車田正男、川村光毅
| A]はじめに | |
| B]上丘の形態学 | |
| 1)神経細胞 | |
| 2)神経網 | |
| C]上丘の線維連絡 | |
| 1)上丘浅層 | |
| 2)上丘深層 | |
| 文献 |

中脳蓋(tectum mesencephali)は中脳の背側の領域を占め、上丘(colliculus
superior)と下丘(colliculus inferior)からなる。ここでは視覚、聴覚の情報と、顔面、体幹、四肢などからの外来性の感覚情報とが互いに干渉し合っている。
中脳蓋は系統発生学的にさまざまな形や大きさを示す。この形態的差異は動物の種の違いによるばかりでなく同一種内(硬骨魚類でとくに著明)においても認められる。これらはそれぞれの動物の習性の違いによるものと考えられる。
下等な脊椎動物の中脳には側線中枢というものがあり、そこでは側線からの情報が他の種類の感覚情報と関連している。より高等な、たとえば両生類以上の脊椎動物になるとこの側線中枢は蝸牛中枢に置きかえられ、さらにこれらの動物の中脳には聴覚情報と他の情報を相互に関連されるための諸領域が出現してくる。爬虫類の一部や哺乳類になると、聴覚の中枢は中脳のより背側に位置するようになりやがて下丘を形成する。
視蓋(tectum opticum)(哺乳類では上丘)はすべての脊椎動物(ナメクジ魚では欠除)において、中脳の背側に位置する層をなした構造として認められる。視蓋は元来視覚の中枢として発達してきたもので、哺乳類以下の動物では視神経線維のほとんどすべてがここに終る。さらに視蓋には脊髄、三叉神経核、側線・聴覚領域からもきわめて多くの線維が終っている。したがって、視蓋は触覚や前庭・聴覚領域などの外来性インパルスの統合中枢であるともいえる。しかし、哺乳類では大脳皮質の発達に伴い視覚の最高中枢は皮質視覚領にとってかえられ、上丘には主として眼と頭の協同運動に関係する視覚性反射機能が残るようになる。
視蓋の構造(各層の厚さや細胞構築など)はそれぞれの動物の違いによって多少差があるが、哺乳類以下の脊椎動物では(硬骨魚類を例にあげると)表面から深部に向かって以下のように区別される(Schroederら、1980)。
1) 帯(状)層(stratum zonale or marginale):縦隆起(torus
longitudinalis)からの線維が終止するところ。
2) 視神経層(s. opticum):視神経線維や膝状体-視蓋線維からなる。
3) 浅線維層および浅灰白層(s. fibrosum et griseum
superficiale):視神経線維をはじめ脊髄・視蓋線維や橋・視蓋線維が主に終るところで、局所回路ニューロン(local
circuit neuron)と思われる細胞も多く存在する。
4) 中心(または深)線維層(s. griseum centrale or
profundum):視蓋遠心路は主にこの層の細胞からおこる。
5) 中心(または深)線維層(s. fibrosum centrale or
profundum):視蓋遠心線維をはじめ両側視蓋を結ぶ交連線維からなる。
6) 脳室周囲層(s. periventriculare):視神経層にまで達する長い上行性の樹状突起をもつ神経細胞よりなる。
一方、哺乳類の上丘は基本的には下等な脊椎動物のものとよく似た層構造を示すが、いくつかの差違もみられる。もっとも著明なことは、視神経層の表層に新たな灰白質の層(浅灰白層)が一つ加わることである。これは視覚領をはじめとする大脳皮質の発達と関連している。
ここでは主にGolgi標本と電顕から得られた所見を中心に、哺乳類(とくにヒトとネコ)の上丘の構造を示し、ついで実験的研究により明らかにされた上丘の神経連絡について述べる。
上丘は主に神経線維あるいは神経細胞からなる七つの層より構成される。上丘の表面から1)帯層(s. zonale),2)浅灰白層(s. griseum superficiale), 3)視神経層(s. opticum),4)中間灰白層(s. griseum intermediale),5)中間白層(毛帯層)(s. album intermediale or s. lemnisci),6)深灰白層(s. griseum profundum),7)深白層(s. album profundum)である(Kanaseki and Sprague,1974)。これらの層は、行動学的立場(Casagrande and Diamond, 1974)から帯層~視神経層からなる浅層とそれより深部の深層に大別される。また、形態的にも浅層と深層との間には著明な差違が認められる。たとえば、Nissl染色による上丘の標本を観察してすぐに気づくことは、浅層にみられる神経細胞は大部分が小型で少数の中型細胞が混在しているのに対して、深層には大小さまざまの大きさの細胞が存在している。したがって、Nissl標本で観察した限りでは浅層の神経細胞は同質性(homogeneous)で、一方深層の神経細胞は異質性(heterogeneous)であるという特徴を示す(図8.1b, c)。
a)Nissl標本による観察
上丘浅層をKluver-Barrera染色標本(図8,1)で観察すると、軟膜の直下には幅が約30μmの神経膠細胞(大部分が星状膠細胞)の層があり、その深部にほぼ同じ幅の有髄線維を主体とする層がみられる。この層は帯層と呼ばれ、神経細胞の数は少ない。この層にみられる細胞の大部分は淡染する小型の細胞である。帯層の深部には神経細胞を主体とする浅灰白層がある。この層にはいろいろの形をした小型(図8.1
bのs)および中型(図8.1 bのm)の神経細胞が存在する。この層の上部には小型細胞が多くみられるが、下部では中型細胞が主に観察される。KanasekiとSprague(1974)によりこの層はさらに三つの亜層に区分されている。視神経層の大部分は有髄線維でしめられている。神経細胞は小型のものが主であるが、そのなかに中型の細胞も混在している。
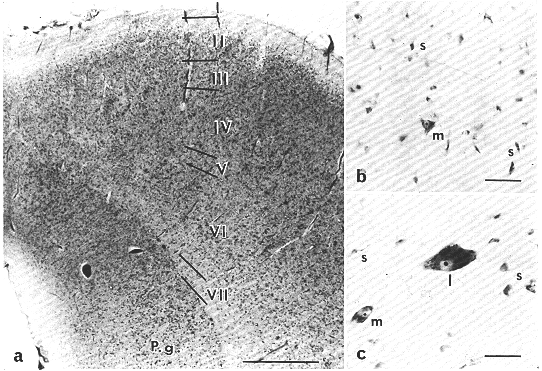
図8.1 a:ヒト上丘の層構造。 Ⅰ::帯(状)層、Ⅱ:浅灰白層、Ⅲ:視神経層、Ⅳ:中間灰白層、Ⅴ:中間白層、Ⅵ:深灰白層、Ⅶ:深白層、P.g.:中心灰白質。[スケール:1mm]
b:浅灰白層の神経細胞。 小型細胞(s)に混じって中型細胞(m)もみられる。[スケール:50μm]
c:中間灰白層の細胞。大型(l)、中型(m)、小型(s)がみられる。[スケール:50μm](旭川医科大学解剖学教室佐藤洋一博士所蔵のKluver-Barrera染色標本による)
視神経層の直下には主として神経細胞からなる中間灰白層がある。この層にみられる神経細胞の大部分は小型ないし中型の細胞で、これらに混って大型の細胞も散在する。この大型の細胞は多極性の細胞体を有しニッスル小体がきわめてよく発達している(図8.
1 c)。中間白層は主に有髄線維からなる層で、これらの線維の間には小型~中型細胞が散在している。深灰白層には神経細胞が多数存在する。この層にみられる神経細胞の形態は中間灰白層みられるものとよく似ているが、後者に比べると細胞密度がいくぶん低い。上丘の最深部に位置する深白層は主に有髄線維からなる薄い層であり、小型の神経細胞に混って中型の細胞が散在性にみられる。
b)Golgi標本による観察
Golgi染色切片上でさまざまな形をした神経細胞が上丘浅層に認められる(図8.2)。これらの神経細胞を、樹状突起およびそれらによって形づくられる樹状突起野の位置・方向(orientation)を基準にして分類すると、水平(型)細胞(horizontal
cells),垂直(型)細胞(vertical cells),多極(型)(multipolar cells)に大別される。
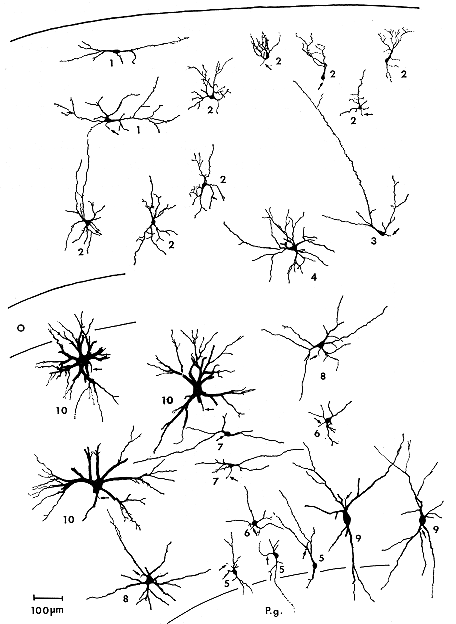
図8.2 ネコ上丘神経細胞の分布を示す模式図
浅層:水平(型)細胞(1)、垂直(型)細胞(2)、樹状突起野の比較的広い垂直細胞(3)、中型の多極(型)細胞(4)。
深層:小型の垂直細胞(5)、多極細胞(6)、水平細胞(7)、中型の多極細胞(8)、垂直細胞(9)、大型多極細胞(10)。
O:視神経層、P.g.:中心灰白質。矢印は軸索を示す。(迅速Golgi染色標本による)
水平細胞(図8.2の1)は小型で、多くは紡錘形の細胞体を有しその樹状突起は上丘表面をほぼ平行に走る。樹状突起の長さは1,000μmに達することもある。水平細胞のほとんどすべては上丘浅層の上半域に存在する。なお、この種の細胞がシナプス前樹状突起(presynaptic
dendrite:PSD)を有することはよく知られている(後述)。
垂直細胞(図8.2の2)は垂直方向に伸びた円柱形の樹状突起を有する小型あるいは中型の神経細胞である。細胞体は通常垂直方向に長軸をもつ紡錘形を呈する。この細胞は浅層のほぼ全域に存在し、一般により深層にみられるものほど形が大きい傾向にある。さらに、この垂直細胞の亜型ともいえる広い樹状突起野をもった細胞(図8.2の3)も浅灰白層の深部から視神経層にかけてみられる。この細胞の樹状突起はほとんどのものが表層に向かって広がっている。
多極細胞(図8.2の4)は小型および中型の細胞で多極性の細胞体を有する。樹状突起は細胞体からあらゆる方向に広がり、樹状突起野はほぼ球形をなす。中型の多極細胞は視神経層に多くみられる。さらに、以上3種の神経細胞のほかに上丘の表層にのみ限局するmarginal
cellsがある。これはきわめて小型の細胞で樹状突起は短くそして微細である。
他方、上丘深層にも数種類の神経細胞が観察される。これらの細胞は細胞体の大きさに基づいて、大型細胞(35~60μm)、中型細胞(20~30μm)、小型細胞(8~15μm)、の3種類に大別される。
大型細胞(図9.2の10)の特徴は、細胞体が多極形を示し細胞体や樹状突起に多くの棘を有することである。この細胞の樹状突起野はほぼ球形をなし、通常直径400μm以上にも広がっている。したがって、一つの細胞から発した樹状突起が二つか三つの層にまたがることもまれではない。大型細胞は上丘深層の外側2/3の領域に多くみられる。
中型細胞は多極細胞と垂直細胞からなり、中程度の数の棘を有する。中型多極細胞(図8.2の8)は径350μmの球形の樹状突起野を示し深層全域に分布している。中型垂直細胞(図8.2の9)は径350μm以下で長さ500~600μmの垂直方向に伸びた円柱状の樹状突起野を有する。この細胞の深層の内側2/3の領域に多く分布している。
小型細胞は多極細胞、垂直細胞、水平細胞からなり、きわめて少数の棘を有している。小型多極細胞(図8.2の6)は深層全域に観察され、径250μm以下の球形の樹状突起野を有する。小型垂直細胞(図8.2の5)は主に深灰白層および深白層にみられ、樹状突起野は径150μm以下、長さ約300μmの円柱状を呈する。水平細胞(図8.2の7)は中間灰白層にほぼ限局して観察されるが、浅層にみられる同名の細胞とよく似た形態を示す。この細胞の樹状突起野は水平方向に軸をもつ幅150μm以下、長さ400~1,000μmの円柱形を呈する。
c)電子顕微鏡による観察
上丘浅層における神経細胞は電顕で観察すると小型細胞と中型細胞の2種に大別される。
小型細胞(図8.3a)は細胞質に乏しく相対的に大きな核を有している。細胞質内には遊離したリボゾームが目立ち、ニッスル小体はほとんど発達していない。細胞体に終止する神経終末の数は少ない。これらの小型細胞のうちで、細胞質や樹状突起の中に多形性のシナプス小胞を含むものも観察される。このような特殊な細胞はPSD細胞とも呼ばれており、上丘浅層では水平細胞がこれに相当すると考えられている(Valverde,
1973;Tigges and Tigges, 1975)。

図8.3 ネコ上丘浅層にみられる小型細胞(a)と中型細胞(b)の電顕像[スケール:3μm]
N:核、NB:ニッスル小体
一方、中型細胞(図8.3b)の細胞質は比較的豊富な細胞内小器官を有する。核周囲部は大部分グリア性の層板によっておおわれており軸索細胞体間シナプスはわずかにみられるのみである。その他の特徴には乏しい。
上丘深層の神経細胞は、その細胞質やシナプス結合の特徴から3種類(大型、中型、小型細胞)に区別され、さらにそれらはGolgi標本によって分類された3種類の神経細胞とほぼ対応する(Norita,
1980)。
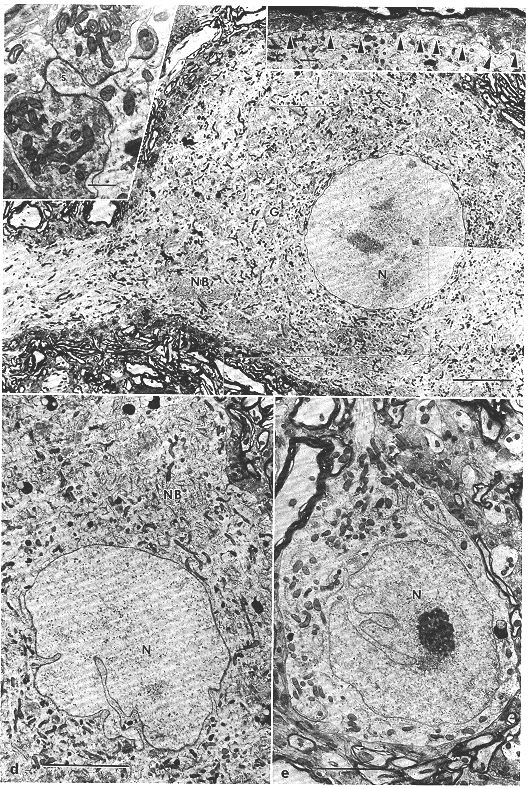
図8.4 ネコ上丘深層ニューロンの電顕像
a:大型細胞。[スケール:10μm]
b:大型細胞の細胞体にみられる棘(s)。この突起には3個の軸索終末が終止している。[スケール:0.5μm]
c:大型細胞の表面は多数の終末ボタンにより覆われている(矢尻印)。[スケール:1μm]
d:中型細胞。核膜の陥入が著明。[スケール:5μm]
e:小型細胞。核膜には陥入がみられ、細胞質は乏しい。G:ゴルジ装置、N:核、NB:ニッスル小体。[スケール:3μm]
大型細胞(図8.4a)はきわめてよく発達した細胞内小器官を含む径35μm以上の細胞体を有し、核膜陥入のほとんど認められない円形の核をもっている。さらにこの細胞に特徴的なことは、細胞体周囲の70%以上がいくつかのタイプの終末ボタン(後述)でおおわれている(図8.4c)ことと、細胞体からの小突起がしばしばみられることである。この小突起には通常二つ以上の終末ボタンがシナプスしている(図8.4b)。
中型細胞(図8.4d)は多極性あるいは紡錘状の細胞体と核膜陥入の著明な核を有する。細胞質内には比較的よく発達したニッスル小体、糸粒体(ミトコンドリア)、ゴルジ装置、リポフスチン顆粒、ライソゾームなどが分布している、細胞体表面の約25~30%は終末ボタンにおおわれており、さらに細胞体からの小突起がみられることもある。
小型細胞(図8.4e)も中型細胞と同様に多極性または紡錘状の細胞体を有する。細胞体の大部分は核によってしめられており、この細胞質内にはあまり発達のよくない粗面小胞体をはじめとする細胞内小器官が散在している。小型細胞はあまり多くの終末を受けておらず、細胞体の表面の10%以下が終末ボタンにおおわれているにすぎない。
a)上丘浅層
上丘浅層にみられるシナプス前要素は少なくとも3種類、つまり1)水平細胞からおこることが確かめられているPSD,
2)視神経終末、そして3)その他の一般的によくみられる軸索終末、に分類される。これらシナプス前要素と上丘ニューロンとの間にはいろいろのタイプのシナプス結合が形成されるが、大部分のシナプス結合は細い樹状突起やそれらの棘にみられる。上丘浅層におけるシナプス結合の特徴として、きわめて多くの連続シナプス(serial
synapses)の存在をあげることができる。
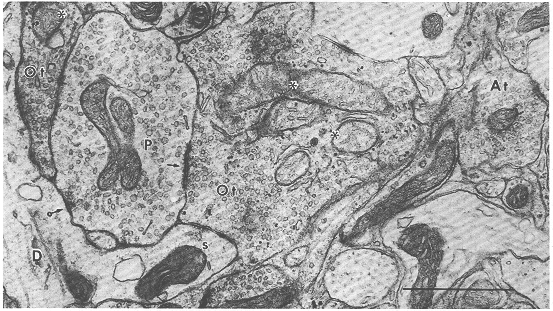
図8.5 ネコ上丘浅層のシナプス前要素の電顕像
PSD(P)、視神経終末(Ot)、一般の軸索終末(At)の3種類のシナプス前要素がみられる。明るい基質の糸粒体(星印)を有している視神経終末は樹状突起の棘(s)とPSDにシナプスしている(矢印)。このPSDはさらに一般の樹状突起(D)にシナプス(環矢印)している(軸索・樹状突起・樹状突起の連続シナプス)。[スケール:1μm]
PSD(図8.5のP)は比較的明るい基質を有し、その内にはまばらな多形性のシナプス小胞、滑面小胞体、暗調な糸粒体などがみられる。さらに、このPSDのもっとも重要な特徴はその細胞質内にしばしばリボゾームの集積がみられることである。この終末は視神経終末や大脳皮質視覚領からの終末に対してはシナプス後要素であり、一般の樹状突起に対してはシナプス前要素となる。したがって、PSDを中心として軸索・樹状突起・樹状突起の連続シナプスが形成されるわけで、この連続シナプスは浅灰白層の上部に特に多くみられる。
視神経終末(図8.5のOt)は比較的大きく不規則な形をした終末である。この終末内には多数の丸いシナプス小胞や、明るい基質をもったややふくらんだ糸粒体(図8.5の星印)がみられる。この糸粒体の形態は上丘浅層における視神経終末の最大の特徴である。この終末は浅灰白層の上半域にもっとも多く認められる。
上丘浅層にみられる第三のシナプス前要素は一般によくみられる終末(図8.5のAt)であり、浅層に存在するさまざまな神経細胞や他の領域(たとえば大脳皮質や傍二丘体など)からの軸索終末である。これらのうち、小型の終末で丸いシナプス小胞を有し非対称性膜肥厚を示すシナプス結合を形成するものの一部は、大脳皮質視覚領からの終末であることが実験的に明らかにされている(Valverde,
1973)。
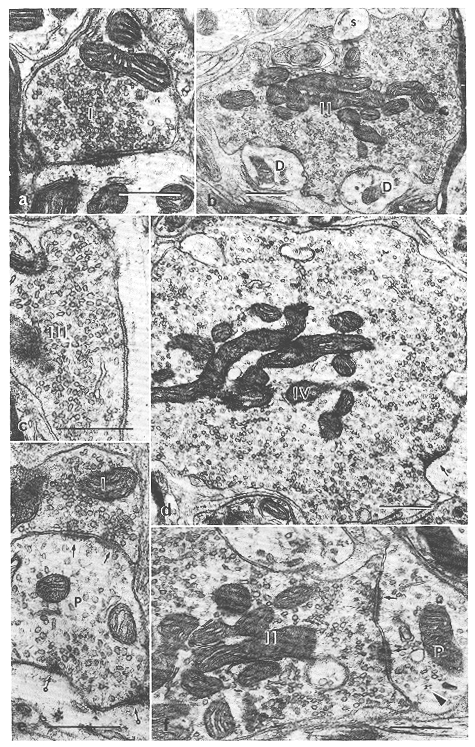
図8.6 ネコ上丘深層のシナプス前要素の電顕像[スケールはすべて0.5μm]
a:Ⅰ型軸索終末(Ⅰ)。樹状突起にシナプスしている。
b:Ⅱ型軸索終末(Ⅱ)。二つの細い樹状突起(D)と棘(s)にシナプスしている。
c:Ⅲ型軸索終末(Ⅲ)。樹状突起にシナプスしている。
d:Ⅳ型軸索終末(Ⅳ)。大型細胞の近位樹状突起にシナプスしている(矢印)。
e:連続シナプス、Ⅰ型終末(Ⅰ)がPSD(P)と非対称型シナプス(矢印)を形成している。後者はさらに一般の樹状突起と対称型シナプス(環矢印)をつくる。
f:Ⅱ型終末(Ⅱ)は非対称性膜肥厚(矢印)を形成しながらPSD(P)にシナプスしている。矢頭印はリボぞおーむを示す。
b)上丘深層
上丘深層にはきわめて多種のシナプス前要素が観察されるが、形態的に少なくとも8種類に区別される。軸索終末が7種類(I~VII型)で他はPSDである。
I型終末(図8.6a)はしばしばみられる径1.5μm以下の小型の軸索終末で、主に細い樹状突起や大型細胞の細胞体との間で非対称型のシナプス結合を形成している。この終末は時としてPSDに終止することもある(図8.6e)。I型終末の一部は大脳皮質からの終末であろうと思われる(後述)。
II型終末(図8.6b)は径3μm以下の中型の終末でしばしば観察される。これは主に中程度以下の太さの樹状突起やPSDとの間で非対称型のシナプス結合を形成する(図8.6f)。
III型終末(図8.6c)はもっともうよくみられる終末で、通常1.5μm以下の小型の終末像を示す。また、時として細長い形態を示し長さ6μmに達することもある。この終末はPSD以外のあらゆるシナプス後要素との間で対称型のシナプスをつくる。
IV型終末(図8.6d)は径約3.5μmの比較的大きな終末である。これは主に大型細胞の細胞体や近位樹状突起との間で対称型のシナプスを形成する。
上丘深層には浅層でみられたと同様なPSDが観察される(Norita and
Sugiyama 1979;Norita, 1980)。このPSDに終止する軸索終末はI型よII型の終末であるが(図8.6
e, f,)、主にII型終末が終る。深層におけるこのPSDがどの神経細胞からおこるのかは現在のところ不明である。なお、V型、VI型、VII型終末は出現頻度が比較的低い終末である(Norira,
1980)。
上丘は機能的な面ばかりでなく神経連絡の特徴からも浅層と深層に大別される。浅層は網膜や大脳皮質視覚領など視覚に密接に関係する領域から直接の入力を受け、外側膝状体や視床後外側核へ線維を出している。これに対して深層は網膜からの直接の入力を受けておらず、脊髄から大脳皮質に至るきわめて広範囲の領域から線維を受けている。深層からは視床膝状体上核などへの上行路のほか、橋、延髄、脊髄への下行性線維がおこる。
解剖学的研究により、上丘浅層は網膜、大脳皮質視覚領、視蓋前域、傍二丘体などから多くの線維を受けることが明らかにされている。網膜からの線維の多くは浅灰白層の上半域に終り、その終末は集積して規則的な斑点構造を形成する傾向がある(Graybiel,
1975)。一方、皮質視覚領からの線維は主に浅灰白層の下半域に終り、視神経終末のようにはっきりとした斑点状の終末の集団をつくることはない。
最近、horseradish peroxidase(HRP)を用いて研究によって、上丘浅層に存在する投射ニューロンの形態や分布が明らかにされてきた。たとえば、外側膝状体へ投射する神経細胞は主に浅灰白層の上半域に分布しており、その細胞体は小型で垂直方向に長軸をもつ紡錘形を示す。このような投射ニューロンの形態はGolgi標本で観察される小型の垂直細胞とよく似ている。これに対し視床後外側核へ線維を出す細胞はほとんどすべてが浅灰白層の下部1/3域にみられる。この細胞は視蓋・膝状体細胞に比べてやや大型で多形性を示す。これはGolgi標本における広い樹状突起をもった垂直細胞と類似する。これらのことから、網膜からの入力は視蓋-膝状体ニューロンの分布する領域に終止し、皮質視覚領からの線維は視蓋・後外側核ニューロンのみられる部位に主として終るように思われる(Graham
and Casagrande, 1980)。
上丘の深層は大脳皮質の広い領域から線維を受けている(Konno, 1979)ほか、きわめて多くの皮質下領域からの入力を受けている(Edwardsら、1979)。ネコの大脳皮質を広範囲に破壊して上丘深層における変性終末を電顕で観察してみると、ほとんどの変性終末は小型の終末で細い樹状突起との間で非対称型のシナプス結合を形成していることがわかる。これらの変性終末の形態はI型終末の形態とよく一致する(図8.7a)。
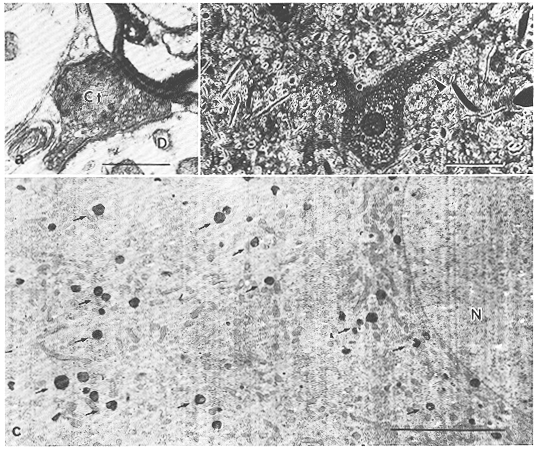
図8.7
a:ネコ上丘中間灰白層にみられた皮質-視蓋変性終末(Ct)。樹状突起(D)との間で非対称型のシナプス結合を形成している。[スケール:0.5μm]
b:HRP法により同定されたネコ上丘の中心灰白層の大型の視蓋-網様体投射ニューロン。多くのHRP果粒が細胞体内ばかりでなく近位樹状突起(矢尻印)にもみられる。[スケール:50μm]
c:Bの大型細胞の電顕像。細胞体内は多数のHRP顆粒(矢印)がみられる。N:核。[スケール:5μm]
つぎに、オートラジオグラフィーを用いた研究により黒質網様部からの線維は中間灰白層にほぼ限局して終り、この終止パターンは浅層にみられた網膜からの終末と同様に斑点状を示すことが知られている(Graybiel,
1978)。黒質からの線維はまた、GABAを神経伝達物質としているらしいことも示されている(Vincentら、1978)が、この終末の形態は先のIII型およびIV型終末と類似している。また、三叉神経知覚核からの線維も中間灰白層に斑点状に終止している(Tashiroら、1980;Ogasawara
and Kawamura, 1982)。このような特異な終止形態が示す入力が何を意味しているのか今後の課題である。
他方、HRP法を用いた研究により、深層における投射ニューロンの形態とその分布が明らかにされてきた。たとえば、脳幹網様体へ投射するニューロンは大型(40μm以上)、中型(25~40μm)、小型(10~25μm)の細胞であり、これらのすべては完全に深層のみに限局して分布している(Kawamura
and Hashikawa, 1978)。その他。オリーブ核、脊髄、外眼筋支配運動核周囲領域などに投射する細胞もほぼ同様な態度をとる。一方、橋核への投射ニューロン(Hashikawa
and Kawamura, 1977)および上丘の交連細胞は小型と中型の細胞で、深層のみならず視神経層にも分布している。図8.7
b.cはHRPで標識された大型の視蓋-網様体細胞である。このようにして標識細胞の微細構造を調べてみると、電顕的に分類されたニューロンをHRP電顕法を用いて同定することができる。
中脳蓋とくに上丘は、種々の形態的特徴をもった神経細胞(上丘内結合に関与する"局所回路ニューロン"と"上丘遠心性ニューロン"とが存在する)と上丘内の浅層とか深層とかの一定の領域内に終止する種々の求心性線維が織りなす複雑な神経網をもったいわゆる層構造を呈する中脳の背側にみられる隆起した構造物である。
機能的役割としては知覚性視覚系(上行性)と運動性視覚系(下行性)とを第一義的なものとみなしえようが、体性感覚や聴覚など他の感覚様態の機能との関連も重要な研究課題であり、これらの問題を具体的実験データを提示して解明していくことが今後望まれる。将来、上丘の形態がより詳細に研究されていく過程で、その機能的役割が一層明らかにされていくと思われる。
Casagrande, V.A. and Diamond, I.T.: Ablation study of the superior colliculus in the tree shrew(Tupaia glis).J. Comp. Neurol., 156:207-238, 1974.
Edwards, S.B., Ginsburgh, C.L., Henkel, C.K. and Stein, B.E.:Sources of subcortical projections to the superior colliculus in the cat. J. Comp. Neurol., 184:309-330, 1979.
Graham, J. and Casagrande, V.A.: A light microscopic and electron microscopic study of the superficial layers of the superior colliculus of the tree shrew(Tupaia gris).J. Comp. Neurol., 191:133-151, 1980.
Graybiel, A.M.: Anatomical organization of retinotectal afferents in the cat:an autoradiographic study. Brain Res., 96:1-23, 1975.
Graybiel, A.M.: Organization of the nigrotectal connection:an experimental tracer study in the cat. Brain Res., 143:339-348, 1978.
Hashikawa, T. and Kawamura, K.:Identification of cells of origin of tectopontine fibers in the cat superior colliculus.An experimental study with the horseradish peroxidase method. Brain Res., 130:65-79, 1977.
Kanasaki, T. and Sprague, J.M.: Anatomical organization of pretectal nuclei and tectal laminae in the cat. J. Comp. Neurol., 158, 319-338, 1974.
Kawamura, K. and Hashikawa, T.: Cell bodies of origin of reticular projections from the superior collicules in the cat. An experimental study with the use of horseradish peroxidase as a tracer. J. Comp. Neurol., 182:1-16, 1978.
Konno, T.: Patterns of organization of corticotectal projection of cats studied by means of the anterograde degeneration method. J. Hirnforsch., 20:433-444, 1979.
Norita, M.: Neurons and synaptic patterns in the deep layers of the superior colliculus of the cat. A Golgi and electron microscopic study. J. Comp. Neurol., 190:29-48, 1980.
Norita, M. and Sugiyama, M.:Presynaptic dendrites and serial synapses in the intermediate and deep layers of the cat superior colliculus. Neurosci. Lett., 11;161-164, 1979.
Ogasawara, K. and Kawamura K.:Cells of origin and terminations of the trigeminotectal projection in the cat as demonstrated with the horseradish peroxidase and autoradiographic methods. Okajimas Folia Anat. Jpn., 58(4-6):247-264, 1982.
Schroeder, D.M., Vanegas, H. and Ebbesson, O.E.:Cytoarchitecture of the optic tectum of the squirrelfish, Holocentrus J.
Comp. Neurol., 191:337-351, 1980.
Tashiro, M., Kudo, M. and Kawamura, S.:Discontinuous spatial distribution of the tectal afferents from the
trigeminal nucleus in the cat. Neurosci. Lett., 20:249-252, 1980.
Tigges, M. and Tigges, J.:Presynaptic dendrite cells and two other classes of neurons in the superficial layers of the superior colliculus of the chimpanzee. Cell Tissue Res., 162:279-295, 1975.
Valverde, F.:The neuropil in superficial layers of the superior colliculus of the mouse. A correlated Golgi and electron microscopic study. Z. Anat. Entwickl.-Gesch., 142:117-147, 1973.
Vincent, S.R., Hattori, T. and McGeer, E.G.:The nigrotectal projection. A biochemical and ultrastructural characterization. Brain Res., 151:159-164, 1978.
人体組織学・神経(編集:小川ら) 8.中脳、288頁―298頁。
朝倉書店(1984年)より許可をえて転載