp. Neurol., 50 : 146-160, 1976.
69) Sprague, J.M., Levy, J., DiBerardino. A. and Berlucchi, G.: Visual cortical areas
mediating form discrimination in the cat. J. comp. Neur., 172: 441-488, 1977.
70) Thompson, R.F., Johnson, R.H. and Hoopes, J.J.: Organization of auditory, somatic
sensory and visual projection to association fields of cerebral cortex in the cat. J.
Neurophysiol., 26 : 343-364, 1963.
71) Tobias, T. J.: Afferents to prefrontal cortex from the thalamic mediodorsal nucleus in
the rhesus monkey. Brain Res., 83: 191-212, 1975.
72) Trojanowski, J.Q. and Jacobson, S.: Medial pulvinar afferents to frontal eye fields in
rhesus monkey demonstrated by horseradish peroxidase. Brain Res., 80 : 395-411, 1974.
73) Trojanowski, J.Q. and Jacobson, S.: A combined horseradish peroxidase-autoradiographic
investigation of reciprocal connections between superior temporal gyrus and pulvinar in
squirrel monkey. Brain Res., 85 : 347-353, 1975.
74) Trojanowski, J.Q. and Jacobson, S.: Areal and laminar distribution of some pulvinar
cortical efferents in rhesus monkey. J. comp. Neur., 169: 371-392, 1976.
75) Van Hoesen, G.W., Pandya, D.N. and Butters, N.: Cortical afferents to the entorhinal
cortex of the rhesus monkey. Sciencc, 175: 1471-1473, 1972.
76) Van Hoesen, G.W. and Pandya, D.N. : Some connections of the entorhinal (area 28) and
perirhinal (area 35) cortices of the rhesus monkey. I. Temporal lobe afferents. Brain
Res., 95: 1-24, 1975.
77) Van Hoesen, G.W., Pandya, D. N. and Butters, N.: Some connections of the entorhinal
(area 28) and perirhinal (area 35) cortices of the rhesus monkey. II. Frontal lobe
afferents. Brain Res., 95: 25-38, 1975.
78) Van Hoesen, G.W. and Pandya, D.N.: Some connections of the entorhinal (area 28) and
perirhinal (area 35) cortices of the rhesus monkey. III. Efferent connections. Brain Res.,
95: 35-59, 1975.
79) Walker, A.E.: A cytoarchitectural study of thc prefrontal area of the macaque monkey.
J. comp. Neur., 73: 59-86, 1940.
80) Wester, K.G., Irvine, D.R.F. and Thompson, R.F. : Acoustic tuning of single cells in
middle suprasylvian cortex of cat. Brain Res., 76 : 493-502, 1974.
81) Woody, C.D., Knispel, J.D., Crow, T.J. and Black-Cleworth, P.A. : Activity and
excitability to electrical current of cortical auditory receptive neurons of awake cats as
affected by stimulus association. J. Neurophysiol., 39: 1045-1061, 1976.
82) Woolsey, C.N. : Patterns of sensory representation of the cerebral cortex. Fed. Proc.,
6: 437-441, 1947.
83) Woolsey, C.N.: Organization of cortical auditory system: A review and a synthesis. In:
Neural mechanisms of the auditory and vestibular systems. eds. by G.L. Rassmussen and W.
Windle, Thomas, Springfield, Illinois, p. 165-180, 1960.
84) Zeki, S.M.: Functional organization of a visual area in the posterior bank of the
superior temporal sulcus of the rhesus monkey. J. Physiol., 236 : 549-573, 1974a.
85) Zeki, S.M.: Cells responding to changing image size and disparity in the cortex of the
rhesus monkey. J. Physiol., 242: 827-841, 1974b.
神経研究の進歩 21/6(1977)1085-1101(医学書院) 許可を得て転載