物理的信号としての音は空気や水のような媒質を伝わる粗密波/縦波である。空気が或る振動数の範囲内で、かつ、或る振幅の範囲内で振動すると、われわれはそれを音として感じる。すなわち、聴覚は音波が空気の微弱振動に基づいて、内耳の聴覚器官に伝わり感覚細胞を刺激し、脳中枢が反応して作り出す感覚である。電気エネルギーをもった刺激が、蝸牛神経→蝸牛神経核→上オリーブ複合体→外側毛帯核→下丘→内側膝状体→第一聴覚皮質へと伝達される。ヒトの可聴域(可聴範囲)は20Hz-20kHzで、この範囲での音(響き)を可聴音という。自然界に存在する超低周波音や超音波はヒトでは少なくとも可聴音としては知覚されない。
単一の周波数からなる音は純音とよばれ、最も基本的な音で、正弦波で表わされるが、現実に耳にする音は複数の純音に分解される複合音である。複合音を複数の純音の成分に分解して調べることを周波数分析という。会話や歌で使われている音声は複合音で、複数の周波数を含み、時間的に複雑な変化をする音である。
楽器の音も複合音で、その波形には基音と倍音がある。楽器の音の成分中、最も周波数が低く強い成分(基音)の整数倍の周波数成分が強いのが普通である。こうした周波数が基音の整数倍である成分を倍音と言う。倍音が含まれた複合音は音色(音質)が強く意識されるようになる。音色は振動の様相によって決まる。それには、波形や振幅の時間的変化が複雑に関係する。波形は通常、倍音の含まれ方によって決まる。
いわゆる楽音は純音ではないが、規則性のある振動が持続するもので、複数の倍音を含んでいる。一般の西洋楽器の演奏に用いられる諸楽器の発する音のように、一定の周波数をもち、部分音(基音と倍音)が基音の整数倍となるような安定した音である。その音が倍音を含む度合いによって、異なった音色に感じる。つまり、基音が同じ周波数でも、2倍音や3倍音の含まれる割合が違うと異なった音色に感じる。楽音の性質は、高さ(振動数)・強さ(振動エネルギーなど)・音質/音色(波形など)の3要素によって決まる。
次に音声であるが、これは時間的には安定した音ではない。例えば、「ta」という音声は、子音「t」に続いて母音「a」が鳴る。音声の周波数を分析するソナグラムはスペクトルの時間的変動を調べるもので、時間を横軸に周波数を縦軸にとり、周波数成分のパワーを濃淡で表すようにした三次元グラフである。有声母音のソナグラムは、いくつかの周波数帯にエネルギーが集中していることが特徴である。これは声帯から口腔まで呼気道の共振(または共鳴)の特性を反映している。この声道内の共振周波数をフォルマント周波数という。その低いほうから第一、第二、第三・・・フォルマントと呼ぶ。これは、声帯音が声道を通過する間に共鳴腔に共鳴して、それらの共鳴周波数帯にエネルギーが集中するために生じるもので、声帯の振動数から特定の周波数の音を選択する過程は、管楽器で色々の音を出す仕組みと原理的に同じである。ちなみに、第一と第二のホルマント周波数によって音声母音の性質/タイプが決定される。
無声(子)音には周波数分析上明瞭なピークがなく、広い周波数範囲に連続的に分布するスペクトルになっている。「s」や「h」などの摩擦子音は、声道の一部の狭い部分を呼気が通り抜けるときの雑音が音源となっている。有声母音の音源である声帯音のスペクトルが、基音と倍音が等間隔に並ぶ成分からなるのに反して、摩擦音のスペクトルでは分離した成分がみられず、ある周波数帯に連続的にエネルギーが分布している。このほか、破裂音、鼻音、半母音なども、それぞれ独特なスペクトルパターンをもっている。
乳幼児の発声は「ア∼」や「ウ∼」という、笑ったり泣いたりするときに発する母音を以って始まる。子音を発声せしめるためには、その発生機構の発達が必要であるし、言葉として巧みに連続的に発語するためには、前頭葉皮質を含む脳が全体として活動していなければならない。すなわち、音声を作る共鳴腔を使って、母音と子音とを組み合わせて音声をつくるためには、この空間の壁をつくる筋肉(喉頭、咽頭、口蓋、口腔を形成する構音筋や表情筋など)とその支配神経(迷走神経、舌咽神経、顔面神経、三叉神経)系統の発達を要する。系統発生学的にみると、鰓呼吸をする魚類から肺呼吸をする陸棲動物に進化する過程で、魚の顔面皮膚に開いていた水の出口と入り口の穴が管状につながり(この中に鼻の原基に相当する嗅覚器がある)、さらに口腔の天井と連続して鼻腔と咽頭が交通する。哺乳類では、両生類で形成され始めた口蓋(一次口蓋)が二次口蓋として完成されて、鼻腔は口に開くのではなく咽頭―気管―気管枝―肺とつらなる気道を形成する。このとき気管の先端の喉頭が上に突き出て鼻腔後方の出口にはまりこみ、その結果、鼻から真っ直ぐに喉頭-気管へと流れる「気道」および口から気道の横を通り「食道」へとつづく通路が分離する。こうして両道の立体交差の形が完成する。ヒトになると、喉頭の位置が低くなり、鼻腔と喉頭が分離され、鼻からでも口からでも容易に呼吸し得て、声帯をもつ喉頭から口に空気を出すことができるようになる。他の動物と違って、ヒトでは喉頭蓋の開閉が可能になったために、気道と食道とは咽頭で立体交差をするのではなく、点滅信号機付きの平面交差をするようになる。そして、声帯で作られた空気の振動を複雑な形に変化させ得る口腔という付属共鳴腔内で子音を含む複雑な音を加工し、発語できるようになった。これはヒトの発声にみられる際立った特徴であり、子音発声機構の発達は言語の発生と深く関係する重要な注目すべき点である。
ところで、上述のような複雑な物理的信号の性質をもったヒトの音声や楽音が、脳内でどのように分析されてヒトは言語を理解し、音楽を聴受することが出来るのであろうか。
こうした考察に入る前に、これまでの前置きが必要であった。
音の認知には3つのレベルがある。すなわち、
a)物理的レベル:物理的現象としての音であって、われわれの知覚作用とは独立して存在する。
b)生理的レベル:鼓膜によって捉えられた空気の振動は神経信号に変換されて脳に伝達される。感覚受容器内や脳内の伝達過程で信号としての音の性質が変形・加工され、認知される。
c)“精神”的レベル:脳が音の神経信号を認識する。未解明の部分が多いが、この段階でさまざまな情報処理が行われて、はじめて意味のある音や声として認識される。
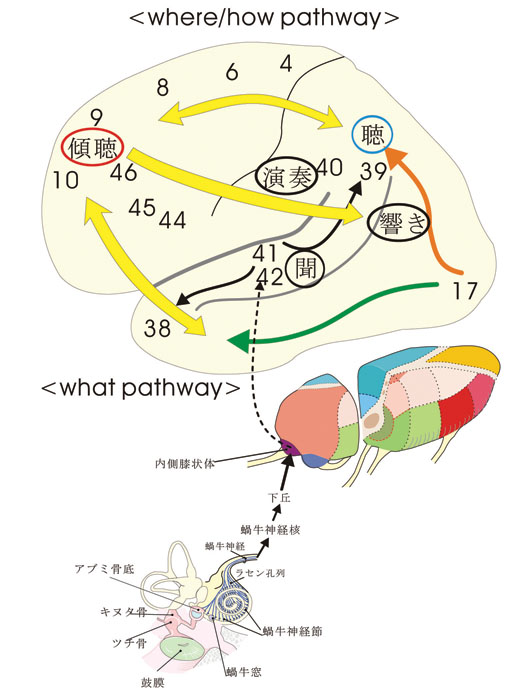
この小論の主題はb)とc)のレベルである。以下に、1)音を聞く、聞こえる, 2)調べ(旋律)・言葉を聴く、 3)音楽・講演を傾聴する、4)楽器を奏でる・歌を歌う、の順に考察する。図1を参照されたい。
外耳道底にある鼓膜を振動させた音の縦波は、鼓室内の耳小骨を、ついで骨迷路内の液体を振動させる。さらに、骨迷路内にある膜迷路の中の内リンパ液に伝わり、膜迷路(蝸牛)の有毛感覚細胞を刺激する。
水棲動物には、水の振動を知覚し液体の流れやそこに含まれる化学物質を感じとる側線とよばれる感覚器官がある。側線器は身体の側面だけでなく、頭部皮下にも複雑な管系を作っており、そこに有毛感覚細胞群が並んでいる。これによって、水圧、水流、低周波振動などが受容され、振動源の方向と距離が認知される。遠隔(teleceptive)受容器としての原始的なもので、第8脳神経が司る聴覚・振動を受容する感覚器官(いわゆる耳)は側線器が分化したものと考えられる。
哺乳動物の聴覚の伝導路についてみると、刺激(興奮)が視床皮質路を通り第一皮質聴覚野(A1)に到達する。蝸牛内での周波数の場所表示(周波数軸)は、少なくともA1までは保存されている。聴覚系には末梢から中枢のA1まで特定の周波数によく反応する部分が順序よく並んでいる。内耳における蝸牛基底膜上の周波数の場所表示はその基部が高周波音、先端部が低周波音に対応している。
蝸牛核は前腹側核、後腹側核、背側核からなり、それぞれの亜核で低周波領域は腹側に、高周波領域は背側に位置する。反応にみられる性質から、前腹側核のニューロンは情報を上位に伝達するという単純な機能をもち、背側核はより複雑な信号分析に関与していると考えられている。
上オリーブ複合体もいくつかの亜核に分けられている。そしておのおの周波数の場所表示は保たれている。外側核には高周波音によく反応するニューロンが多く、内側核には低周波音に反応するニューロンが多い。
外側毛帯核も腹側核と背側核に分けられ、腹側核は音響驚愕反応に関与する。背側核は両側の下丘に線維を送っており、その多くは抑制性神経伝達物質GABAを含んでいる。
下丘は脳幹で最大の聴覚系の細胞集団である。中心核、外核、周囲核と区分されるが、中心核が最大で聴覚伝導路の中継核である。中心核では同一の周波数に同調したニューロンが面状に並び、同心円状に層構造の配列をしている。そして低周波音に応ずるニューロンは背側に、高周波音に応ずるニューロンは腹側に位置している。また、中心核には抑制性神経伝達物質GABAが含まれ、周波数同調を先鋭化する構造となっている。このように、中枢神経系では抑制作用の機構によって周波数同調、周波数弁別能が高められている。また、特定の時間情報に反応するニューロンもある。
内側膝状体は下丘から同側性に入力をうけている。腹側核、背側核、内側核に区分され、腹側核のニューロンは下丘中心核から主たる入力をうけてA1に投射している(視床皮質路)。なお、腹側核は周波数ごとの層構造を示す。
第一聴覚皮質(A1)は内側膝状体腹側核から主たる入力をうける。閾値付近の刺激レベルでは周波数の場所表示が存在する。
末梢レベルの蝸牛神経では音の時間パターンはニューロンの発火パターンの時間変化によって表わされるが、中枢ではこの時間パターンを統合した時間情報として抽出される。われわれは、音声や音楽の時間的に変化する音を聞いたとき、刻々と変化する響きの高さや強度、あるいは倍音などを追って、その時間パターンの違いによって音のもつ意味を知覚している。
ここで、周波数分析について述べておく。内耳にある蝸牛殻基底膜の振動様式をみると、純音刺激で基底膜のかなり広い範囲が振動する。最大振幅を示す位置は、高音のときはアブミ骨側に近く、低音のときは殻頂側に近く、周波数にしたがって規則正しく並んでいる。しかし、最大振幅点は特定の周波数にだけ共振するわけではなく、他の周波数の音に対しても小さな振幅をもって共振している。このように、内耳レベルでは周波数分析は未完成の状態である。この点、脳の一部として発生した網膜が、内部の神経回路網により、かなり高度に情報処理された信号を外側膝状体に送っているのに対して、聴覚系では前述のように、内側膝状体に信号が到達するまでには中脳以下の多くの神経核を経過することによる情報処理が必要とされることは注目に値する(後述参照)。
微小電極法を用いて、聴覚系の単一ニューロン活動を調べ、それぞれの周波数における閾値を測定した結果、横軸に周波数を、縦軸に音圧(音の強さ)を二次元座標上にプロットしてそのニューロンの閾値曲線を作成する。閾値曲線の内側に相当する音にそのニューロンは反応するので、この範囲をそのニューロンの応答野という。その結果をみると、聴覚伝達系における上位の核ほど個々のニューロンの応答野が鋭くなり、下丘で聴ニューロンの応答周波数範囲が最も狭い。聴覚系の周波数分析は下丘レベルで完成していると言う。
これに対して、内側膝状体と聴覚皮質(A1)では、スペクトルパターンの弁別が行われている。無麻酔ネコでの実験で、内側膝状体の大半のニューロンは、純音に対して音の始まりのみに発射するon反応、終わりのみに発射するoff反応,両方で発射するon-off反応を示す。応答野は狭いが、多くは多峰性で、下丘のニューロンの応答野よりも広い周波数範囲にわたっている。大脳皮質聴覚野ではこの傾向がよりはっきりしている。これらのことは、内側膝状体以降の主要機能が、周波数分析とは異なったものであることを示している。多峰性の応答野は、下位からの入力が収束していることを示しており、何らかの統合が行われていることを示唆している。
無麻酔動物での微小電極法やヒトでの臨床病理学的研究を参考に皮質聴覚野の機能を調べる。「音声知覚の仕組み」について、音声の弁別は、識別ニューロンによるのか、感受性ニューロンによるのか、丸山(1995)は、無麻酔ネコ聴覚野での実験研究の結果次のように考えている。すなわち、「視覚や聴覚など複雑な内容の知覚における神経機序に対して二つの考えがある。一つは或る知覚に対応する検知ニューロンが存在するというものであり、もう一つは或る知覚には一群の細胞集団の空間的(または時間的)活動パターンが対応するというものである。実験結果から考えて、母音識別ニューロンは検知ニューロンに相当し、母音感受性ニューロンは後者に相当する。すなわち、二つの仕組みが共存することを示している」。ネコとヒトの間に音声弁別(仲間の鳴き声の聞き分け)の仕組みや音声のスペクトルパターンに多くの共通性があることから考えて、両者間には基本的に同様の性質が存在すると考えられる。
興奮が感覚性言語野を含む後連合野内で進行する。音声の識別・認識には多くの時間情報が聴覚中枢で統合処理されるが、そのような神経機構が必要となる。
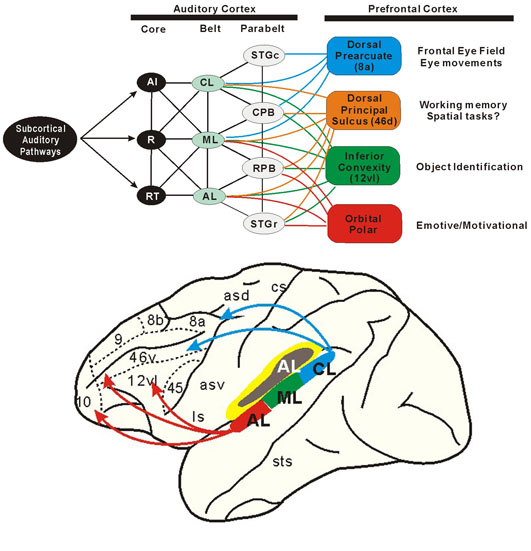
最近サルの聴覚領皮質で以下のことが調べられた(Romanski ら, 1999)。core部分(A1; auditory core area, R; rostral core, RT; rostrotemporal core) は、各振動数に対応した単純な音をうけとる。ついで、coreからmedial & lateral belts (CL, ML, AL) に興奮が伝わり、さらに lateral belt から parabelt へと進行する(図3参照)。この過程を通じて聴覚処理の内容が高められる。第一次聴覚野(A1)から belt へ軸索が伸びている。そして、いくつかの音が合する22野 (belt, parabelt) では、雑音 (white noise) に反応するニューロンが存在する。また、ネコに音を聞かせたときにも、第一および第二皮質聴覚領野にtuningの幅に広狭がみられ、潜伏時間にも長短があるなど種々の性質をもったニューロンの存在が知られている(He ら,1999)。
サルの上側頭葉の最前部は扁桃体との結合もみられることから、感情表現に関連する聴覚領域と思われる。上側頭溝(STS)周囲の皮質域には、後背部は言語系に、前腹部は情動系に、また、中央部を含めて全域的に多種感覚性ニューロンが存在しており、さまざまな感覚の入力がここに集中して終わっている(川村、1973, 1977)。
サルの側頭連合野に属するTG域やTE域前方部は鈎状束とよばれる両方向性の連合線維によって、前頭葉眼窩面皮質(OF)と相互に強く結びつけられている。これらの皮質 (TG,TE,OF) はともに扁桃核と密接に相互結合をもっている(Kawamura と Norita,1980)。つまり、感覚性言語野と結びついた情動表現が、運動性言語野と関わりをもつ能動的な情動行動と結びついている。前頭前野内で「組み替え」処理がなされたものが前補足運動野に伝えられ、意欲や発語(構音運動のプログラミング)などに関与するとされている帯状回皮質内の補足運動野や運動前野に興奮が伝えられる。そして、随意運動を司る一次運動領のニューロンを興奮させ、錐体路系を作働させる。
ヒトの脳では、シルヴィウス溝(外側溝)壁面皮質にHeschl 回と呼ばれる第一聴覚野(A1)があり、その後方に連続して存在する感覚性言語野(ウエルニッケ野)は、A1から多量の線維をうけている。このことから、聴覚野では単音の弁別が行なわれ、しかる後に、言語野で単語の弁別認知が行われると考えられる。また、失語症/失音楽症を示した臨床例を分析してみると、A1に到達した楽器音や音声のシグナルは、この皮質からさらに進むプロセスの中で相互に関連し合いながらも、音楽的要素と言語的要素とに機構上仕分けされるように思われる。
興奮が前連合野(前頭前野)に到達する。ここでは演奏と前頭葉の活動の関係が問題となる。前頭葉はヒトおよび動物の何等かの行為、すなわち能動的活動を司る場で、広義の運動系すなわち自発的、能動性を計画し、命令し、実行させる。4野や大部分の6野の運動系領域へは後連合野からの直接の投射はみられない。後連合野からの主たる入力が終止するのは前頭前野で、いくつかのシナップスを経て、一次運動野に興奮が伝えられる。サルの大脳皮質―線条体結合の仕事をみると、主溝を中心とした領域からは主として尾状核に、その背側(6野, 前頭前野)や眼窩面に近い腹側野からは尾状核と被殻へ投射しており、第一次運動野、体性感覚野を中心とする領野からは主に被殻に投射がある。その他に頭頂葉(尾状核と被殻両者へ)や側頭葉(とくに尾状核へ)からも線条体に投射がある。
後連合野からの刺激は運動性言語野を含む広い範囲の前頭前野に入力する。言語表現能力を備えた領域ないしその近傍域でいわゆる音や形象の「組み替え」がなされる。音の内容が解釈され、創造性の基盤が形成される。フローチャート式に書けば、感覚野→後連合野→前連合野→高次運動野→第一次運動野という図式になる。高次運動野とは、この十数年来 Evertz や丹治らにより研究されてきた領域で、運動前野、補足運動野(内側面6野)などが含まれる(丹治、1999)。
音楽「作品」という形で表されている、人間の内的「表現」は、演奏という「運動」によって成立する能動的表現を通じてもたらされる「外化」と、そこで「外化」され響きとなって伝達されている波動としての「表現」を聴くという受動的表現との二つの面に分けて考えられる。受動的表現については前項で論じたので、ここでは演奏における表現、すなわち何等かの事象を他者伝えようとする意志ないし意欲と運動との関連を含めて考察する。
大脳皮質の一次運動野の興奮が脊髄の運動ニューロンに伝えられ、筋肉を随意的に動かすという古典的「錐体路系」という神経投射路がある。しかし、人はいつも随意的な運動にだけ支えられているわけではない。身体全体の運動系を調和させ、統御し、運動を円滑ならしめる機構が存在するはずである。前脳(終脳と間脳)が未発達の動物にも、環境に働きかける上で必須の基本的なシステムとして能動的作用が備わっている。それはいわゆる「錐体外路系」といわれるシステムで、小脳を中心とした回路、大脳基底核を中心とした回路、脳幹網様体や脊髄を中心とした運動系の回路と言われるものがこれに属する。この「錐体外路系」は動物の運動系の基本的な形態的基盤をなすもので、系統発生的に古いシステムである。
動物の発達・進化に伴い、その脳機能は高次化する。皮質連合野について言えば、後連合野が機能を発揮した後に、動物間のコミュニケーションや協同作業を通じて、環境への能動的働きかけが起こり、前頭葉、とくに運動性言語野(下前頭回後部領域で「構音性の言語の座」と言われ後方に隣接する一次運動野内の構音筋支配域と関連が強い、別名ブローカ野)を含む前頭前野、が発達してくる。サルが環境条件を変えられたとき、自分で新たに適切な課題解決を考え出した時のみ報酬を与えるとういうパラダイムを施行させ、その結果を観察・分析することによって行われる「意欲」に関する研究があるが、その際に特異的に反応するニューロンが帯状回で見いだされている(Shima と Tanji , 1999)。ここは外側面の前頭前野と相互に連絡する領域で意欲に関係する領域と言われている。なお、帯状回や前頭前野には中脳腹側被蓋野(VTA)からのA10 ドーパミン含有線維が多数終わっており、ここは意欲、情操、道徳、運動などに関連する領域とされている。
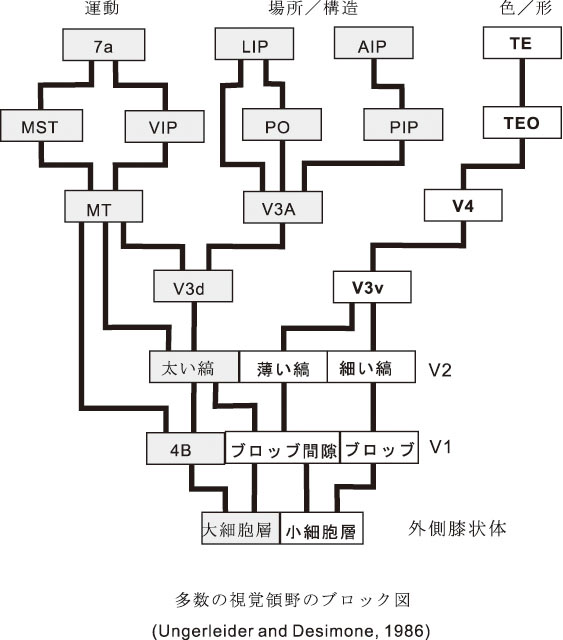
視覚系の研究で知られている背側および腹側神経路 (Goodale と Milner, 1992) になぞらえて、最近の興味深い仮説として聴覚系でも後連合野から前頭前野へ興奮の流れに背側および腹側神経路を対応させて、各々、「where」と「what」の機能を推定している(Romanski ら, 1999)。視覚および聴覚の領野のブロック図と大脳皮質内の興奮の流れについては、それぞれ、図2および図3を参照されたい。
このように、①頭頂連合野を経由して前頭葉の背側運動前野に向かう興奮の流れ(背側経路)と、②側頭葉前部を経由して前頭前野の主溝に接する腹側域(10野)に向かう興奮の流れ( 腹側経路)が霊長類(サル)の脳で存在し、視覚情報はこれらの二つの経路にわかれて処理される(Ungerleider と Mushkin, 1982)。背側経路は自己を中心として (egocentric) 物体がどこに (where)、どのように (how) 配置されているかという、空間位置の情報を処理することに関与している。他方、腹側経路は、視覚的または聴覚的事象が Gestalt (形や色;旋律や音色)としてどのようなもので (what)、いかなる意味ないし評価がなされるかということに関与している。このプロセスを通じて視覚系では、単純な形(点とか線とか)から複雑なものに至る種々の形象を認知し、色彩や動きなども加味されて情動的な視覚反応も示すようになる。同様に、聴覚系にも振動数に対応した純音、基音から、複合音、(協)和音、不(協)和音、メロディー、ハーモニー、さらに、リズム、音高ないしピッチ(これは、脳幹、小脳レベルの事象が大きく関わる)、さらに音の長さ、音と音との間の間歇(休符)、音階を伴ったフレーズ等々に関連する領域や回路についても今後の研究が俟たれるところである。この際、間脳に到達までの構成にみられる視覚系と聴覚系の間の相違(前述)を反映した情報処理過程の内容が、皮質レベルでの情報処理の同異として検討されるときには、そこに若干の問題が生じてくるように思われる。
論を先に進めることにする。視覚系についても、聴覚系についても、後連合野で受容され、処理された情報が、目的行動(演奏や彫刻作製)に進行されるためには、適応的な行動(運動、動作)に変換される必要がある。前頭前野を介する神経回路を経て運動系に高次神経活動が伝達されると言うことは、能動的発現に際して、前頭前野内で内容の組み替えがなされ、行動への意味づけをする上で必要な converter の役割が演じられているものと思われる。その意味で視覚および聴覚の後連合野から前頭葉への二つの投射経路を考察することは重要である。
「背側経路」について頭頂葉と運動前野のニューロンを調べてみると、類似した応答特性をもったニューロンが両域で存在していることが明らかにされている。従って、背側経路の空間情報は比較的直接的に運動の企画(プログラミング)に利用されている。また、ワーキングメモリー(作業記憶)の空間的タスクの遂行にも関与している。視空間、音空間から運動空間への写像変換(conversion)がこの経路を用いて比較的容易に施行される。
「腹側経路」については、視覚および聴覚の情報が側頭葉前方域に伝わったのち、鈎状束を経由して前頭葉の主に前頭前野の腹側部あるいは腹外側部に入力される(サル)。この経路で運ばれる物の同定や情動に関わる"ゲシュタルト"(形象、全体的ハーモニー、色彩、音色も含めてこの用語を使用することにする)の情報は、背側経路で運ばれる空間情報と比べると運動情報との関係は直接的ではなく、比較的独立しているようにみえる。そして、複雑な情報を伝達していると考えられる。視聴覚刺激が運動情報に変換される前に、前頭前野の前頭葉腹(外)側部(10野)のニューロンを発火させて、情報を処理し、それを運動系に伝達させるという方式をヒトの脳は採っているのである。
視覚系でこの問題を研究している坂上ら(1999a, 1999b)によれば、この前頭前野のニューロンの応答は、色や形といった視覚刺激の物理的な特徴ではなく、視覚刺激がどのような行動を指示しているかを反映するものである。このような情報を「behavioral significance (connotation)(行動的意味)」と呼ぶ。感覚情報でも運動命令でもない、その間を媒介する「意味の表象」である。特定の情報を行動に結びつけるためには、学習が必要で、前頭前野の「行動的意味」をコードするニューロンは、学習によってその活動を、より適切な刺激―反応関係を反映する方向に変化させている(Assad ら, 1998)。よって、前頭前野腹外側部からの出力は、もはや感覚情報ではなく、運動プログラミングに利用可能な翻訳された行動情報として、運動前野や補足運動野に送られる。
このようにみたとき、狭義の運動は随意的なものであるが、広義の運動は情動や意欲や選択/判断を含めた能動性機能とみなすことができる。この視点に立って演奏における運動と表現の問題を考察することは重要である。
能動的 logos の座である運動性言語野を含む前頭前野と、受動的 logos の座である感覚性言語野は連合線維によって結びつけられており、各logos野の近傍の前頭葉下部と側頭葉前方部はそれぞれ投射線維系により辺縁系(扁桃体、海馬傍回)と結びついている。このように、皮質連合野とpathosの座である大脳辺縁系が相互に密な関連性を形態的に有していることは注目される。皮質皮質間結合の研究結果によれば、こうした領野間のニューロンの働きは一旦、運動性言語中枢近傍の前頭葉皮質でシナプスを形成し、この“仲介域”で「組み替え」(pathosとlogosとの質的向上を含んだ相互浸透)を起こしたのちに、運動系皮質域に伝達されることになる。感覚性の認識が情動と結びつき、それが直接的に運動系を働かせるという方式を採らずに、生物体にとって外界の情報が集約されている前頭前野における処理を介して運動系を活性化させている。情動機能の発現は扁桃体や側坐核からの入力により、また意欲の衝動は帯状回(とくに前方域)からの入力により、維持されている。この際、視床下部や中脳からのぺプチドやアミン系からの入力が加味される。この経路を通った後に広義の運動野が活動してはじめて演奏における表現の関係が了解される。この際、重要な役割を演じているのは、小脳の学習機構と共に、線条体の運動調節統御、それに脳全体としてのバランス的・ゲシュタルト的な機能であろう。
興奮が運動野を含む前頭葉から後連合野にフィードバックされる。要約的に言うと、この投射にも背側と腹側の経路(両方向性成分を含む経路)があるが、すくなくともサルの段階では、ほとんどの線維が腹側経路を通り背側経路の成分は量的に少ない(川村、未発表)。宮下グループ(Hasegawaら, 1999; Tomita ら, 1999)はサルを用いて、このフィードバック・ルートの機能は側頭葉に保存された記憶を想起する際に、それを検索するインプットの指示を前頭葉から側頭葉に与えて、記憶を取り出していることを明かにした。すなわち、宮下らは図形の連想学習をさせた後に脳梁の後半部を切断したサルを用いて、図形連想時における側頭葉のニューロンの電気活動を調べた結果、側頭葉内記憶を検索する信号は前頭葉が制御しているという結論に達した。すなわち、前頭葉の働きを側頭葉が受けて心象(頭の内にある表象イメージ)を起こさせうることを示唆している。視覚系と聴覚系の類似性を勘案して、これは聴覚性の音(符)の順序付けを連想作用としてイメージ的に想起する過程-連想→作曲→演奏→記憶とつながる-においても関与する可能性がある。音楽家はこの回路を働かせて側頭葉に貯蔵されている記憶を想起させて、「響き」を前頭前野の働きにより活性化してひきだし、照合するように練習を行っていると考えられる。
ここで音楽を演奏する際に脳がどのように活動しているか、また、響きを捉えてよい演奏をするには、どのように脳の活動の原理にマッチさせたらよいかという点について考えてみよう。作曲家や演奏家は作品という形で具象化しようとする内的響きを、音空間として形象化させ外化させている。この作用を通常は創造と呼んでいる。専門家の脳は音源が実際に存在しない場合、つまり一般に言う響き、ないし音楽が外的空間として鳴り響いていない場合でも、音のイメージが連合野内に連続的に想起され、何等かの意味的まとまりをもった形象=作品へとこれをまとめあげるよう訓練を積む結果、演奏において、すなわち何等かの作品を、それも楽譜の形で与えられた場合でも、その楽譜から、いまは楽譜となっている作曲家のいわば原イマージュへと響きのルーツをたどり、再びこれを外的な響きとして再構築していく。彼らはそのtechneを身につけていく。fMRI(機能的核磁気共鳴映像装置)で調べてみると、演奏家の脳では楽譜をみたときに聴覚性言語野が活動しており、音の流れを一種の言語ないし記号の流れとして捉えているらしいのである。この際、視覚性言語野は活動していない点から考えて、楽譜を単に視覚的にイメージングするのではなく、記憶に書き込まれ符号化された連続的な内音の響きを、小脳・大脳関連ループと大脳・基底核・視床関連ループのなかで学習化された意識下(随意性でないという意味)でスイッチオンして呼び覚ますことによって、この内的響きを身体運動によって外化させるために必要な運動系を発動させて演奏を行っていると考えられる。聴覚のみならず、視覚、体性(皮膚および粘膜を含む)感覚、内臓感覚などの全感覚が総動員されて、そこに感動が生まれてくる。それによって、言語野に創り出されてくるimage(イマージュ)を具象化させるという、内的イマージュの外化の過程が響きの創作であろう。それが音楽の創造、とくにこの場合は演奏による創造を成立させる。一般的に言って、創作活動とはすなわちヒトの脳の総合的働きによって形成される内的イマージュを作品として外化させる働きを指すが、その際に、言語野における概念を作品における原イマージュとして捉えることは可能であろう。なお、こうした原イマージュの形成には、とくに記憶との関わりが重要であり、既に記憶されているものの外化を一般的には創造と判断しないのであるから、各皮質における記憶と関わりながらも、その関連性の中から記憶にないものを組み立て、イマージュとして外化させることが要求される。とはいえ、音楽の場合、創造に用いられる素材としての音は原則として12種類であり、これはすべて音楽家の脳に記憶されている。したがって、音楽の創造は基本的には12の音の組み合わせ方の問題につきると言い得るが、限られた文字数にもかかわらず文学表現の可能性が無限であるのと同様に、音楽の創造の可能性も無限である。ただし、限られたものから何故このような無限性が抽き出され得るかの脳の働きについては今後の解明に俟たなければならない。
原イマージュがゲシュタルト化される時には、前頭前野→高次運動野→第一運動野(そして一部の興奮が後連合野、とくに情動および言語の関連領域にフィードバックされる)として構成される神経回路が言語野および情動の回路と結びついて形成される。歌詞の内容を識って傾聴するオペラや声楽曲において、意味内容を知らずに聞く場合に較べて情動中枢がより強く刺激される。そして、芸術作品によって伝えられるべき事柄の認知へと享受者の脳を活性化させるが故に、言語中枢の関与が重要な役割を演じていることをあらためて意識させる。
校正の段階で貴重なご教示を賜った丸山桂介先生(東京音楽大学音楽学)、丸山直滋先生(新潟大学聴覚生理学)および鬼頭順三先生(名古屋大学解剖学)に感謝いたします。
Assad, W.F., Rainer G., and Miller, E.K. Neural activity in the primate prefrontal cortex during associative learning, Neuron 21(1998)1399-1407.
Baizer, J.S., Ungerleider, L.G., and Desimone, R. Organization of visual inputs to the inferior temporal and posterior parietal cortex in macaques, J. Neurosci. 11 (1991) 168-190.
Desimone. R., Schein. S.J., Moran. J., and Ungerleider. L.G. Contour, color and shape analysis beyond the striate cortex, Vision Res. 25 (1985) 441-452.
Goodale, M.A., and Milner, A.D. Separate visual pathways for perception and action, TINS 15 (1992) 20-25.
Hasegawa, I., Hayashi, T., and Miyashita.,Y. Memory retrieval under the control of the prefrontal cortex, Ann. Med. 31 (1999) 380-387.
He, J., Hashikawa T., Ojima, H., and Kinouchi, Y. Temporal integration and duration tuning in the dorsal zone of cat auditory cortex, J. Neurosci. 17 (1997) 2615-2625.
Kaas J.H., and Hackett, T.A. ‘What’ and ‘where’ processing in auditory cortex, Nature neurosci. 2 (1999) 1045-1047.
Kawamura, K. Corticocortical fiber connections of the cat cerebrum. I, II,III, Brain Res. 51 (1973) 1-22,23-40,41-60.
川村光毅
連合野"の線維結合(I)皮質間結合
−−サルとネコの皮質間結合の比較と"連合野"の発達についての試論−−
神経研究の進歩 21(1977)1085-1101.
Kawamura, K., and Norita, M. Corticoamygdaloid projections in the rhesus monkey. An HRP study, Iwate med. Ass. 32 (1980) 461-465.
丸山直滋 聴覚情報の受容と認識 新医学大系 7巻「刺激の受容と生体運動」pp.91-113, 1995, 中山書店
Romanski, L.M., Tian B., Fritz J., Mishkin M., Goldman-Rakic P.S., and Rauschecker, J.P. Dual streams of auditory afferents target multiple domains in the primate prefrontal cortex, Nature Neurosci. 2 (1999) 1131-1136.
Sakagami, M., Tsutsui, K. The hierarchical organization of decision making in the primate prefrontal cortex, Neurosci. Res. 34 (1999a) 79-89.
坂上雅道、筒井健一郎 神経活動から見た前頭前野の2つの柔軟性 -認知機能の背景として-、 脳の科学 21 (1999b) 617-625.
Shima., K., and Tanji., J. Role of cingulate moter area cells in voluntary movement selection based on reward, Science 282 (1998) 1335-1338.
丹治 順 脳と運動-アクションを実行させる脳 1999 共立出版。.
Tomita, H., Ohbayashi, M., Nakahara, K., Hasegawa, I., and Miyashita, Y.
Top-down signal from prefrontal cortex in executive control of memory retrieval, Nature 401 (1999) 699-703.
Ungerleider, L. G., and Mishkin, M.
In : Analysis of visual behavior, eds by Ingle, D. I., Goodale, M. A., and Mansfield, R. J. W. , 1982, pp.549-586, MIT Press, Cambridge, Massachusetts.
臨床精神医学 2001年 7-16頁 増刊号 (アークメディア)より許可を得て転載。