条件反射と高次機能(Conditioned reflex and higher functions of CNS) |
川村光毅
サマリー
デカルトによって最初に提示された「反射」の概念がどのように変遷し、発展してきたかを問いかけ、条件反射の成立とその発展の経過をたどりながら、シェリントン、セーチェノフ、パブロフが提唱した概念について考察しようと試みた。ケプラー / ガリレオによって築かれていた当時の最新の科学である力学的法則を背景に組み立てられた17世紀の「反射」の概念は、時代と共に進歩する実験科学の研究の成果によって、脊髄反射の研究から高次神経活動の分野にわたるまでに拡大されるに至った。以上をベースにして、現在われわれは、精神活動のメカニズムの解析に挑もうとしている。

デカルトの「反射」の概念は、科学的実験に裏打ちされた思考によって、その後大きく発展した。すなわち、脊髄レベルの反射弓から脳幹レベルの複合反射弓へ、さらに、前脳とくに大脳皮質を取り込んで大きく質的に発展を遂げた概念として、「脳の反射」、「条件反射(感覚信号系から言語信号系へ)」の段階に立ち至っている。また、「全身体的反射」として、内分泌系や免疫系を含む液性伝達系をも視野に入れたとき、セーチェノフ / パブロフが確立した条件反射(一般にネルビズムと称されている)は、さらに発展した局面として「情動」の範囲をも包括するに至る。近年、神経科学や分子生物学が著しい発展をとげているが、その立場から条件反射による獲得形質の種内世代間の「継承」や進化の問題や、さらに、条件反射第二信号系(言語信号系)の成立を可能にした前脳形成、新皮質形成、言語関連領野などの問題について考察してみたい。
1. デカルト(Rene Descartes, 1596-1650)の反射の概念と機械論――評価と批判
反射という概念を最初に確立したのはデカルトであった。外界の刺激が神経の興奮の伝導によって脳に達し、その結果として起こる生物体の自動的反応を彼は反射と名づけた。デカルトは、精神と物質との二つの実体があるとする二元論に立ち、動物は複雑な自動時計であるいう機械論的自然観に立っていた。彼は動物精気という液体あるいはガスが脳室に満たされており、脳室の壁には気孔があり、そこをとおって動物精気 (spiritus animales) が中空の構造をもった神経へと入っていくと考えていた。一方感覚についてはそれぞれの神経管の真中に一本の糸があり、それは感覚器官から脳まで伸びていると考えた。現在反射とわれわれがよんでいるものの素朴な機械論的説明をここにみる。このように、デカルトは新しい自然観を構築し、自然界の現象を力学的法則によって運動しつづけるものと考えた。彼は理性を重視し学問における真理を求め、光学、幾何学、力学、化学、生理学、解剖学、医学、数学と広い自然の諸分野を考究した。かれは、中世のスコラ学的思弁を排し、純粋直截な精神の働きイコール直観にあたえられる明晰な観念を出発点とした。そして、数学的真理をも含めて、いっさいの知識を疑い、その結果<疑う我>の存在は絶対に疑いえないという自覚から<我は思う、故に我あり・Cogito, ergo sum>を直観的に確実で明晰な第一の真理であるとし、探究の出発点とした。すなわち思考する理性をもとにし演繹的に知識を組み立ててゆく合理主義をその立場とした。しかしその一方で、人間の精神活動に関しては自然の力学的法則に基礎をおいたデカルトの反射の概念では説明できなかった。デカルトは神の存在意義を強調する中で、人間の理性的霊魂という観念論的原理を力学的法則に対するアンチテーゼとしてのもう一つの真理に止揚して二元論を展開した。
 a)簡単な反射:脊髄レベルの反射弓
a)簡単な反射:脊髄レベルの反射弓
反射の最も基本的なタイプは脊髄にみられる。皮膚、粘膜、筋紡錘、腱器官などの受容器からの感受性(求心性)入力が刺激として後根から脊髄内すなわち反射中枢に入り、後角で知覚に関係する神経細胞とシナプスをつくり、介在ニューロンを経て、出力(遠心性)成分の前角の運動ニューロンに刺激を伝える。
このように、知覚性インパルスを運んで脊髄に入る知覚線維と、運動性インパルスを運んで末梢の筋や分泌腺などの効果器に伝える運動線維がある。知覚線維については後根から入り運動性の前角細胞に連絡するものと、そのまま脊髄を上行するものとがあり、前者において脊髄レベルの反射弓が成立する。膝蓋腱を叩いて行う膝蓋腱反射などは、もっとも簡単な反射弓であり、知覚神経線維軸索と運動ニューロンとが直接シナプスを形成してできたと考えられるもので、これを単シナプス性反射とよんでいる。これに対して、両線維の間に数個の神経細胞が介在するものを多シナプス性反射といい、反射弓はより複雑である。元来、脊髄は神経管から発達した中枢神経系の構造物のなかで最も原始的な形態を留めているものであるが、この一見単純な反射弓の構成の中にも伸筋・屈筋に関連する対立する反射弓の間に抑制性の側枝を介して相互に影響しあっているという脳内の神経回路の基本型ともいえる機能の原型がすでに萌芽的に脊髄内にみられることは留意すべきことであろう。
b)複雑な反射:脳幹レベルの反射弓
脳幹の上行路および下行路並びに脳幹網様体を介する一層複雑な反射で、ひとまとめにして、脊髄・脳幹・脊髄
(SBS)
反射ともよばれるものがある。下行神経路として、たとえば、外側前庭脊髄路は伸筋支配運動ニューロンに対しては興奮性に、屈筋支配運動ニューロンに対しては抑制性に働いている。一方、橋網様体脊髄路は伸筋・屈筋支配の脊髄運動細胞に対して逆に働いていることが証明されている。また、感覚入力が直接橋・延髄に入ってくる顔面知覚や聴覚や味覚に関連する反射としては、脳神経レベルで一層発達したものとして、いわゆる単純型と複雑型とが共存して反射弓(ないし系)を形成しているように思われる。このように脳幹レベルにおいては、単純反射系がいくつかのニューロンが介在する回路網を作ることによって一層複雑且つ複合化したものとしてとらえる必要があり、脳幹網様体に関連して意識レベルの問題も含まれてくる。神経回路網の解明は非常に困難であり、集団(マス)または系(システム)として把握することが求められる。すなわち、反射弓を構成する神経細胞の数が増大するにつれて、生体は外部からの入力に対して相互に影響を与えながら新しく発展させた反射系として反応し出力を生みだすようになる。
筋紡錘からの刺激に対して筋肉を一定の長さに保つようにはたらく伸張反射や、侵害受容性刺激に対して防衛的にはたらく屈曲反射などでは、脊髄レベルの反射がメインであるが、脳幹レベルから下行する興奮性および抑制性の刺激がバランスを保ちながら作用している。以上の他に、特定の名称が付けられた多数の反射がある(たとえば、瞳孔対光反射、前庭動眼反射、持続性頸反射など)。また、呼吸・循環を含む自律系にはたらく自律性反射は、単に延髄レベルだけでなく視床下部まで反射中枢の概念がひろがり、姿勢中枢や歩行中枢に関与する反射を論ずる場合には、少なくともその概念は中脳の上部までひろがる。
c) より高次の反射:
ここまでくると、当然の帰結として、脳内の反射中枢の概念は小脳、大脳基底核までひろがってきている。そして、暗黙の内に大脳皮質も組み込まれてくる。動物の随意的運動は、進化の過程で、多種多様な反射運動が成立し、それらが組み合わさり、その結果、連続的に質的にも大きな発展をとげて構築されたものとして理解される。以上のように、反射の概念がひろがり、次第に前脳(終脳と間脳)を含む上位の中枢を包括するにつれて、単に複雑な運動のメカニズムを理解する上での基礎概念としての「反射」ではなく、そこから発展した質の高い、知覚・認知を理解の対象として視野に入れた段階に達するようになる。かくして霊魂という神の住処は存在しなくなるのである。
ここで、上にみてきた事柄を基に、単純な脊髄反射から高次中枢を取り込んだ反射の概念の変遷を縦断的に整理してみる。かつては「感覚器官とその神経による外部入力と出力としての筋運動」のみに注目して、セーチェノフの反射の第二局面(後述)である情報処理機能が考えられた。しかし、今日の知見からすれば、入力は外部環境からの入力だけではなく体内環境の情報もまた重要な入力となっている。さらにこの第二局面(情報処理器官の機能)に重要な影響を与える要因として、内分泌系(ホルモン)、免疫系(サイトカインなど)その他の多くの生理活性物質によって形成される場の条件が挙げられなければならない。このように考えると、反射の第三局面としてセーチェノフが想定した「強化された筋肉的反応」としての情動が「生体の全機能(大脳皮質、皮質下を含めた認識機能や自律系・内分泌系、免疫系を含む)を動員した環境への対応」という反射機能であることが理解できるであろう(後述、4.参照)。
3.チャールス・スコット・シェリントン ( C. S. Sherrington, 1861-1952)の「神経系の統合的作用」という概念の提唱
 シェリントンがLiverpool 大学の生理学教授時代にYale大学に招かれて行った記念講演を基に著わした「The
Integrative Action of the Nervous System(神経系の統合的作用, 1906 年発刊)」という古典書がある。「脳は多数の反射を有機的に統合して複雑な運動を作り上げる作用をもっている」という意味で、反射学を学問として確立し近代神経生理学の基礎を築いた概念として位置づけた点で評価されている。輝かしい成果を生んだこの基本概念も時代に制約された欠点を指摘されることを免れない。実際、多数の反射の合成ないし複合として運動を説明できるかどうか。脳幹よりも上位中枢の脳の機能を対象とする問題に関しては有効なものと考えられるだろうか。シェリントンの偉大な功績の評価は建設的批判を加味してその両面から考察することが必要である。
シェリントンがLiverpool 大学の生理学教授時代にYale大学に招かれて行った記念講演を基に著わした「The
Integrative Action of the Nervous System(神経系の統合的作用, 1906 年発刊)」という古典書がある。「脳は多数の反射を有機的に統合して複雑な運動を作り上げる作用をもっている」という意味で、反射学を学問として確立し近代神経生理学の基礎を築いた概念として位置づけた点で評価されている。輝かしい成果を生んだこの基本概念も時代に制約された欠点を指摘されることを免れない。実際、多数の反射の合成ないし複合として運動を説明できるかどうか。脳幹よりも上位中枢の脳の機能を対象とする問題に関しては有効なものと考えられるだろうか。シェリントンの偉大な功績の評価は建設的批判を加味してその両面から考察することが必要である。
4.イワン・ミハイロヴィッチ・セーチェノフ (И.М.Сеченов,1829~1905)の「脳の反射」という概念
 シェリントンよりも一世代前(32歳年輩)にロシヤで活躍した生理学者セーチェノフは中枢神経系とくに脳の実験生理学的研究に先鞭をつけた。そして脳の活動に反射原理をひろげ、パブロフの高次神経活動の学説を生み出すのに道をひらいた。彼は感覚反射から思考への移行や思考の本性についての問題などの解明に大きな貢献をした。
シェリントンよりも一世代前(32歳年輩)にロシヤで活躍した生理学者セーチェノフは中枢神経系とくに脳の実験生理学的研究に先鞭をつけた。そして脳の活動に反射原理をひろげ、パブロフの高次神経活動の学説を生み出すのに道をひらいた。彼は感覚反射から思考への移行や思考の本性についての問題などの解明に大きな貢献をした。
モスクワ大学医学部に学んで、医学博士取得後、彼は、クロード・ベルナール (1813-1878) , ヨハネス・ミューラー (1801-1858) のもとで研究した。1862年に、セーチェノフは再び、クロード・ベルナールの研究室で、反射運動を抑制する神経中枢の実験的研究を行った。帰国後、これらの実験にもとづいて「脳の反射」(1863)を発表した。
セーチェノフ (I. M. Sechenov) は反射の性質をめぐる議論の中で反射は常に三相の構造をもっていると主張する。第一は外部(あるいは内部)環境が感覚受容器に及ぼす刺激。第二は脊髄あるいは脳への伝達。第三は外部すなわち筋肉へむけての伝達である。今日的表現でいえば、入力、情報処理、出力ということになる。
この構造はカエルなどの実験的研究を通じて下等動物に関してはよく知られており、興奮と制止とが、神経過程の主要な要素として分析されていた。また、環境からの感覚的な刺激作用がない場合には、動物でまったく反射が起こらないということも証明されていた。以上の証拠を基にセーチェノフは、反射を基盤にした脳の機能としての心的活動は、通常複数の感覚刺激があってはじめて生みだされると結論した。
彼は、ヒトも含めて高等動物の脳の活動が反射の特徴のうちの二つをもつことを、すなわち反射のはじまりは感覚器官への刺激作用にあり、反射の終りは筋肉の活動にあるということを論証した。いわば反射の「第一局面」はまず達成された。次の問題として、感覚器官への刺激作用の後で、且つ筋肉の活動の開始前に起こる事象が、反射の「第二局面」、つまり脳内における結合によって説明が可能かどうか。さらに、また、思考や情動が反射によって説明されうるかどうか。セーチェノフは脳のなかに、反射の「第三局面」つまり「筋肉的局面」を強化したり制止したりする機能をもつ一定の中枢の存在を仮想した。彼は情動を、強化された"筋肉的反応"という言葉で説明した(前述、参照)。これは、いわば、個体が環境に対して適応しようとして躍動する全身体的な "mouvement" である。
彼の研究上の指導原理は、世界の物質的統一性、デテルミニスム決定論(註:ビタリスム生気論に対する用語で、生物体においても無生物におけると同様、すべての現象の存在条件は絶対的に決定されていると考える立場)であり、脳の活動に反射原理をひろげ、動物およびヒトの心理活動の反射理論の発端となり、パブロフの高次神経活動の学説を生み出すのに道をひらいた。事実パブロフは、彼におよぼしたセーチェノフの影響について「われわれの研究の出発点は1863年の終わりで、セーチェノフの<脳の反射>が世にでた年である」と指摘している。
5. イワン・ペトローヴィッチ・パブロフ (И.П.Павлов,1849-1936)の条件反射の概念
 無条件反射は、動物の生命を維持していくための基本的な生理現象であって動物の種に応じていろいろな反射弓をもっている。無条件反射活動はいずれも脳脊髄での構造的発達につれて各種の反射弓が形成されることによって機能化するものであり、それぞれの動物の種に共通してみられるものである。そして動物は進化の段階に応じ、ますます外界への適応性を拡げるようになった。つまり動物は、それぞれ生後の経験、つまり「条件づけ」によって、それぞれの個体が獲得した反射活動によって行動するようになったのである。そこでこれらの反射活動を無条件反射に対しパブロフは条件反射と名づけた。
無条件反射は、動物の生命を維持していくための基本的な生理現象であって動物の種に応じていろいろな反射弓をもっている。無条件反射活動はいずれも脳脊髄での構造的発達につれて各種の反射弓が形成されることによって機能化するものであり、それぞれの動物の種に共通してみられるものである。そして動物は進化の段階に応じ、ますます外界への適応性を拡げるようになった。つまり動物は、それぞれ生後の経験、つまり「条件づけ」によって、それぞれの個体が獲得した反射活動によって行動するようになったのである。そこでこれらの反射活動を無条件反射に対しパブロフは条件反射と名づけた。
このように、無条件反射は生物の種に属し、条件反射は個体によって形成される。条件反射は個体の生活のなかで獲得され、反復され強化されるが、用いられないと消去されてしまう。しかし、同じ種類の条件反射が反復して形成されるような環境に生物が置かれた場合には、生物個体と環境との間に持続的結合関係が成立すると考える。このようにして獲得された環境への適応の仕方が 継承され、外見上"遺伝"したかのごとく、その種に定着される。条件反射が無条件反射に転化すると表現される。
ここで生物の種属(同種の個体の集団)と個体(集団を構成する要素)との関係について考えてみたい。個体群である動物の種は、それを構成する個体の無条件反射の結果として、ある個体は環境に適応することに失敗して滅亡し、あるものは成功して生き延びる。こうしたことがきわめて長い時間経過の中で繰り返されたとき、あたかも動物種が条件反射を獲得したかのごとくみえ、その結果として種の環境への適応と放散と繁栄が(時にはその逆が)もたらされる。一方条件反射は個体が獲得するものであるが、高次の神経機能を有する動物種では、ある個体がいつの時にか獲得した環境適応への有利な条件反射が、親から子へ、あるいは群の中で次の世代へと継承されて行き、結果として、あたかもその種に固有の無条件反射のようになっていると考えられないであろうか。誤解を与えないために付言すれば、ここで言いたいことは、パブロフの有名な音による唾液分泌の誘発という条件反射がイヌの群の中で次の世代へ継承されるかどうかと言った議論ではなく、例えば保護された幼鳥あるいは幼獣を人工飼育して自然に戻すときに、その種が継承している"しきたり"に順応できず大変苦労をしている例などを見ると、種の無条件反射とみられるものの中には実は条件反射が継承されている場合もあると考えられるのである。パブロフが提唱した条件反射の今日的重要性は以下に述べるような大脳機能の解明に重要な一石を投じた点で評価さるべきであり、条件反射が"獲得遺伝"によって種に定着するといった考え方は、現在では種の進化とDNA、RNAの進化という最も今日的課題の中でわれわれが検証を迫られている問題ではなかろうか。
条件反射が無条件反射へと転化するという誤解されやすい単純化された概念について、ここでしばらく近代的神経科学の眼から眺めて少し解釈を試みてみようと思う。明らかに条件反射は学習された行動様式を獲得する一つの手段である。それは、脳内の神経回路の可塑性とその refinement に基づいた高次神経活動であり、個体の発達過程のなかで獲得された神経活動がパタン化されたものである。神経活動のパタンとその表現型としての生物の行動様式は、親から子へと一見遺伝されているようにみえるが、世代間に伝わる条件反射ないし「(古典的)条件づけ」の繰り返しとみられる現象である。この際、学習は集団によってなされ、集団生活のなかで維持されている。一般に、種属内集団に特有の高度の行動様式というものは、遺伝子によって規定されている(DNA-dependent)ものと,条件反射獲得の連鎖として世代間に継承されているものとが混在 (mix) して成立していると考えられる。遺伝 (nature) と環境 (nurture) の問題は、互いに矛盾する単なる対立概念ではない。これを機械論的に、静的に、質的に変化をする事のない事象としてとらえていては問題は解決しない。対象は幾世代にもわたるダイナミックな生物学的変化の問題なのであるから、形式論理的にではなく、弁証法的 ( an und fur sich) に考察することが要求される。進化の問題を考えるときには、確率論的に起こった突然変異(mutation)の重複と自然選択の繰り返しが長い世代にわたって経過する間に生じたものが根底にありそれらが継承されてきているということを基本概念としてつかんでおく必要がある。
しかしながら、現代の科学は、いまだ条件反射と遺伝と進化とを結びつけて考察できる段階には達していない。それを承知の上でのコメントであるが、19世紀の三大発見の一つとされるCharles Darwin (1809-1882) の生物進化と自然淘汰(種の起原、1859、刊行)の問題は、今や核酸(DNA、RNA)レベルの遺伝的変化(複製エラー、遺伝子重複、共生進化など)が環境に適応する過程で偶然と淘汰ないし選択への道をゆっくりした速度で(生物の生存安定度と複製すなわち子孫の数を決める生殖能力に依存するが)進行しながら、新しい遺伝子をつくりつつ進化していくという生物学的事象につながっている。現在、脳内の神経回路形成に関わる分子生物学的アプローチの問題を、系統発生学的に認識するという基盤の上に立って、条件反射学を見直すべき時が到来しているのではなかろうか。
さて、一定の段階に達した生物において、神経系統の発生によって刺激を受容する部分と刺激を伝達する部分とが分化する。このような分化によって刺激は局所的なものから伝播される性質のものに変わり、また刺激に対する反応の仕方も分化した。このようにして、筋肉の収縮、腺の分泌というような特殊化が起こり、「興奮性」とよばれる刺激反応性を獲得した。また、生物体が自己を保つためには、身体の各部分からくる刺激に総合的に反応する必要がある。そのためには入力される刺激が一ヵ所に集合されて、まとまった運動を可能にするように身体の各部分にたいして適切な指令が伝達されなければならない。
高等動物の神経中枢は(1)前脳と(2)中脳・後脳との二つの部分に大きく分かれており、機能的にみればこれはおのおの条件反射と無条件反射の器官ともみなしうる。唾液分泌と音の関係でいえば、条件反射の成立後は、イヌは音を聞いただけで、唾液が分泌されるようになる。この条件反射が形成される以前には、その音は聴神経を刺激し、その興奮は大脳へと伝えられるが、聴覚刺激に触発されて大脳から唾液腺へそれを伝える回路はなかった。条件反射が形成されたのちには、この興奮が大脳から唾液腺へと伝えられるようになる。つまり条件反射の形成とはそのような神経の興奮の伝導路が一時的に形成されることを意味する。
大脳が発達して多様な条件反射を形成する可能性が生まれることは、生物体が変化しつつある環境のなかでその変化にすばやく適応して生存しつづける可能性が増大することであって、いうまでもなく生物体の自己保存に有利なことである(上述、ダーウィンの自然選択の記述を参照)。
複雑な条件反射の形成にとって必要なもう一つの条件は刺激の小さな差異を区別することである。実験の結果によればイヌにはかなり高度にこの能力があることがわかる。はじめは刺激が一般的な形で新しい条件反射の成分になるが、あとになると刺激は分化する。たとえば音を条件刺激として条件反射を形成すると、はじめいろいろな異なった振動数をもつ音がみな一様に条件刺激になるが、一定の振動数の音だけを使って条件反射の形成をつづけると、ついにはその特定の振動数をもつ音だけが条件刺激となり、その他の振動数をもつ音は無関心刺激になる。このようになることを感受性が分化する、あるいは、刺激が分化するという。また、大脳半球の後頭葉と側頭葉を切除したイヌに条件刺激を与えても、物体の形とか運動とかを刺激として条件反射を形成することができない。上述のような条件反射の研究を手がかりにして、パブロフは条件反射が形成されるにあたって大脳内でどのような生理的過程が行われているかを追究して、大脳生理学を科学的基礎の上にすえた。
上にみてきたように、条件反射が形成されると一時的に新しい神経路が成立する。そして条件刺激(音など)による興奮は大脳皮質の一定の領域に到達し、この新しい回路を介して下位にある無条件反射の中枢に伝えられ、無条件刺激(食物など)の非存在下でも反射反応が起こる。また、ここで、条件反射形成時の条件刺激となる「感覚」の性質についてあらためて考えてみると、感覚にも階層性(ヒエラルキー)があることが推察できる。たとえば、① 生命維持、種属保存に必須でただちに反応せざるを得ない感覚、②それほど急は要しないけれどもより有利な条件を得るために反応する感覚、③必ずしも反応しなくてもよいが自分のおかれている環境条件を認識している感覚、などである。この感覚の階層性は脊髄・脳幹から大脳皮質に至る下位(または低次)中枢から上位(または高次)中枢にひろがる形態的・機能的発達段階の程度に照応する。かくして動物が高等になるにしたがって条件反射が複雑化し、無条件反射よりも大きな役割をもつようになる。中枢神経系の発達は反射の進化に照応しており、中枢神経系をもつ動物はすべて条件反射を形成する可能性をもっているが、その最高部位である大脳皮質の発達は、条件反射の進化に互いによく対応している。このようにパブロフの条件反射学説は、高次神経活動の反射的性格を明らかにした。
条件反射が研究される以前には、大脳の機能についてはほとんど何も具体的には知ることができなかった。わずかに、動物の大脳の部分的損傷とその機能障害の観察に基づいて、大脳の一定の部分が一定の機能と関係があると推定した(Fritsch and Hitzig, 1870, イヌの大脳皮質運動領の電気刺激で大脳局在論の曙)。条件反射の研究によって、働いている大脳の中で行われている眼にみえない機能をたとえば唾液の分泌というような測定できるものを通して推定することが可能になった。条件反射学説は心理現象を神の摂理による精神活動を介在させること無しに、ある意味では機械論を一層発展させて、弁証法的方法にしたがって研究しかつ理解する道を開いた。
条件反射形成における条件刺激として種々の感覚がある。感覚にも階層性(ヒエラルキー)が存在し(前述、参照)、その発達と条件反射形成のレベルとの関連について言及しておきたい。嗅脳やそれに関連する大脳の領域は、発生学的にみて爬虫類から哺乳類へと進化する過程で、嗅覚に依存して地表面での二次元的生活をする中で発達が促がされたと考えられる。すなわち、環境に適応し個体に有用な形質を獲得しつつ一定の方向に進化する過程で嗅脳およびその関連領域が前脳内に形成される。このように、嗅覚関連領域が脳の前端部に形成されその内部に大脳辺縁系(情動、内分泌、自律系の働きの
"中心"部位)を構成する構造物が原始的な型 (prototype)
として形作られていることは注目に値する。
嗅覚系と同様、感覚の階層性としては低い段階に属する化学物質を受容する味覚系や、これまた低周波振動としての水圧の感覚器官であり、且つ、重力感知や平衡機能などの生存上基本的
(basic)
な感覚(他に味覚機能も兼備している)を司っている側線系があり、それらの感覚刺激は後脳(橋・延髄)に入ってくる。味覚
(contact sense) の入力が嗅覚 (teleceptive sense)
の刺激と同様に直接に扁桃体に投射するのも興味深いが、これらの側線系、味覚系の中枢が動物が進化するにつれて脳幹レベルから(小脳を介した経路と共に)前脳レベルに前方移動して発達してくる。次いでより高いヒエラルキーに属する視覚系や聴覚系の刺激が加わり、それらの中枢が後脳から中脳、さらに間脳、大脳新皮質へと動物の高等化に従って前方に形成されてくる。端的に言って、視覚に依存して三次元空間に適応した動物であるトリなどでは中脳視蓋が大きな役割を担うようになった。樹上生活や直立二足歩行を獲得したサルやヒト(準三次元空間適応動物)においては、さらに種々の要因が加わり、大脳皮質が量的にも質的にも発達をとげた。このようにみてくると、動物一般とくに大脳皮質の発達段階が低い動物も含めて考察したとき、感覚刺激の発達段階に応じて脳幹を含めた脳の対応したレベルで条件反射が形成されると考えられる。動物は種々の感覚刺激に曝された環境に適応した行動様式を条件反射の形成を通じて獲得するのだが、進化論的立場からみて、感覚のヒエラルキーの問題は条件反射の質の問題とも関連する重要な問題である。
7. 高次の符号としての言語の"反射"概念への導入とその確立:第二信号系すなわち言語条件反射
直立二足歩行を獲得した高等霊長類の皮質運動領は、身体を支え歩行を可能にした下肢(脚)の領域 や locomotion の用具であることから解放され道具の作製を可能とした自由上肢(手)の領域が refine されて量質共に大きな発展を遂げる。かくして、道具を用いた労働を協同して行うことができるようになる。
新たな環境におかれて "活性化"したこれらの動物の大脳皮質運動野は、種々の感覚による条件刺激によって "活性化"された感覚野の活動と相俟って動的に変化し、高度の構造をもった皮質に発達する。パブロフはこの事象を、大脳皮質構成に関する「動的モザイク説」と名付け、皮質領内で興奮領域と抑制領域とが静的に定常化しているのではなく、その部位と範囲が瞬間、瞬間ダイナミックな変化をするとして説明している。この段階まで発達した大脳皮質内において、多種多様な感覚刺激の組合せ (cross-matching) による条件反射が形成され、これらの条件刺激を符号として認識するというコミュニケーションの基盤を生みだされ、社会集団のなかで労働するという過程で信号が複雑化してきた。パブロフは「人間が社会的になってきた所にはじめて質的に異なった機能―言語機能系が生じた」と言っている。すなわちコミュニケーションの最高に発展した形態が言語による交流である。なお、後述するように、これらの各段階の底流にあるものは「遺伝子の変化が生みだすその複雑化」である。
最も高度に発達した脳髄をもつヒトにおいては、大脳新皮質の発達に伴い高度な符号の認知が可能になり、伝達事項の複雑化とともに内容が概念化し抽象化した結果として感覚性言語領(ウェルニッケ領野、ここで言語の意味・内容が理解される)が後連合野内に形成される。さらに、環境からの刺激をうけて後連合野内の皮質活動は活発になり、皮質間に発達した神経回路(皮質皮質間結合)を用いて一層脳の発達が促がされ、前頭葉内に運動性言語領野(ブローカ領野、ここで言語の内容が構築され能動的に表現される)が形成されるに至る。このように言語は、動物間の単なるコミュニケーションの手段として成立し発達したものであるが、進化論的にみて個体が環境との接触(相互作用)の中で高度の「条件反射系」を使って思考内容を表現し、抽象化した概念を客観的方法をもって世代間を超えて継承できる手段として発達してきた。これがすなわち、人間社会においてのみにみられる、文化、文明の世代間を超えた継承の基盤である。
ここで、以上の文脈の中で話題にされてきた条件反射と言語の関係について整理してみたい。一般の動物は、一つの無条件反射系と一つの条件反射系の二つしかもっていない。これに反して、ヒトは三つの高次神経系、すなわち、一つの無条件反射系と二つの条件反射系をもっている。一つの無条件反射系とは、皮質下領域にみられる反射あるいは「本能」の体系を言い、二つの条件反射系とは、 大脳皮質にある第一信号系あるいは感覚信号系と第二信号系あるいは言語信号系を言う。第二信号系は、系統発生的にみて、ヒトがサルのような動物段階から社会的集団生活を営むように進化していく過程で獲得した条件反射活動であると考えられる。
このように、動物とくに高等動物の行動は条件反射で構成されている。条件刺激となるものは動物ではすべて具体的な自然の事象であるが、ヒトでは言語が条件刺激となって信号系に加わる。パブロフは言語を第二信号系と名付け、動物とヒトに共通な基礎的な信号系を第一信号系とよんで区別した。言語が条件刺激となって働くとき、それは具体的な自然の事象を表示し、信号の信号として働くのであって、第一信号系とは性質を異にしている。第二信号系は<言語条件反射>とよばれることもあるが、パブロフ学説にしたがえば、条件反射の一種にすぎないのではなく、第一信号系とは質的に異なる高次の信号系なのである。ヒトを含む高等動物では、無条件反射の中枢は大脳皮質下から脊髄にあり、第一信号系の中枢はひろく大脳皮質に形成される。第二信号系は新皮質、とくに言語機能に関係する連合野にその中枢の座があるとされている。次の項でこの問題に立ち入ってみよう。
8.皮質連合野と大脳辺縁系の神経回路と条件反射学説
1) 皮質連合野 (文献、川村、1977、参照)
大脳皮質には皮質連合野と呼ばれる高度な情報処理機能を有する領域が、高等な動物ほど発達している。感覚野からは皮質連合野へ皮質皮質間線維(連合線維)が送られている。大脳皮質連合野には、前(頭)連合野(または前頭前野)と後連合野があり、後連合野はさらに頭頂連合野、後頭連合野および側頭連合野に分けられる。各々の感覚皮質野と皮質連合野との間およびこれら複数の皮質連合野間の結合様式はたいへん複雑である。大脳皮質間の結合の特徴は、第一に、体性感覚野、視覚野、聴覚野など、各々の同じ感覚野内の領域が、比較的短い連合線維によって、強く結合していることである。反面、異なる感覚領野間の結合、あるいは、第一次感覚領野と皮質運動野との間の結合は非常に弱い。第二の特徴として、皮質連合野間の相互結合はかなり強く、高等な動物の脳ほど顕著である。高次機能活動の基盤となる条件反射の形成は、これらの結合、すなわち、神経回路網の上に成立する興奮伝達ないし神経活動の総合的な所産である。
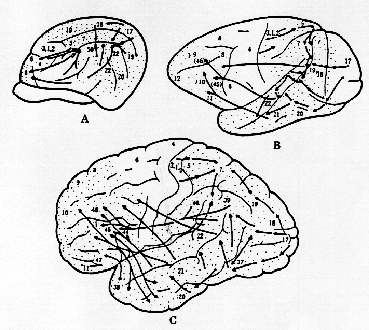
2) 大脳皮質連合野と条件反射第二信号系 (図1)
動物は外界からの感覚刺激を知覚し、認知する。皮質レベルで連合線維間のシナプスが替わるにつれて、一般に神経細胞はより複雑に細工され、処理された情報としての刺激を受けとるようになる。サルにおいて、後連合野内の特定の領域、すなわち、上側頭溝
(superior temporal sulcus) 後部の周囲皮質(posterior STs 域)
に体性感覚、視覚、聴覚などの入力が、一定のパターンをもって集中して終わっている。同様の皮質レベルの感覚刺激伝達の集中性は、ネコにおいても認められ、サルのSTs域とネコの中S上溝
(middle suprasylvian sulcus) 周囲皮質 (MSs域)が対応している。サルの脳にみられるこのSTs域は、視覚性、聴覚性および体性感覚性の性質をもつ各連合野に囲まれており、ヒトの大脳皮質の発達段階にくらべて、たとえ質的に"低次"で萌芽的であるとしても、皮質内の位置的関係や線維結合上の類似性からして、進化論的にみて、ヒトの39野(角回)や40野(縁上回)ー感覚性言語中枢域(Wernicke中枢)ー
に発達し分化する可能性を内蔵している領域と考えてよいように思われる。この段階はパブロフの条件反射理論でいえば第一信号系から第二信号系への移行段階であるといえよう。さらに高次の符号である言語を媒介として抽象的思考の段階に進むためには、より発達した前(頭)連合野(前頭前野)への入力が必要となる。この過程のなかで前頭前野内に能動的性質をもった言語に関する領野(Broca
中枢)が新しく形成される。この段階になって初めて概念を形成し判断を下し、推理を試みるという思考能力を備えた前頭葉皮質が形成され発達してくる。こうして、言語が特殊な信号系として追加されることによって、新しい神経活動の原理、すなわち抽象と普遍化が行われるようになる。すなわち、質的に発展を遂げた高次の言語条件反射といわれる第二信号系が形成されるのである。
図1.
3) 連合野と扁桃体・海馬を中心とする辺縁系との関係
(図2,図3)
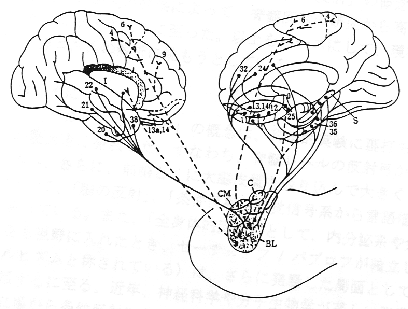
側頭連合野や前頭連合野の腹側部は扁桃体および海馬周辺皮質とかなり強い相互的な線維結合が認められている。扁桃体は視床下部、視床背内側(MD)核、側坐核などにも線維を送っており、視床下部やMD核を介して、前頭葉皮質と密接な関係をもっている。海馬
は皮質連合野との間には直接的結合はなく、嗅内野や海馬台といわれる領野を介して、間接的に結合している。さらに、この嗅内野や海馬台は脳弓を介して乳頭体、さらに視床前核、帯状回と順々に連結し、いわゆる情動の回路
(Papez,1937) 内の重要な位置を占めている。
また、海馬内に空間や場所を認知する "記憶"ニューロンが存在すること、および扁桃体内に外的刺激の生物学的意味(報酬性か嫌悪性か)を判断する
"認知・記憶 "ニューロンが存在することが明らかにされている。このように海馬は、環境への定位に関する新しい情報を、新皮質に蓄えてある保存された情報と照合するという一種の連想作用ともいうべき重要な役割をもっている。これが行動と結びつくとき、個体の探索活動として現れてくる。パブロフ
によれば、探索活動とは定位反射(「これはなんだ反射」の別名がある)を基礎にして形成された複雑な反射活動である。一方、扁桃体は、主として側頭連合野と前頭前野眼窩面皮質を含むかなり広範囲にわたる皮質連合野と相互に連絡しており、相互の情報交換により、外界刺激と情動とを連動させて意味づけ(価値判断や評価)をすることを可能にしている。海馬からくる空間的ないし環境内の出来事の連合記憶の情報は、扁桃体にもインプットされ、さらに、視床下部を中心とした情動性神経回路網の内に取り込まれる。このように扁桃体は、快、不快の情動の発現を惹き起こす要(かなめ)となっている。ここでわれわれは、パブロフの学問上の師であるセーチェノフが著書「脳の反射」のなかで意欲や情動という当時としては科学の対象外と思われていた心理機制を念頭において、これを制御された
"筋肉的反応"という環境に対応する概念として捉えていたことに驚きの眼をもって想起するのである。
図2.
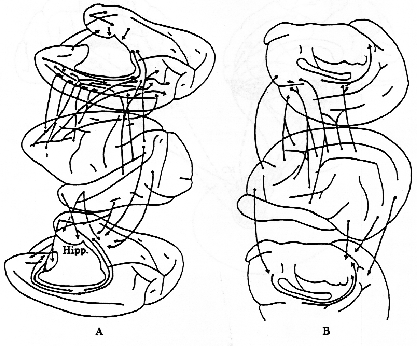
図3
脊椎動物は脳の発達段階からみて以下の5-6のカテゴリーに分けられよう。すなわち、1.主たる発達が中脳以下の脳幹に留まっている動物 2.前脳(間脳と終脳)が発達した動物、3.新皮質が発達し領域化が進んでいる動物、4.連合野がかなり発達している動物、5a.感覚性言語野の発達段階に達している動物、5b.運動性言語野も発達し完成している動物、である。4-5b
を一括して 4.皮質連合野が顕著に発達している動物
とすると、4つに大別される。
ところで、大脳皮質の発達がほとんどみられない爬虫類以下の動物の行動様式の基礎となるものは基本的に無条件反射である。これらの動物では中脳視蓋近傍の領域が条件反射の中枢としての役割を担っていると思われる。発生学的にみると、中脳・後脳境界部で形態形成初期の過程で起こる重要な遺伝子の発現のカスケード
(Pax→Wnt→Enなど)
がみられることは注目に値する。すなわち、この領域は脊椎動物の発達段階で捉えられる重要な領域で、中枢神経系のいわゆる
"第一次"または"低次のオーガナイザー領域 "であるとみなされている。鳥類における特異的な線条体の発達は別個に論ずべき問題として脇におくとして、哺乳類霊長目(ヒトを除く)にいたる段階まで動物の脳は発達を続け、皮質連合野が次第に皮質内で広い領域を占めるようになるが、言語野といわれる領域が形成されるまでには至っていない。条件反射第一信号系の基盤までの形成にとどまっている。さらに発達してヒトの段階になると、前脳形成機構の内で、新皮質ニューロンとくに連合野の形成に、さらにとくに、言語野の形成に関与する遺伝子の発現とその進化が第二信号系の発達に深く関与しているはずである。上述の発達段階の分類に照らして言えば、サルは3.-4.の段階にあり、ヒトは5.の段階に達している。
ところで、前脳形成に関与する転写調節因子としては、脳の大脳化にともない終脳の神経上皮に部位特異的な発現パタンを示す遺伝子群として、マウスにおいて現在Dlx-1,
Dlx-2, Nkx-2.1, Nkx-2.2, Otx-1, Otx-2, Emx , Gbx-2, BF-1, BF-2,などが知られている。これらのほとんどの遺伝子はハエの段階にすでに存在している遺伝子
(Distal-less, Dll ; NK-2 ; orthodenticle, otd ; empty spiracles, ems ; unplugged など )
のhomologues(相同遺伝子)であり、遺伝子の構造としては多様化している。マウスは3.の発達段階に位置する動物である。従って、これらの遺伝子の解析をすることにより、新皮質形成の機構を調べることができる。われわれは現在、サルやましてヒトのこれらの相同遺伝子を手中にしていないが、logical
に言って、これらは齧歯類のものよりも遺伝子の進化(重複、変異など)が進み
同一gene family
内に亜型がより多く存在し、いわゆる多様化がみられる筈である。この遺伝子の多様化は進化の過程で突然変異の繰り返しによる遺伝子の重複と自然選択により複雑化したもので、同じように前脳の形態形成に関与していたとしても、新皮質形成とか、連合野形成とか、言語野形成とか質的に発展した働きを具有しているものであろう。このように、前脳形成に関わる転写調節遺伝子を発生学的に系統的に調べることにより、サル、ひいてはヒト大脳新皮質の領域化や連合野に関連した遺伝子、さらには、ヒトをヒトたらしめる言語中枢を規定している遺伝子の探索の方向に次の世紀には突入することができよう。
参考文献
Creutzfeldt, O.D. (1983): Cortex Cerebri, Leistung, strukturelle und funktionelle
Organisation der Hirnrinde, Springer-Verlag.
Engels, F.(1896) : Neue Zeit, 猿が人間になるについての労働の役割.
大月書店編集部編.
林 髞 (1944): 大脳生理学 三笠書房
平澤 興 (1950): 大脳の最高中枢 日本医書出版
鹿島晴雄 (1994) : Pavlov, I. P.
心理活動の客観的研究、続・精神医学を築いた人びと 上巻、133頁-145頁(松下編)、ワールドプランニング
Ltd.
Kawamura, K. (1973a,b,c) : Corticocortical fiber connections of the cat cerebrum. I. The
temporal region, II. The parietal region, III. The occipital region, Brain Res.,51:
1-21, 23-40, 41-60.
川村光毅 (1977): "連合野"の線維結合 (I) 皮質間結合
ーサルとネコの皮質間結合の比較と"連合野"の発達についての試論ー
. 神経進歩 21: 1085-1101.
川村光毅 (1982): 角回を中心とする皮質間連絡. 失語症研究 2: 208-214.
川村光毅 (1985): 前頭葉の解剖学 --前頭前野の皮質間結合を中心に--.
精神医学 27: 611-617.
川村光毅 (1988): 連合野の統合. 新生理学体系(監修:勝木ら)第12巻:高 次脳機能の生理学(鈴木、酒田
編). 5-12. 医学書院.
川村光毅 (1993) : 認知機能についての機能解剖学的考察.
生物学的精神医学(小島、大熊編)第4巻 認知機能からみた精神分裂病 183-198.
学会出版センター.
Nieuwenhuys,R., Voogd,J & van Huijzen, C. (1988): The Human Central Nervous System. A
Synopsis and Atlas. 3rd Ed.,Springer, Berlin-Heidelberg.
Pavlov, I.P. (1927) : 条件反射による動物の高次神経活動(行動)の客観的研究の20年
岡田靖雄、横山恒子 訳、1979 岩崎学術出版社.
Pavlov, I.P. (1927) : Lectures on the Activity of the Cerebral Hemisphere, Leningrad.
林 髞 訳、1937. 条件反射学
―大脳両半球の働きに就いての講義― 三省堂; 川村 浩 訳、1975.
大脳半球の働きについて― 条件反射学― 岩波書店.
Pavlov, I. P. (1927) : Conditioned Reflexes, An Investigation of the Physiological
Activity of the Cerebral Cortex. (Transl. & Ed. by G.V. Anrep) Oxford Univ. Press.
パヴロフ選集 (1962)
上巻、下巻 (コシトヤンツ編、東大ソ医研訳)
合同出版社
Pavlov, I. P. (1941) : Lectures on Conditioned Reflexes, Vol.1 (The Higher Nervous
Activity (Behaviour) of Animals) & Vol. 2 (Conditioned Reflexes and Psychiatry) ,
(Transl. & Ed. by W.H.Grantt) , Lawrence & Wishart Ltd., London
Sechenov, I. M. (1952-1956) : Selected Physiological and Psychological Works, prepared for
print by the Academy of Sciences of the U.S.S.R., Moscow.
Sherrington, C. S. (1906) : The Integrative Action of the Nervous System, London:
Constable & Co. Ltd. New Haven: Yale Univversity Press
Wells, H. K. (1956-1960): Pavlov and Freud, International Publishers Co., Inc.USA,
中田実、堀内敏 訳、1966. パブロフとフロイト. 黎明書房.
図の説明
図1.
ネコ(A)、サル(B)、ヒト(C)の大脳皮質内において見られるであろう種々の感覚性刺激の伝達経路を示す模式図。アラビア数字はBrodmannによる皮質区分番号を表す。皮質連合野が点描されている。(川村、1982より)
図2.
サル(A)とネコ(B)における新しい皮質から古い皮質への連合線維の投射の比較。(川村、1977より)
図3.
扁桃体と大脳皮質の相互線維結合。BL、扁桃体基底外側核; C、扁桃体中心核; CM、扁桃体皮質内側核; I、島; S、海馬台。数字はBrodmannの大脳皮質領野を表す。(Nieuwenhuysら、1988より改変)
本節を書いた意図を明らかにしたいため、余白を利用して一言したい。パブロフの条件反射理論を再評価するキッカケをつくりたかったことが、正直なおもな動機である。ネルヴィズムの立場から認知機構については解明がかなり進むであろうが、精神機能のもうひとつの大きなジャンルである情動機構については、異なる側面から、あくまでも物質に第一義的な基礎をおいて、論を進める必要があろう。紙数の制約もあり、ここではこの視点からの考察が十分でない。情動(emotion/passion)の生物学的意義を含めた論旨についてはほかの拙論(情動の機構と歴史的考察。脳の科学、20、709-716,1998)を併読いただけると幸いである。また、最近の興味深い単行本の参考文献としては、以下のものがある。
1) Jean-Didier Vincent(1994): Biologie des Passions, (ed. Odile Jacob)
2) Antonio R. Damasio (1996): Descartes' Error-Emotion, Reason and the Human Brain, (Papermac)
3) Daniel Goleman (1996): Emotional Intelligence (Bloomsbury)
4) Terrence Deacon (1997): The Symbolic Species -The Co-evolution of Language and the Human Brain (Penguin Books)
5) Josef LeDoux (1998): The Emotional Brain -The Mysterious Underpinnings of Emotional Life (Weidenfeld & Nicolson)
脳と神経-分子神経生物科学入門(金子章道、川村光毅、植村慶一編、共立出版株式会社、1999発行)
284頁-296頁より許可を得て転載