
視蓋・小脳投射系を中心とする視覚運動および聴覚運動に関する考察
川村光毅
| はじめに | |
| 1.上丘の構成と機能 | |
| 2.下丘の構造と機能 | |
| 3.視蓋(上丘と下丘)から橋核への投射 | |
| 4.橋核小脳皮質投射(視覚と聴覚) | |
| 5.視蓋および視蓋前域から下オリーブ核への投射 | |
| 6.小脳の"視覚"野 | |
| 7.小脳の"聴覚"野 | |
| 8. ファウスト(F)とメフィスト(M)の対話 | |
| 文献 |
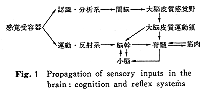
視覚系にしろ、聴覚系にしろ、体性感覚系にしろ、感覚入力が中枢神経系内で伝達される仕方に大別して二種類ある。一つは、認識分析系でいくつかの中枢核を介したのち大脳皮質に伝達される系で、他は、脳幹内で神経回路が形成される運動反射系でこれには小脳が関連する。第一の認識系はどちらかというと新しい系で、基本的に独立した系となっており、大脳皮質でも皮質下の中継核でも異なる感覚入力に反応する細胞は分離して存在している(たとえば視覚伝達の中継核である外側膝状体や聴覚のそれである内側膝状体など)。これに対して、運動反射機能に関連するシステムの方は古い系で、網膜から上丘に達する視覚性入力と、コルチ器官内の有毛細胞から蝸牛核、上オリーブ核などを経て下丘に達する聴覚性入力とが、さらに小脳にまで伝達される神経路を考えてみると、視覚または聴覚に反応する神経細胞群は領域的にみる限り必ずしも分離されてないことに気付く。たとえば、橋核内の背外側部、小脳虫部の中央部では両者ともほとんど同一領域内に存在する。小脳には体性感覚系や前庭系の情報が特定の部位に入ってくるが、これらの入力も運動系と密接な関係をもっている。このほかに、嗅覚性、味覚性などの入力も存在すると思われるが、嗅覚、味覚、情動などに関連する小脳の研究はこれからの分野であろう(Fig. 1)
。
以下に、視覚および聴覚刺激(しばしば遠隔受容刺激teleceptive impulsesというように古風な名称でまとめられる)の小脳への伝達について、この約10年間に得られたわれわれの研究成果(実験材料は主としてネコ)を中心に若干の考察を加えて総括してみたい。
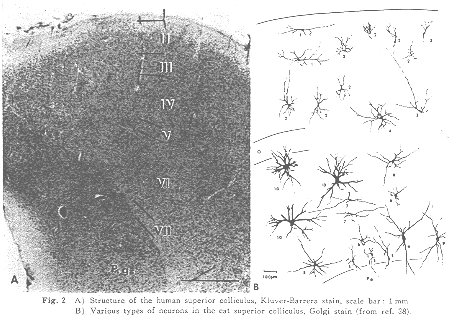
哺乳類の上丘の顕著な特徴は、線維層(白質層)と細胞層(灰白層)が交互に、ほぼ表面に平行して並ぶ7層(帯層、浅灰白層、視神経層、中間灰白層、中間白質層、深灰白層、深白質層)から成る構造をしていることである(Fig.2)。その構造の配列は、ほとんどすべての哺乳動物で驚く程よく似ている。詳細は他書20,38)にゆずるが、大部分が小型(8-15μm)の神経細胞から成り浅層(主として浅灰白層)は“視覚性”の領域で網膜および視覚領皮質からの線維が終わる。それより深部の中間層・深層には中型の細胞が多くみられ、とくに深層の外側2/3には大型(35-60μm)の細胞がみられ、一見して浅層とは構造を異にする。ここには聴覚領、体性感覚運動領、連合領など視覚領以外の大脳皮質全域31,34)からの線維が終わる。そのほか、視蓋前域、二丘旁体、下丘の外
前述したように、上丘深層は、聴覚、体性感覚などの情報を受けとっている。聴覚や体性感覚の上丘への入力も視覚入力に関する程ではないにしても、かなりの局在性(tonotopic, somatotopic)をもっている49-51)。視覚にしろ聴覚にしろ、動く光源または音源の目標に対して、上丘は眼や頭を動かして、その位置を知り、眼球(や頭)の運動方向を規定しているといえよう。Robinson41,42)によれば、頭部座標系における目標位置(頭の中での眼の位置)から眼球位置を差し引いて網膜誤差を得、それに基づき誤差修正の過程で、目標位置を頭部を基準とする座標(somatotopic)系内の位置に変換する仕組みがある筈である。(閉眼時において音源を定位する場合も同様に考えられよう。なお、眼球運動の発生機構については加瀬の総説23)を読まれたい。)
下丘は視蓋の後方域が隆起した構造物で、哺乳動物では前方域が隆起した上丘とは明瞭に区別される。上丘とは異なり線維と細胞の密な層が交互に配列するいわゆる層構造は認められない。ところで、retinotopicの局在配列がみられる上丘の浅層に相当する部分は下丘では中央部を占める中心核(central nucleus)で、ここにはほぼ同心円状に配列するtonotopicの局在が存在する。この部位は聴覚性認識・分析の伝達系の中継核で(大脳皮質聴覚領からの線維もこの中心核に終止する)、主要な上行性の投射域は内側膝状体である。これに対して、中心核の外側と背側を板状にとりかこむように存在する外核と中心周囲核は、とくに前者は、皮質聴覚領7,44,53)や外側毛帯核の背側核および腹側核35)から弱い投射を受けるが、主たる入力は同側の中心核の腹外側部、対側の蝸牛核と同側の内側上オリーブ核と外側毛帯の腹側核からおよび下丘交連線維である3,36)。小脳核、知覚運動領皮質、脊髄、三叉神経核などからの入力は証明されておらず、この点上丘の深層へ投射する体性感覚運動に関連した入力を含む種々の求心線維の存在とは趣を異している。黒質・下丘投射の存在は証明されているが、下丘内終止線維の分布域やその粗密性についてのデータに欠ける(恐らく外核への投射が多いもではないか?)。
なお、下行性の遠心路に関しては、次項に述べる橋核への投射(ほとんどが外核と中心周囲核が起始領域である)のほかには、強い投射が中心核から、弱い投射が外核や中心周囲核から両側性に上オリーブ核や蝸牛核に対して存在する13)。驚くべきことに、橋下部や延髄の網様体には下丘から直接は線維を送っておらず、下丘・下オリーブ路も存在しない。
最後に、上丘と下丘間の連絡であるが、下丘の外核と中心周囲核から上丘の中間層と深層に対してはかなりの強い投射がみられるが15)、上丘から下丘への投射の存在は否定的である28)。なお、ネコおよびサルで網膜神経節細胞の軸索が上丘を経て一部下丘の中心周囲核に達するものがあるという21)。
橋核から起こる苔状線維はすべて小脳皮質に終わる。途中、小脳核へ終止するものがあるかどうか疑問視されており、存在したとしても僅かである8)。また、橋核ニューロンは小型で、介在ニューロンはほとんど無いといわれている。
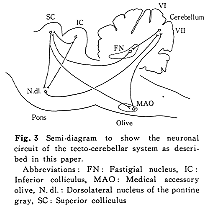
本項および以下の項で視蓋・小脳投射系について述べるが、その主な経路の全体像を模式的にFig.3に示してある。まず、上丘・橋核投射ニューロンについてみると、起始細胞は上丘の視神経層より深部に存在する中型(径, 25-40μm)および小型(径, 10-25μm)の円形、楕円形または紡錘形の細胞で、そのうち小型細胞が80%以上占める14)。そして、同側性に背外側亜核に投射する。厳格ではないが、上丘前方部(中心視野再現域)からのものは橋核の前方域に上丘後方部(周辺視野再現域)からのものは橋核の後方域にというように、上丘・橋核路には前後方向に局在配列がみられる28)。視覚認識系の神経路(たとえば網膜外側膝状体路)において、起始域と終止域が点対点(point to point)の対応で連絡している事実と比較して興味深い。
他方、下丘・橋核路の起始細胞は、大部分が外核と中心周囲核内に存在する小型(径、10-20μm)のニューロンで中心核からの投射は非常に弱い13)。注目すべき点としては、同じく上丘へ線維を送る外核や中心周囲核内の細胞と軸索を共有するものは稀で、ニューロン自体は混っているが独立して存在している15)。橋核内の終止域をみると、上丘からの終止域とかなり重複している。しかし、よく比較検討してみると、上丘・橋核線維の終止域はより尾側にまでひろがり、また、下丘・橋核線維の終止域はより腹側までひろがっており、僅かながら両者間に相異が認められる26)。橋核を経由する視覚入力と聴覚入力の小脳皮質終止域にも僅かながら差異が生じている47,48)。
大脳皮質から橋核への投射を視蓋からの投射と比較してみると興味ある事実に遭遇する。すなわち、聴覚皮質からの橋核内終止域は背外側部に限局しており、上丘や下丘からの線維の終止域とほぼ一致している6)。すなわち、視覚および聴覚性(teleceptive)の運動・反射に関連した情報を伝達する視蓋橋核路は、少なくともネコにおいては、橋核内の非常に限局した部位(背外側亜核)で大脳皮質から下行する聴覚性の“フィードバック”情報と統合される可能性がある。これに反して、視覚皮質からの軸索終末は、橋核吻側の腹側核に特徴ある柱状配列を示して終っている。視蓋・橋核路の終止部位との間に全く重複は認められない。さらにいえば、この視蓋からの投射線維の終止域は、一部、その中央部および外側において感覚運動皮質からの投射域と重なり、視覚関連領域であるClare-Bishop域(中シルビウス溝外側壁皮質部位)や前シルビウス溝周囲皮質の前方域(Muckeら37)により視覚性誘発反応が得られた部位)からの線維の終止域とも重複する部分があるように思われる。
詳しい知見に乏しいが、このほか、視覚性の情報として橋核に入ってくるものには、外側膝状体腹側核が橋核の吻側内側部に9,12)、視蓋前域からの入力も橋核内側部や一部外側部にも終止する11,22)。さらに、小脳核や脊髄からも橋核への投射が存在する46,54)。
橋核(主として前方部)で調べた電気生理学的仕事4,10)によれば、ほとんどすべての橋核ニューロンは一つの種類の感覚(視覚性、聴覚性または体性感覚性)の刺激にのみ反応する。一方、無麻酔ネコを用いて調べたBoydとAitkin5)の報告によれば、橋核背側および外側のニューロンは単一ユニットの反応で、視覚と聴覚の両方の刺激に反応する(bimoda)ものが40%存在するという。
視蓋橋核線維の終止域である背外側亜核からは小脳虫部のVIおよびVII小葉(とくに後者)に非常に強い投射があり、このほかに、VIII、IX小葉や、第I脚、第II脚、旁片葉および正中旁小葉にも弱く投射が存在する19,30)。この虫部皮質中央部は、古くから生理学的研究47,48)により視覚および聴覚性の入力を受けることが知られており(“classical teleceptive area”)、微小電気刺激を与えるとサッケードをひき起こすことも明らかにされている45)。HRPをこの小脳領域に注入して橋核内の起始ニューロンをネコで調べてみると、橋核内に前後の方向に柱状に配列する起始細胞集団がA,B,C,Dと4群認められる19)。A群は背外側亜核にあり、前述のごとく、上丘14,28)、下丘13,26)、大脳皮質聴覚領6,28)からの投射を受ける。B群は橋縦束に接してその腹側および内側部にあり、一部が大脳の視覚領からの投射域と重なる。また、C群は外側亜核にあり、視覚前域からの投射を僅かながら受け、D群は内側亜核にあり、外側膝状体と視覚前域からの投射を受けるものと思われる(前をみよ)。このようにみてくると、小脳皮質虫部中央部には、視覚および聴覚に関連する種々の領域からの入力がかなりの程度集中していることがわかる。また、最近、旁片葉および虫部垂最前部は、橋核前方部の腹側および内側域を介して大脳視覚領皮質からの強力な投射を受けていることが明らかにされた43)。
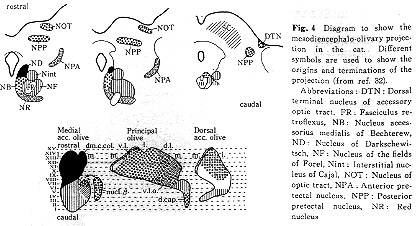
上丘からは、その中間層および深層にあるニューロンの一部が対側性に内側副オリーブ核の尾半内側部に線維を送っているが、下丘から下オリーブ核への直接投射は存在しない。視蓋前域(前および後視蓋前核)および副視索背側終止域(通常視蓋前核には含めない)からの投射はすべて同側性で一定の局在性をもって背側副オリーブ核の前半域外側部やβ核、さらに主オリーブ核の一部(背帽域を含む)に終わる32,40)。しかし、上丘からの線維の終止域とは重複しない(Fig.4)。また、大脳皮質視覚領と聴覚領はいずれも下オリーブ核には線維を送っていない。以上のようにみてくると、視覚および聴覚性の刺激は上丘を介して内側副オリーブ核尾半内側部に、また、視蓋前域を介して、背側副オリーブ核、β核、背帽核、主オリーブ核の一部(腹側板の前方内側部と腹外側突出部ventrolateral outgrowth)に入ってくる。これらオリーブ核領域におけるニューロンの反応特性を調べた生理学的研究は背帽域(dorsal cap)を除いては発表されていない。視覚性刺激と聴覚性刺激のinteractionに関しては、上丘や橋核(前述)では調べられているが、下オリーブ核においては全くみられてないのが現状である。
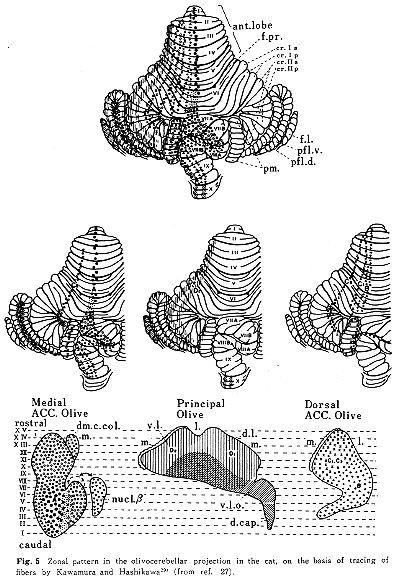
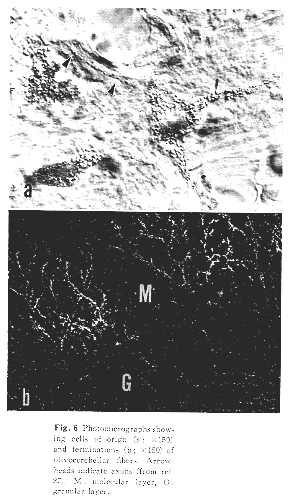
小脳皮質虫部第VI~VII小葉には上丘からの入力が橋核背外側亜核を介して苔状線維として入ってくるが、そのほか、内側副オリーブ核を介して登上線維が同一小脳皮質域に集中して入ってくる27,29)(Figs.5&6)。この領域における仕事としては、視覚野と聴覚野とが領域内に重複しているという知見にとどまっており、神経細胞レベルの相互作用に関する生理学的研究は未だみられない。この小脳虫部領域での視覚性機能については、近年、Kaseら24,25)の電気生理学的研究により、衝動性(saccade)眼球運動のみならず滑動性追従(smooth persuit)眼球運動にも関連した領域であることが明らかにされた。これに比肩し得るネコやサルを用いた聴覚(運動)性機構に関する研究が望まれる所であるが、音源の定位(orientation)に関与するものと思われる。
次に、小脳皮質虫部の第VI~VII小葉にあるプルキンエ細胞の軸索終止部位である室頂核尾半部について述べる。現在、われわれは、電気刺激を視神経交叉部に与えて誘発電位を小脳で記録しその性質を調べている。室頂核で得られた応答は、第VII小葉のそれに比してspike発射頻度が少なく、また、spike発射後の抑制は約365msec続きプルキンエ細胞のものよりも約2倍も長い。抑制後のrebound現象も著明である(発表準備中)。
室頂核尾半部のニューロンが軸索を上丘および視蓋前域に送ることも明らかにされた18,33,52)。以上の所見結果から、上丘・橋核および下オリーブ核・小脳・上丘という神経回路のループの存在が考えられるが、これらの視覚および聴覚に関連する運動反射ないし光源や音源の定位に密接に関与しているものと思われる。この際、光や音に反応する運動・反射性の機構には以上みてきた神経経路の構成から考えて相補的なものがあるのであろうか。
形態学的研究の結果から、小脳皮質へ伝達される聴覚性刺激の主たる経路は、内耳の有毛細胞からいくつかのニューロンをかえて下丘の中心核に達した上行性の聴覚認識系の成分のうち、その一部が下丘内で外核および中心周囲核(とくに前者)の亜核に伝えられ、次いで背外側橋核を経由して小脳のVIおよびVII小葉に至るものと考えられる。この経路は脳幹レベル内で形成されるいわば反射性あるいは運動性の経路ともいうべきものである。このほかに、この系を制御すると考えられる聴覚領皮質からの入力があり、この下行性の投射系もやはり同側性に背外側橋核に終止している。この皮質橋核路には、直接、橋核(恐らく橋核細胞の樹状突起遠位部に終止する)に至るものと、上丘および下丘内で一旦ニューロンをかえたのち橋核の同一亜核内に到達するものと、少なくとも二種類が存在する。しかしながら、これら“反射・運動系”と“認識・制御系”との間に起こる橋核内における統制の機構についてはいまだ明らかにされてない。また、これらの視蓋域や橋核域内における聴覚情報と他の感覚情報との間の相互作用などについても多くの未解決の問題が残されている。
最近の解剖・生理学的研究によれば、視蓋部(上丘や下丘)や背外側橋核には、聴覚性入力のほかに大脳皮質、脳幹、さらに脊髄レベルからの非聴覚性の入力も明らかに存在する。こうしてみると下丘・橋核・小脳投射系は聴覚刺激の伝達にかかわる役割を第一義的とするとはいうものの、決して単純なものではない。
われわれは、電気生理学的手法を用いてこれらの点について研究を進めるにはいまだ至っていないが、文献的に調べてみてもその数は少ない。たとえば、AitkinとBoyd1,2)のネコを用いた研究によれば、聴覚性反応に関する限り、小脳虫部皮質および背外側橋核における神経細胞の性質は、下丘核の中心核内にみられる細胞のように応答周波数範囲の狭い且つ鋭い反応野が得られるというタイプのものよりもむしろ広い周波数範囲の音に反応する(broadly tuned)という外核や中心周囲核に存在する細胞のもつ反応特性に類似しているという。これらの諸領域が聴覚運動性の小脳皮質求心路の中継核に相当するというわれわれの知見は彼らの所見結果を支持するにふさわしい形態的基盤を提供するものと考えられる。
M:青息吐息の御様子ですね。御邪魔でしょうか。
F:胡散臭い奴だ。勝手にしろ。
M:私にとっては、聴覚系の方が面白そうですね。人間は視覚で物事を判断しすぎる。
F:認識・判断系のことを問題にしているのではない。そちらは大脳の出番で、今の勤めは小脳だ。
M:小脳の機能として、視覚運動反射とか聴覚運動反射とか、また、光源や音源の定位に関連するなど御託宣のようですが、大脳や脳幹網様体との関連も禄に顧みないでよくも言えたもんだ。さっぱり実体的イメージが湧きませんね。
F:小癪な。暇とアイデアを呉れ。小天地にそれがあるのなら連れて行け。
M:いつもの御性急がまたぞろ出てきた。あちらでは、前庭機能の考察も不可欠ですよ。これは聴覚機能と関係がある。どちらも第八脳神経でしょう。
F:昔の生成論は知らないが、前庭系と聴覚系との間には血筋の違いが明瞭なのだ。一方は古く羊水中で釣合いをとる時分から発達し、小脳への最初の闖入者だ。他方は、新参者だ。小脳界隈にはすでに強固な縄張がつくられてしまっていたので前庭組のように大挙して直接入りこむことがむずかしかったのだ。代りに延髄の蝸牛核あたりで擬似小脳組織を作っているんだろう。和衷協同という訳にはいかぬ。
M:独善がりもいいとこだ。ともかく、視覚系も新しい。視覚系人間などという言葉も耳にする。
F:その通りだ。しかし、聴覚系も同様に、いや、それ以上に新しい。
M:オランダ人は視覚的でドイツ人は聴覚的だ。レンブラントとベートーヴェンが象徴的だ。同じくゲルマン系だが。
F:話を逸らしては困る。皮相な現象論をふリまわす輩なんぞに用はない。
M:大変なお冠だ。とにかく、視覚と聴覚とは別物。視覚は間脳に入り、聴覚は延髄に入る。それに一方は粒子と波の折衷混体で、他方は単に波として源(みなもと)が説明される。
F:似た処もあるではないか。小脳でも橋核でも、縄張りがほとんど完全に重なっている。視蓋・小脳投射系に於いては、上丘も下丘も同声異俗。物質の最高段階にまで発展した形態などといばり腐っている大脳皮質にだって完全独立王国といえる場所は純粋に認識・分析系に属する所だけのようで、後連合野にも前頭前野にも重複したり隣接している部位がいくつかある。
M:視聴覚合体論か。我田引水とはこのことか。ところで、相補性とか多重感覚性とかおっしゃっているようですが矛盾を感じませんか。
F:唯我独尊的中枢は脳の中には一つもない。対立物は相互に浸透し、それらは高い段階で統一される。
M:物の臭みが強すぎて流行(はやり)ませんな。
F:意識が物質より根元的だと考えるような干からびた尨犬の脳髄からは、実りある発展はあり得ない。
M:羊皮紙綴りの知識は灰色だ。実際に験することだ。豊かな人生はみどり色だ。言い残してはございませんか。
F:耳目を属(しょく)すという言葉がある。眼を開いて物事に刮目するなら、萬物流転の位置を確と定めることも左程苦にもなるまい。眼を閉じてもそれができるだろうか。こう考えてみると、視覚と聴覚の連続性という意味が少しははっきりする。前方は網膜にまかせられるとしても、側方・後方はコルチ器官の助けが要る。これが相補性だ。脳研究者は、生理学的・生化学的言葉で説明ができない限り迷うものだ。
M:Cogito, ergo sum. Nihil sub sole novum.
1) Aitkin LM, Boyd J:Responses of single units in cerebellar
vermis of the cat to monaural and binaural stimuli. J Neurophysiol 38:418-429, 1975
2) Aitkin LM, Boyd J:Acoustic input to the lateral pontine
nuclei. Hearing Res 1:67-77, 1978
3) Anderson RA, Roth GL,
Aitkin LM, Merzenich MM:The efferent projections of the central
nucleus and the pericentral nucleus of the inferior colliculus in the cat. J Comp Neurol
194:649-662, 1980
4) Baker J, Gibson A,
Glickstein M, Stein J:Visual cells in the pontine nuclei on
the cat. J Physiol 255:415-433, 1976
5) Boyd J, Aitkin L:Responses of single units in the
pontine nuclei of the cat to acoustic stimulation Neurosci Lett 3:259-263, 1976
6) Brodal P:The corticopontine projection in the
cat. The projection from the auditory cortex Arch ital Biol 110:119-144, 1972
7) Diamond IT, Jones EG,
Powell TPS:The projection of the auditory cortex
upon the diencephalon and brain stem in the cat. Brain Res 15:305-340, 1969
8) Dietrichs E, Bjaalie JG,
Brodal P:Do pontocerebellar fibers send
collaterals to the cerebellar nuclei? Brain Res 259:127-131, 1983
9) Edwards SB, Rosenquist
AC, Palmer LA:An autoradiographic study of ventral lateral geniculate
projections in the cat. Brain Res 72:282-287, 1974
10) Glickstein M, Stein J,
King RA:Visual input to the pontine nuclei.
Science 178:1110-1111, 1972
11) Graybiel AM:Some efferents of the pretectal
region in the cat. Anat Rec 178:365, 1974a
12) Graybiel AM:Visuo-cerebellar and cerebello-visual
connections involving the vental lateral geniculate nucleus. Exp Brain Res 20:303-306, 1974b
13) Hashikawa T:The inferior colliculopontine neurons
of the cat in relation to other collicular descending neurons. J Comp Neurol 219:241-249, 1983
14) Hashikawa T, Kawamura K:Identification of cells of origin of
tectopontine fibers in the cat superopr colliculus:an experimental study with the
horseradish peroxidase method. Brain Res 130:65-79, 1977
15) Hashikawa T, Kawamura K:Retrograde labeling of ascending and
descending neurons in the inferior colliculus. A fluorescent double labeling study in the
cat. Exp Brain Res 49:457-461, 1983
16) Hikosaka O, Wurtz RH:Visual and oculomotor functions of
monkey substantia nigra pars reticulata. IV. Relation of substantia nigra to superior
colliculus. J Neurophysiol 49:1285-1301, 1983a
17) Hikosaka O, Wurtz RH:Effects on eye movemonts of a GABA
agonist and antagonist injected into monkey superior colliculus. Brain Res 272:368-372, 1983b
18) Hirai T, Onodera S,
Kawamura K:Cerebellotectal projections studied
in cats with horseradish peroxidase or tritiated amino acids axonal transport. Exp Brain
Res 48:1-12, 1982
19) Hoddevik GH, Brodal A,
Kawamura K, Hashikawa T:The pontine projection to the
cerebellar vermal area studied by means of the retrograde axonal transport of horseradish
peroxidase.Brain Res 123:209-227, 1977
20) Huerta MF, Harting JK:The mammalian superior colliculus:studies of its morphology and
connections. in Comparative Neurology of the Optic Tectum, ed. by Vanegas H, Plenum
Publishing Corporation 1984, p 687-777
21) Itaya SK, Van Hoesen GW:Retinal innervation of the inferior
colliculus in rat and monkey. Brain Res 233:45-52, 1982
22) Itoh K:Efferent projections of the pretectum
in the cat. Exp Brain Res30:89-105, 1977
23) 加瀬 学:眼球運動-その発生機構について-.眼科 MOOK, No. 22, 41-53, 1984
24) Kase M, Miller DC, Noda
H:Discharges of Purkinje cells and
mossy fibres in the cerebellar vermis of the monkey during saccadic eye movements and
fixation. J Physiol(London)300:539-555, 1980
25) Kase M, Noda H, Suzuki
DA, Miller DC:Target velocity signals of visual
tracking in vermal Purkinje cells of the monkey. Science 205:717-720, 1979
26) Kawamura K:The pontine projection from the
inferior colliculus in the cat. An experimental anatomical study. Brain Res 95:309-322, 1975
27) 川村光毅:小脳の帯状構造とオリーブ小脳投射.生体の科学 31:139-148, 1980
28) Kawamura K,:Brodal A:The tectopontine projection in the cat:an experimental anatomical study with
comments on pathway for teleceptive impulses to the cerebellum. J Comp Neurol 149:371-390, 1973
29) Kawamura K, Hashikawa T:Olivocerebellar projections in the
cat studied by means of anterograde axonal transport of labeled amino acids as tracers.
Neuroscience 4:1615-1633, 1979
30) Kawamura K:Hashikawa T:Projections from the pontine nuclei
proper and reticular tegmental nucleus onto the cerebellar cortex in the cat. An
autoradiographic study. J Comp Neurol 201:395-413, 1981
31) Kawamura K, Konno T:Various types of corticotectal
neurons of cats as demonstrated by means retrograde axonal transport of horseradish
peroxidase. Exp Brain Res 35:161-175, 1979
32) Kawamura K, Onodera S:Olivary projections from the
pretectal region in the cat studied with horseradish peroxidase and tritiated amino acids
axonal transport. Arch ital Biol 122:155-168, 1984
33) Kawamura S, Hattori S,
Higo S, Matsuyama T:The cerebellar projections to the
superior colliculus and pretectum in the cat:an antoradiographic and horseradish
peroxidase study. Neuroscience 7:1673-1689, 1982
34) Konno T:Patterns of organization of the
corticotetal projection of cats studied by means of the anterograde degeneration method. J
Hirnforsch 20:433-444, 1979
35) Kudo M:Projections of the nuclei of the
lateral lemniscus in the cat:an autoradiographic study. Brain Res
221:57-69, 1981
36) Kudo M, Niimi K:Ascending projections of the inferior
colliculus in the cat:an autoradiographic study. J Comp
Neurol 191:545-556, 1980
37) Mucke L, Norita M,
Benedek G, Creutzfeldt O:Physiologic and anatomic investigation
of a visual cortical area situated in the ventral bank of the anterior ectosylvian sulcus
of the cat. Exp Brain Res 46:1-11, 1982
38) 車田正男、川村光毅:中脳蓋-上丘を主として-.P286-295(橋本、山元編:人体組織学。第8巻;神経、朝倉書店、1984)
39) Ogasawara K, Kawamura K:Cells of origin and terminations of
the trigeminotectal projection in the cat as demonstrated with the horseradish peroxidase
and autoradiographic methods. Okajimas Folia Anat Jpn 58:247-264, 1982
40) Onodera S:Olivary projections from the
mesodiencephalic structures in the cat studied by means of axonal transport of horseradish
peroxidase and tritiated amino acids. J Comp Neurol 227:37-49, 1984
41) Robinson DA:Models of the saccadic eye movement
control system. Kybernetik 14:71-83, 1973
42) Robinson DA:Oculomotor control signals(眼の働きを操る神経信号)。生体の科学26:165-177, 1975
43) Robinson FR, Cohen JL,
Mey J, Sestokas AK, Glickstein M:Cerebellar targets of visual pontine
cells in the cat. J Comp Neurol 223:471-482, 1984
44) Rockel AJ, Jones EG:The neuronal organization of the
inferior colliculus of the adult cat. I. The central nucleus. J Comp Neurol 147:11-60, 1973
45) Ron S, Robinson DA:Eye movements evoked by cerebellar
stimulation in the alert monkey. J Neurophysiol 36:1004-1022, 1973
46) Ruegg DG, Eldred E,
Wiesendager M:Spinal projections to the
dorsolateral nucleus of the caudal basilar pons in cat. J Comp Neurol 179:383-392, 1978
47) Snider RS, Eldred E:Electro-anatomical studies on
cerebro-cerebellar connections in the cat. J Comp Neurol 95:1-16, 1951
48) Snider RS, Stowell A:Receiving areas of the tactile,
auditory, and visual systems in the cerebellim. J Neurophysiol 7:331-358, 1944
49) Stein BE:Development of the superior colliculus.
Ann Rev Neurosci 7:92-125, 1984
50) Stein BE,
Magalhaes-Castro B, Kruger L:Superior colliculus:visuotopic-somatotopic overlap. Science
189:224-226, 1975
51) Stein BE,
Magalhaes-Castro B, Kruger L:Relationship between visual and tactile
representations in cat superior colliculus. J Neurophysiol 39:401-419, 1976
52) Sugimoto T, Mizuno N,
Uchida K:Distribution of cerebellar fiber
terminals in the midbrain visuomotor areas:an autoradiographic study in the cat.
Brain Res 238:353-370, 1982
53) Van Noort J:The structre and connections of the
inferior colliculus. An investigation of the lower auditory system. Assen, Van Gorcum,
1969
54) Walberg F, Brodal A:Spinopontine fibers in the cat. An
experimental study. J Comp Neurol 99:251-287, 1953
55) Wurtz RH, Albano JE:Visual-motor function of the primote
superior colliculus. Ann Rev Neurosci 3:189-226, 1980
56) Wurtz RH, Goldberg ME:Activity of superior colliculus in
behaving monkey. III. Cells discharging before eye movements. J Neurophysiol 35:575-586, 1972
脳と神経 36(12):1149-1158,1984(医学書院)