大脳辺縁系の解剖(limbic system, le grand lobe limbique) |
川村光毅、小野勝彦

| 辺縁葉limbic lobeと辺縁系limbic systemについて | |
| 海馬の解剖 | |
| 扁桃体(扁桃核)の解剖 | |
| 中隔核とその関連領域 | |
| 辺縁系のサブシステム | |
| おわりに | |
|
辺縁葉limbic lobeと辺縁系limbic systemについて
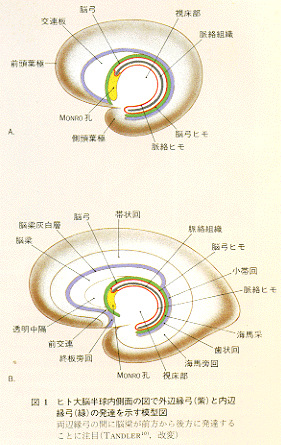
大脳辺縁系という用語の内容は、文献的に考察してみると年とともに拡げられルースになってきていることがわかる。歴史的にみると、前脳胞から発芽した突起(内に腔所を有する)から大脳半球が内側部と外側部と2つリング状に形成されるが、そのリングの中心は室間孔(Monoro孔)であるとMeynert(1872)がすでに記載している。さらにこの内側環部は帯状回、海馬形成、前梨状皮質から構成され、皮質域はこのinner ringに終焉すると述べている。その後1878年にBroca,P.が、Monro孔の周囲の脳の中心部分を縁どって(limbus)リング状にとりまいている
表1 辺縁系の分類と範囲(小池上)5) |
広義の嗅脳 1)だいたい古皮質とその皮質下部にあたるもの(狭義の嗅脳) 大脳辺縁系 後部:終板旁回(梁下回)・前有孔質・梨状葉前野・梨状葉皮質・扁桃体 島および弁蓋部・側頭葉極部・上、中、下側頭回の前部・前頭葉眼窩面後部(Area13)・視床の一部(前核を主とす)・視床髄条・手綱核・[脚間核]・[視床下部]・中脳辺縁系領野 |
表2 辺縁系の分類と範囲(小池上)5) |
| 1. 固有辺縁系 limbic
structures proper 海馬・海馬采・歯状回・海馬鈎・海馬旁回(海馬回)・帯状回(前部)・扁桃体(扁桃核)・梨状葉・梨状葉前野(前梨状葉)・中隔部・脳梁灰白層 対角帯とその床核・嗅結節・梁下野(旁嗅領)・前有孔質・終板旁回(梁下回) 2. 旁辺縁系領域paralimbic area 分野13(後眼窩回)・島・前障・側坐核・帯状回(後部) ・ 視床前核・視床髄板内核・視床枕核・手綱核・脚間核・視床下部(とくに乳頭体)・視床旁下部・中脳辺縁系野(Nauta)・背側および腹側被蓋核 ・ (Gudden)・上側頭回・側頭葉極部・楔前部 |
灰白質領域を一括して、"le grand lobe limbique"大(脳)辺縁葉と呼称した。他方、辺縁系という用語はこの辺縁葉という名称に由来しており、主として比較解剖学的研究をベースにして導入されたものである。通常、辺縁葉といった場合に、帯状回、脳梁灰白層、海馬、歯状回、海馬支脚(または海馬台)、前海馬支脚、旁海馬支脚、嗅内野、前梨状皮質、中隔、嗅結節、扁桃核(とくに内側核と皮質核)などが含まれる。さらに、終板旁回(または梁下回)、眼窩面皮質後部、島前部、側頭葉極など細胞構築学的に古皮質に類似しているということで辺縁系の内に含める学者もいる。さらに詳しい説明については、小池上著"大脳辺縁系"を参照されたい(表1,表2,)。
このような種々の領域を解剖学的用語で一色にまとめ上げることは相当に無理があり、概念のない言葉が流行する結果となる。また、機能的意味合いをもたせた辺縁系という用語も当を得たものではなく、単に暗黙の推量があるだけである(Brodal,1981)。この点、大脳皮質連合領や網様体という用語のあいまいさと実によく通じている。辺縁系という用語は意味のない言葉となりつつあり、とくに本稿の表題のように形態学との関連のなかで使用されるとき然りである。しいていえば、辺縁系とは通常視床下部および中脳の一部と密接に(多シナプス性であるにしても)相互に結合しているワンセットの複合構造体であるといえよう。
それにしても、海馬(sea horse, hippocampus,ギリシャ、ローマの神話に登場する海神ポセイドン-別名ネプトウヌス-が乗る海の怪物ピッポカンポスの胴についている魚の尾の形に似ていることから名づけられた)といい、アンモン角(Ammon's
horn-海馬足pes hippocampi-その断面がアンモン貝の化石に似た形をしていることからつけられた名称、またアンモンはエジプト人の崇拝する太陽神)といい、扁桃核(または扁桃体、旧約聖書によくでてくる木の実アーモンドは悲嘆にくれたセラスの姫君の落涙結実物で涙の形をしている)にしろ、辺縁系の構成部分の領域に実によく神秘的名称が付けられているものである。
海馬(アンモン角ともよばれる)は原始皮質ないし古皮質(archicortex
or allocortex-old pallium)とよばれ、終脳の蓋板につづく半球内側面の部分が翼板の肥厚によって発生の早い時期に形成される大脳皮質の一部である。海馬は内部に閉じこめられているために外からはみえない。発生初期には脳梁の背側に位置しており、脳梁が背尾方に発達してくる時に同伴して次第に発達し、尾方では脳梁膨大の腹側で狭い小帯回gyrus
fasciolarisとなる。さらに腹側前方に進んで数珠玉を並べたような外観を呈する歯状回gyrus
dentatusとなり、また側脳室内に突出して海馬足pes hippocampi(固有の海馬、アンモン角)を形成する。海馬の尾方発達に伴い吻側部は退化し、完成された脳では単に脳梁の表層を被うだけの脳梁灰白層indusium
griseumとして残っている(内に内側縦条と外側縦条を入れる)。
いわゆる海馬はその発生初期から乳頭体と一部結合しているが、この結合部の内に割って入った形で新皮質の発達に伴って脳梁が膨大化し尾方に発達するので、この結合も伸張され脳弓fornixとよばれる弓状彎曲体(アーチ)が形成され、海馬足の上に海馬采fimbriaをつくって終わる。なお、左右の脳弓間には海馬交連が形成される。このように脳弓が迂回した経路になっているのは海馬が発生の途上で尾方へ引きずられたためである(図1)。
原始皮質archicortexは両生類において出現する。(固有の)海馬に歯状回および海馬支脚(subiculum,
およびその近傍領域)を含めて海馬体または海馬形成hippocampal
formationと一括してよばれることが多い。完成されたヒトの海馬は側脳室下角の床部に沿って前方・内方に折り込まれた形に入りこんでおり、海馬溝を被うようにその上に位置を占めている。その部分に相当して側脳室には凹みがある。したがって側脳室に接している面が最深層で脳室上衣層で被われた海馬白板alveusとよばれる白質(有髄線維の薄い層)から成っている。
海馬は細胞構築学的にみて特徴的であるが、一見貧弱かつ単純である。表層から深層(脳室面)へと順に、①(外)叢状叢、②錐体細胞層、③多形細胞層の3層構造から構成されている(図2)。それぞれ新皮質のⅣ、Ⅴ、Ⅵ層に相当すると考えられている。一般に叢状層に求心性線維が入り、錐体細胞層および多形細胞層から遠心性線維が出ている。一方、これに髄鞘染色が加味されると海馬の層構造は複雑となる。すなわち、側脳室の方から順に、①脳室上衣層、②海馬白板、③上昇(行)層str.oriensまたは内叢状層internal
plexiform layer,④錐体細胞層str.pyramidale,⑤放射(線)状層str.radiatum,⑥網状層str.reticulare
s.lacunosum,⑦分子層str.moleculareまたは外叢状層external plexiform layer,⑧内髄層str.medullare
involutaまたは帯状層str.zonaleに分けられる。なお、海馬cornu ammonis内の皮質分野区分名として、Lorente
deNo(1934)は海馬台側から歯状回側へ順にCAl(a,b,c)CA2,CA3(a,b,c),CA4に分けた。H1~H3(Vogt)やhlからh5(Rose)という区分法もある。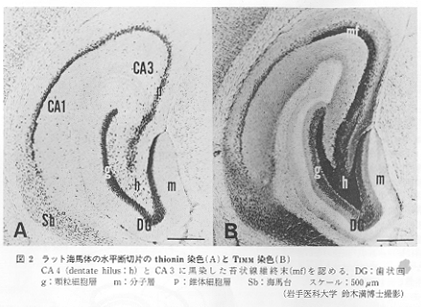
海馬形成hippocampal formation(海馬体)あるいは海馬領域hippocampal regionとして通常固有の海馬(hippocampus
proper,またはアンモン角)とともにまとめられる歯状回と海馬支脚(または海馬台:海馬を支える台のような外観を呈する)について以下に述べる。歯状回gyrus
dentatus(旧名fascia dentata)は動物の種による差が大きい領域であるが、海馬に連続して海馬溝の背壁にロシアのマトリューシカ人形のように内に隠れている痕跡的な脳回である。一方、海馬溝から腹方、外方に側副溝までつづく領域は広義の海馬台subiculum(とくに細分すればprosubiculum,
subiculum, presubiculum, parasubiculumとなる)と嗅内野(entorhinal area,28野)を含み、おおよそヒトの海馬旁回(旧名の海馬回を使用する人もありまぎらわしい)に相当する。組織学的には、歯状回は表層から深層へ、①辺縁層str.marginale,②分子層str.moleculare,③顆粒層str.granulosum,④多形細胞層または錐体細胞層と区分された明瞭な層構造を呈しており、この錐体細胞層は海馬のCA4の同名の層に移行している。他方、海馬旁回は皮質の幅も広くなり層的分化も進み6層構造を呈する古い皮質periallocortex(Lorente
deNo, 1934)に属し、その(おそらく)Ⅳ-Ⅴ層は海馬のCA1の錐体細胞層に連続している。なお、嗅内野は梨状葉皮質piriform
cortexの後部の大部をつくりあげている(図3)。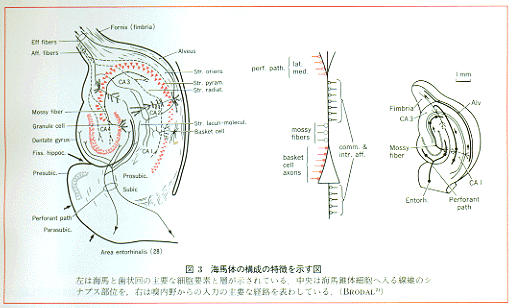
海馬体を東の横綱とすれば、西の横綱の位置を占めるのが扁桃体であろう。哺乳動物の扁桃核は、尾状核や被殻と同様に、半球胞の腹側壁が側脳室の内腔に隆起状に発達した神経節丘の後下部から生じる。側頭葉が形成されるにつれて神経節丘の腹側が前方に移動し、側脳室下角の前端の前上部に扁桃体が位置するようになる。ヒトの扁桃体は側頭葉前部の海馬旁回鈎(uncus,海馬旁回の前端が後外側に曲った部分)のすぐ下にみられる。一般に扁桃体と海馬は
"関係が深い"
と考えられているが、このように発生の過程を調べてみると、両者は互いに独立分離して発達し、たまたま最終的に定着した位置が比較的接近しているという理由が大きいのかもしれない。
扁桃複合体amygdaloid complexという全体名称があるように、扁桃核は構築の異なる幾つかのグループまたは亜核に分けられる。動物種により発達の程度が異なるのは当然としても、領域により必ずしも境界が明瞭でないこともあって、区分や命名法が研究者により多少異なる厄介な構造物である。従来から比較解剖学上嗅覚との関連で発達してきたと考えられている扁桃体(およびその周辺皮質部)の亜核間の関係が動物が高等になる程、複雑になっていることは興味深いし、また注目に値する(小池上1971)。
動物種間の相違はあっても、扁桃体にみられる一般的構成パタンは似ている。すなわち、小細胞群のことを抜きにすれば、皮質内側核群corticomedialと基底外側核群basolateralとに大別される。基底外側核群(基底核と外側核)は動物が高等化するにつれて発達し、ヒトで著明である。一方、内側核・中心核および皮質核は逆にヒトで発達が悪い。なお、鳥類以下の原始線条体(または嗅線条体)は哺乳類の扁桃体に相同とされている。
大脳半球の内側面で、左右の側脳室前角を分離し、脳梁と脳弓の間に垂直に張られた1対の薄い板(透明中隔板)の内にある灰白質の層板から構成されており、終脳由来の核である。その大部分の領域は前交連よりも前方を占めている。下等哺乳類でよく発達している。ヒトでは発達が悪く、中隔野の上部には神経細胞が殆どみられない透明中隔があり、その下部("precommissural"septumという)には外側核と内側核がみられる。その他の小さい細胞集団として、中隔海馬核(背側中隔核)・中隔海馬采・前交連床核・側坐核などをあげる研究者もいる。さらに、終板旁回(旧名、脳梁下回)や対角帯核などの皮質部も含めることがある。
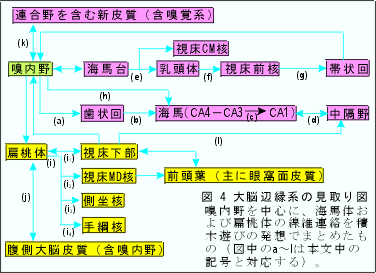
辺縁葉ないし辺縁系に関する専門家達の著書や研究論文を、形態学的なものだけに限っていくつか読んでみただけでも、海馬体(海馬・歯状回・海馬台)・扁桃体・視床下部(当然乳頭体も含む)・中隔野・嗅内野など(固有辺縁系領野limbic structures proper)の間の結合関係、さらには、大脳基底核、視床核の一部、中脳辺縁系野・側頭葉極部など(旁辺縁系領域paralimbic areas)まで含めた神経連絡路となる素人の目には迷路のように複雑である。まして研究者による、また研究方法による所見の相違や動物種間の差などを遂一考察してまとめあげることは困難である。小異を捨てて重要な点に注目して要述的にまとめてみたい。以下に、大脳辺縁系についての鳥瞰図(図4)を示し短い説明を付けておく。なお、神経路に関する全体的な知識は本号の "線維連絡" の章から得ていただきたい。
(a):嗅内野(皮質の2層と3層に神経線維の叢がある。詳しくは、細胞構築学的にも線維結合の上からも、内側部-28a野-と外側部-28b野-とに分けられる)からの内側および外側貫通線維perforant
path。海馬台を通り抜けて海馬溝を越えるのでこの名がある。主要なものは歯状回の顆粒細胞の尖端樹状突起が存在する分子層の外層(外側貫通線維)と中層(内側貫通線維)に終わる。exteroceptive
の情報を運ぶ。なお、内層には対側歯状回からの交連線維が終わる(海馬采からの線維とともにpsalterium,紘またはhippocampal
commissureとして入る)。
(b):歯状回の顆粒細胞からおこりCA4,CA3の錐体細胞の樹状突起に終わる苔状線維とよばれる線維。
(c):CA3と(おそらく)CA4(歯状回のhilus域)の大型錐体細胞の神経突起の分枝recurrent
collateralsがCA1の小型錐体細胞の樹状突起(網状層)に終わる。Schaffer線維という。
(d):主としてCA3よりおこり中隔の外側核に終わる。中隔・海馬路はコリン作働性線維を多く含み内側核からおこり海馬内の広範囲の領域(上行層)に終わる。interoceptive
の情報を運ぶ。
(e):いわゆる海馬乳頭体路で海馬台からおこり脳弓を通って乳頭体外側部に終わる。アンモン角(固有の海馬)からの投射はない。
(f):Vicq d'Azyr束ともよばれ乳頭体の主として内側部からおこる。細かくいえば、視床前核は乳頭体の内側核から同側性に、外側核から両側性に線維を受けている。
(g):視床前核群(とくにAM核、AV核)から帯状回皮質への投射には部位局在の関係が存在する。
(h):嗅内野および(おそらく)海馬台から白板線維alveus,alvear
pathとして海馬(主としてCAl)に終わる。貫通線維の一部も終わる。標的はバスケット細胞と(おそらく)錐体細胞の基底樹状突起。
(i):扁桃体からの皮質遠心性投射(発生的に古い皮質内側核群との結合が強い)
(i1):分界条stria terminalisおよび内側前脳束medial forebrain bundleを通る。視床下部腹内側核(VMH)に多く、また外側核(LH)にも終わる(他に視索前野、分界条床核にも終わる)。視床下部扁桃体投射は、主として皮質内側核群に終わる。
(i2):下視床脚inferior thalamic peduncleを通り、MD核の発生的に古い部分である内側部(大細胞性領域)に投射する。この投射域は嗅覚野が存在する眼窩面皮質との結びつきが強い。
(i3):連合縦束longitudinal association bundleを通る。一部が尾状核腹側部、中隔核にも終わるが主たる終止域は側坐核である。なお、側坐核は黒質内側部および中脳腹側被蓋域からドーパミン含有線維を受けており、被殻腹側部を介して中脳網様体の腹内側部(いわゆる中脳のlocomotor
region)へ、線条体からの線維とともに投射している。
(i4):下視床脚を通る弱い投射である。なお、手綱核は視床髄条を介して中隔核、視床前核、外側視床下部域、対角帯核、外側視索前野からの神経線維を受けている。また反屈束(手綱脚間路)を介して中脳の脚間核へ線維を送る。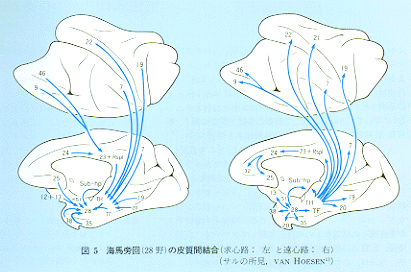
(j):扁桃体-大脳皮質間結合(発生的に新しい基底外側核との結合が強い)。側頭葉前部、前頭葉眼窩面皮質、帯状回と相互に結合する。
(k):(j)と同様な領域との相互結合の他に比較的広範囲の新皮質領域からも嗅内野への投射がみられる(図5)。前頭葉からの線維のうちかなりのものが帯状束内を通路とする。
(l):弱い結合が、おそらく存在する。
図4から読みとれるように、大脳辺縁系を、①海馬台-乳頭体系、②海馬-中隔系、③扁桃体-視床下部系の3系に分けることができる。①と②を海馬系としてまとめれば、③の扁桃体系と対比させられて2つに大別できよう。
1975年頃すでに、小池上は当時の一般的見解として次のように述べている。すなわち、"辺縁系は新皮質と対立するいわゆる古い脳部を代表するものであり、自律神経系の最高中枢である視床下部に対し、ある程度の制御を与えているものであり、種族保存・自己保存などの本能的な機能ととくに関連が深く、また情動行動や記憶にも関係し、臨床上てんかん発作との関連が重視せられ、その他いろいろの関係から脳の基本的構造上、新皮質-脳脊髄系と対立せしめて考えるべき重要なものと考えられるようになった。視床下部-自律神経系(視床下部およびより下位の自律神経中枢とその末梢を綜括する)と、脳幹網様系(視床汎性投射系をふくむ)に対して、辺縁系は概して前者の系統と関係が深く、新皮質-脳脊髄系は後者すなわち脳幹網様系と関連が深い。"
10年後の今日、この見解に何をつけ加え、どこを訂正し、どのようにより正確に記載することができるであろうか?モノアミン系投射線維(帯状束と脳弓を通って海馬に入る。なお、歯状回には至らないようである)、ニューロンの活性物質(GABA・ソマトスタチン・CCK・ニューロテンシンなど)、シナプスレベルでの結合、さらに嗅覚系や脳幹や大脳基底核との関連などの知識が集積してきている。本能、情動、記憶との密接な結びつきなど、いわれてから久しいが、今日は、これらの心理学的言葉は徐々に生理学的、生化学的用語におきかえられようとしている。形態学的に調べてみて大脳辺縁系は、一方で終脳の新皮質(
"連合野" も含めて)との相互結合関係が意外に強く(van
Hoesen1982)、他方では視床下部との結びつきも強いという複雑な特異なシステムであるように思われる。新皮質から海馬体への入力は必ず嗅内野・海馬台(ヒトでいう海馬旁回parahippocampal
gyrus)を介している。これは連合線維(皮質-皮質間結合)のチェインである。皮質間の相互結合という点からみたとき、固有の海馬ないし海馬体は、"大脳の辺縁"
の奥まった所にあって視床下部を含めた脳幹部から入力される生存に必要な要素と連合野からの高度な情報を一時的にでも結びつける事により、一般記憶の記銘過程や空間記憶の保持に関連した作業に関与しているらしい。これを支持すると思われる行動・生理学的な証拠も提示されている。他方、扁桃体と大脳皮質との関係は、海馬体とは対照的に、連合線維系の結合ではなく投射線維系である。扁桃体は側頭葉極や下部側頭葉皮質や前頭葉の腹側部および眼窩面皮質など、感情とか情緒とかに直接または間接的に関連すると思われる皮質と比較的強く相互に結合している。その上、扁桃核は味や臭いや自律神経系統の皮質下核と結合しており、亜核内(発生的に新しい部分と古い部分がある。前述)での役割分担も示唆されており、皮質-扁桃核間の神経回路が働くことによって、賞罰、報酬などの意味づけ、動機づけ(ないし連合表出)などの形成がなされる所なのであろう。このようにみてくると、大脳辺縁系は大別して、①記憶変換器としての海馬体系と、②感情表出複合体としての扁桃体系とから構成されているとみなしてよいだろう。しかしながら、両者とも量質の差があるにしても、ともに大脳皮質と脳幹部からの入力(感覚系についていえば単に嗅覚系のみならずすべての種類のものが入ってくると考えられる)に制御ないし調節されており、しいて2系の働きを結びつけようとするならばその主要な接点は嗅内野ないし海馬旁回(海馬台を含めて)にあるのかもしれない。上に述べたような神経回路ないしシステムから構成される辺縁系の行動・生理学的研究に立脚して、Mishikin(1982)のグループは視覚再認記憶に関する連合野・扁桃体・海馬・視床を包含した仮説的なモデルを提唱していることを特記しておく。
追記:著者校正後に、海馬と扁桃体の間に以前考えられていたより広範な相互結合がサルで存在するという研究論文(Exp.Brain
Res., 64/3, November, 1986, 515~526)が現れた。今後の研究の一つの焦点と思われるので注目したい。
Clinical neuroscience 5/2 (1987) 132-138 ,
中外医学社、より許可を得て転載