認知機構についての機能解剖学的考察―正常と異常(Mechanism of recognition, anatomical aspects) |
川村光毅

| はじめに | |
| 1)認知機能の要素について | |
| 2)大脳皮質間の結合について | |
| 3)大脳皮質連合野関連の諸問題 | |
| 4)皮質連合野と扁桃体・海馬を中心とする辺縁系との関係 | |
| 5)抽象的認識の獲得と前脳機能の乱れ | |
| 6)おわりに |
外界からの刺激、すなわち、光、音、熱のような物理的エネルギーまたは物質の選択的な結合によって感覚受容細胞の膜に変化が起こり、それが電気現象としての興奮を惹き起こし、末梢神経をへて脊髄と脳に伝達される。中枢神経内の電気的興奮は、神経伝導路と称するシナプス回路を上位中枢に向かって進み、前脳部で感覚・知覚の解析がなされる。その結果、一部がいわゆる記憶として保持され、繰り返し解析結果が総合的に判断される。そして、その解析結果が環境に対して運動系および自律系の反応を通じて表現される。前脳部の構造としては、大脳新皮質、海馬、扁桃体、基底前脳部(または前頭葉基底部)、基底核、視床、視床下部などが含まれる。もっとも、生物学的に全体的に認知と行動の問題をみてみると、最も下等な単細胞動物では、これらのプロセスはすべて一つの細胞内で行われているが、高等動物になるにつれて、それぞれに分化した器官が役割を分担するようになり、次第にその機能が分化してくる。動物は外界の刺激を知覚し、認知して、あるいは、体内環境を受容し、かつコントロールして、自律系反射を含む活動を基本とした反応を行うが、神経系が発達し脳と脊髄(中枢神経系)が高等化するにつれて、その反応形態は一層複雑巧緻となる。
1)認知機能の要素について
感覚器官を通して、視覚、聴覚、身体感覚、嗅覚、味覚の刺激が脳の内に情報源として入ってくる。嗅覚と視覚の刺激は前脳から、聴覚と味覚の刺激は後脳から、身体知覚は後脳と髄脳から中枢神経系内に入り、そこの感覚性ニューロンを興奮させることから、認知機能の最初のステップが始まる。興奮活動の主たる流れは、脳内を上行して前脳レベルの上位中枢にまで達するものと、後脳レベルの下位中枢に向かうものとに分かれる。動物性機能に関わるものとして、前者は大脳の認知機能に後者は小脳の運動機能に関連しているものと考えられる。この他、植物性機能に影響を与える刺激は脳の広い範囲に拡散する。ここで問題とするのは動物性機能のうち大脳にまで上行する認知ないし認識の系に関わるものであるが、第一次の大脳感覚野への上行性伝導路についての系統的な記述は一般の教科書にゆずることにする。さて、嗅覚系を含めてすべての感覚系の刺激・興奮は視床核を経由して大脳に至り、そこでそれぞれ特定の皮質感覚野に伝えられる。この伝達様式であるが、個別の感覚は、一般に模式的に考えられているように直列的に第一次、ついで第二次の皮質領野へとすべての興奮が伝わるのではなく、皮質への入力は並列的になっている。すなわち、それぞれの皮質感覚野は視床皮質繊維が終止する異なる複数の領域をもっている。それぞれの感覚様態を伝える脳幹からの入力繊維が終止する感覚性視床特殊核は各々が独立しており、しかも、特定の感覚野以外には繊維を送らない。大脳皮質のうち、感覚野および運動野としてその機能が末梢刺激の知覚あるいは効果器への作用と対応する領域はむしろ小さく、大脳皮質には非常に大きな皮質連合野と呼ばれる高度な情報処理機能を有する領域が、高等な動物ほど発達しており、その最たるものがヒトにおいてみられる。上記の皮質感覚野からは皮質連合野へ皮質皮質間繊維(連合繊維)が送られている。感覚と知覚とを、どちらも刺激が感覚器官へ直接に働きかけて発生し、その働きかけが終わると停止するものという意味で区別せずに、一括して「感覚」と呼ぶ場合もある。しかし、この両者を区別するとすれば、外界の事物または現象の個別的な要素的な性質(感性的特性)が反映された映像が「感覚」であるのに対して、これらの感覚を基礎にしながら、体験される諸感覚が孤立したものとしてでなく、統合的に一つの像として、つまり総合的に意味のあるものとして反映されたものが「知覚」である。さらに、知覚と認知の関係について言えば、現時点で知覚している対象の知覚像を、記憶している一般的表象と照合して「是これ也」と判断することを「認知」すると言う。用語の問題に深入りしたが、これらの事象が大脳皮質の感覚野および連合野で起こるのである。ところで、大脳皮質連合野には、前(頭)連合野(または前頭前野)と後連合野があり、後連合野はさらに頭頂連合野、後頭連合野および側頭連合野に分けられる。各々の感覚皮質野と皮質連合野との間およびこれら複数の皮質連合野間の結合様式はたいへん複雑である。いわゆる大脳の高次機能といわれるものは、これらの結合、すなわち、神経回路網の上に成立する興奮伝達ないし神経活動の総合的な所産である。要素的感覚の認知から符号・図形・空間・音質・化学物質などの認知ないし認識や意味付けの段階をへて、複雑な符号認識の最たるものである言語認識あるいは抽象的(論理的)認識の問題を総合的に研究する者にとって、現在の解剖学、生理学、生化学などを背景とした分析的戦略のみでは克服できそうにない壁が立ちふさがっているように見える。分析的定量的な解析を基盤としつつ、さらに進んで、言語認識の領域に踏み込むためには、脳を構成する物質の機能の質的変化を発展(飛躍)的に捉えるという観点から考察しなければならない。記号ないし符号の任意性がまだ萌芽的なサル以下の動物の身振り記号と、記号の任意性を獲得したヒトの音声記号(これによって初めて抽象的概念が表される)とは異なっており、進化的には神経細胞の量的増大として進んできたのには違いないが、その量的変化がヒトという動物種固有の機能として、質的に異なったものをうみだした。ヒトの認知を研究の対象とするとき、生物進化のプロセスの中で起こったこの重要な質的変化に注意をはらわなければならない。われわれは、言語活動の上に成立する抽象的認識にいたるまでの脳内プロセスに迫る研究を正しく進めるために、物質のダイナミズムを基盤とする哲学を必要としている。そこには、神やオカルト的要素が入り込む余地はない。
大脳皮質間の結合を解剖学的にみたとき、その特徴としていえることは、第一に、
体性感覚野、視覚野、聴覚野など、各々の同一皮質感覚野内の領域が、比較的短い連合繊維によって、互いに強く結合されていることである。その反面、異なる第一次の皮質感覚領野間の直接的な結合、あるいは、第一次感覚領野と皮質運動野との間の結合は皆無に近いか、存在したとしても非常に弱い。しかしながら、皮質皮質間結合の第二の特徴として、一般により高次の領域と考えられている皮質連合野間の相互結合はかなり強く存在しており、この結びつきは高等な動物の脳ほど顕著である。このことは、最近、行動生理学的研究によって明らかにされた以下の事実と一致する。すなわち、イヌ、ネコの類では同一知覚間照合(intramodal
matching)が可能であるが、異種知覚間の照合(intermodal matching or
cross-modal transfer)は、サルの段階になって初めて可能となる。この場合には、個々の知覚から独立したイメージ(image)が成立していると推察されるので、次の高い段階である言語的符号ないしシンボルと照合されるまでの途は、すでに踏み出していることになる。次いで、興味深いことは、大脳辺縁系に属する海馬周辺のいわゆる古い皮質領域と、最も新しいこの皮質連合領域とが相互に強く結合しており、この結合関係も、動物が高等になるほど、領域的にも、量的にも大きく強くなっていることである(図1)(6-14)。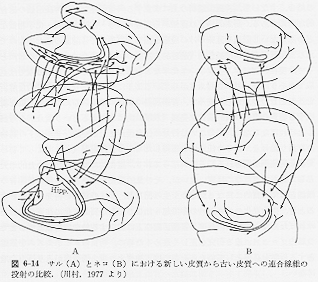
以上の事実は、筆者が、ネコ、サルにみられる連合繊維の結合様式を調べてみた結論であり、1930,40年代に発表されたチンパンジーの皮質間結合の知見をも勘案して、元来実験的に調べることのできないヒトの脳の場合にも当てはまるというのが、筆者の見解である。
見る、聞く、触れる、などの感覚が、感覚器官で受入れられたのち、興奮がニューロンを介して、上位ないし高位の中枢に運ばれて知覚され、そして認知される。このプロセスでは興奮域の周囲が皮質視床間のいわゆる反響回路により抑制的な機能領域となり、皮質感覚野で「浮き彫り現象」的に処理されるという特徴がある。Pavlov流に言えば、大脳皮質の一群の神経細胞に興奮が起こり、他の神経細胞に外抑制の法則によって、誘導のため抑制が起こる。このために、特定の客体が選びだされたり集団化されたりする。次いで、より高次に位する感覚性連合野(後連合野)で、古い皮質、たとえば、海馬などと照合されて、統合的に認知される。ヒトの場合に重要なことは、このほかに、抽象概念を理解できる基盤である言語認識という機構が加わっていることである。次項では、この抽象概念の形成を司る脳領域について考察する。
① 後連合野で起こっていること;
後連合野において、外界からの感覚刺激を知覚し、認知する。皮質レベルで連合繊維間のシナプスが替わるにつれて、一般に神経細胞はより複雑に加工され、処理された情報としての刺激を受けとるようになる。視覚系に例をとれば、点から線、形、容貌といったように。感覚器官、脳幹、間脳、第一次感覚野、後連合野内という一連の刺激伝達系と称される神経路内では、段階的(stepwise)に受けとる信号パターンが異なっている。サルにおいては、この連合野内にある特定の領域、すなわち、上側頭溝後部の周囲皮質(posterior
STs 域)
に体性感覚、視覚、聴覚などの入力が、一定のパターンをもって集中して終わっている。同様の皮質レベルの感覚刺激伝達の集中性は、ネコにおいても認められ、サルのSTs域とネコの中S上溝周囲皮質
(MSs域)が対応している。サルの脳にみられるこの STs域は、視覚性、聴覚性および体性感覚性の性質をもつ各連合野に囲まれており、ヒトの大脳皮質の発達段階にくらべて、たとえ質的に"低次"で萌芽的
(rudimentary)
であるとしても、皮質内の位置的関係(やおそらく繊維結合上)の類似性からして、進化論的にみて、ヒトの39野(角回)や40野(縁上回)ー感覚性言語中枢域(Wernicke中枢)ー
に発達し分化する可能性を内蔵している領域と考えてよいように思われる(図2)(6-15)。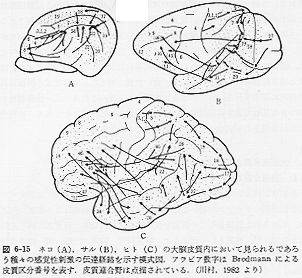
系統発生学的ないし進化論的にみて、この段階の完成が進むなかで、感覚的知覚・認知から簡単な弁別学習の経過をへて符号認知への基盤が形成されるものと考えられる。さらに高次の符号(言語)を媒介として抽象的思考の段階に進むためには、より発達した強力な前(頭)連合野(前頭前野)への入力(形態学的表現でいえば刺激の伝達経路)の存在が必要条件となる。この条件がみたされた時に不断の社会的接触と労働を通じて脳が全体的に発展していく(運動野、感覚野、後連合野の活動で皮質ニューロンは相互に影響を与えつつ機能的に質的発展をとげる)。この過程のなかで前頭前野内に能動的性質をもった言語に関する領野が新しく形成されてくると考えられる。この段階になって初めて概念を形成し判断を下し、推理を試みるという思考能力を備えた前頭葉皮質が形成され発達してくる。このようにみてくると、「言語」の使用と直立歩行の結果として自由となった手を用いる労働、および、その後の生産の重要な鍵となった「道具」の使用がサルからヒトへの進化の根源となったことが理解される。なお、 Pavlov(1927)は感覚、知覚認知と言語活動の関係について考察し、ヒトと動物に共通のものである感覚を「第一信号系」と呼び、これに対置して、ヒトにのみ特有のものである言葉を、感覚の信号(ないし符号)という意味で「第二信号系」と名づけた。ヒトの主観的世界(心理や意識や創造性)の中で、客観的実在を思いうかべる記号としての言葉の認知と理解(感覚的構語機能、いわゆる Wernicke 言語中枢において)並びに主観的内的活動を何らかの物理的媒体を通して表出し他の人に伝達しうる言語の構築(積極的構語機能、いわゆる Broca 言語中枢において)も、両者ともに、物質に基盤を置いた自然科学の対象となりうる。こうして、言語が特殊な信号系として追加されることによって、新しい神経活動の原理、すなわち抽象と普遍化が行われるようになる。
② 前頭連合野で起こっていること;
次に、形態学的に前頭連合野への入力についてみてみると、①視床皮質投射(背内側核MD核より前頭前野への投射)と、②後連合野から前頭前野への皮質・皮質間投射の増加が考えられる。ここで、サルとネコとを較べてみると、ネコの後連合野からの前連合野に向かう連合繊維は、量的にはわずかであるが、サルの段階になるとかなり多くなり、その上、集中性の傾向が認められる。しかも、サルのSTs域内の起始ニューロンの部位と前頭前野内の終止部位との間の関係をみると、この皮質間の連合繊維投射系には、ある程度の部位局在性が明らかに存在する(図3)。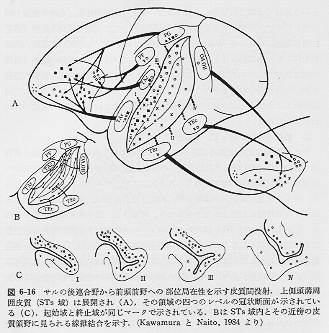 たとえば、STs域の尾側半からは前頭眼野(8野)に投射があり、一方、STs域の吻側半からの投射をみるとその背側壁からは、前頭前野の主溝の背側皮質部分に、その腹側壁からは主溝の腹側皮質部分に、それぞれ、多くの連合繊維を送るという傾向が認められる。また、興味のひかれる点は、
Walker (1940)が細胞構築学上45野と名づけた、弓状溝下行枝の前方域へ投射する起始ニューロンの分布が、STs域内でBruceら(1981)が見いだした
STP域 (superior temporal polysensory area,
上側頭溝周囲皮質内にみられる多種感覚性反応ニューロンが多く存在する領域)の範囲によく一致していることである。一方、逆方向である前連合野から
STs域近傍への投射は発達が悪く、弓状溝前方域や主溝周囲の比較的限られた領野から連合繊維を受けているにすぎない。また、嗅内野など、ヒトでいう海馬旁回の付近からの後連合野への投射も、ある程度の量が存在しており、さらに、鉤状束を形成する前連合野の腹側部分と、側頭連合野の前方域(側頭極)との間の相互結合も、サルの段階ではかなり強く認められる。
たとえば、STs域の尾側半からは前頭眼野(8野)に投射があり、一方、STs域の吻側半からの投射をみるとその背側壁からは、前頭前野の主溝の背側皮質部分に、その腹側壁からは主溝の腹側皮質部分に、それぞれ、多くの連合繊維を送るという傾向が認められる。また、興味のひかれる点は、
Walker (1940)が細胞構築学上45野と名づけた、弓状溝下行枝の前方域へ投射する起始ニューロンの分布が、STs域内でBruceら(1981)が見いだした
STP域 (superior temporal polysensory area,
上側頭溝周囲皮質内にみられる多種感覚性反応ニューロンが多く存在する領域)の範囲によく一致していることである。一方、逆方向である前連合野から
STs域近傍への投射は発達が悪く、弓状溝前方域や主溝周囲の比較的限られた領野から連合繊維を受けているにすぎない。また、嗅内野など、ヒトでいう海馬旁回の付近からの後連合野への投射も、ある程度の量が存在しており、さらに、鉤状束を形成する前連合野の腹側部分と、側頭連合野の前方域(側頭極)との間の相互結合も、サルの段階ではかなり強く認められる。
次に、サルの前頭連合野の弓状溝の前方域について、その領域が視覚性および聴覚性の選択的注意に関わっていることに触れておきたい。すなわち、この前頭葉内の領域には、物体を注視した時に反応するニューロンが部位局在的に存在しており、また、音源を選択的に注意して聴いた時にとくに反応するニューロンも視覚性注視反応ニューロン存在領域の周囲に位置して認められている(弘前大学・鈴木のグループにより,1986)。これらのニューロンが同定された領域は、物体の位置や視空間の認知機能に関連している頭頂連合野
(5,7
野)からの連合繊維を受けており、視覚性および聴覚性、さらには体性感覚性の認知(ないし認識)を考察する上で今後重要な機能解明へのターゲットとなってくると思われる。
以上、皮質連合野の発達と機能およびその関連について考察したが、前頭前野から後連合野にいたるいわばフィードバックともいうべき逆方向の皮質皮質間投射は、前頭前野への投射に比べて、量的に非常に少ないことをとくに注目したい。なお、連合野で高度に処理された情報のアウトプットについて、その一部を次項で考察する。
大脳皮質連合野、とくに、側頭連合野と前頭連合野の腹側部は扁桃体および海馬周辺皮質とかなり強い相互的な繊維結合が認められている(図4)。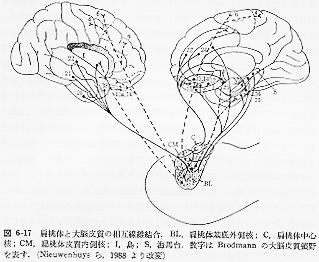
扁桃体とくにその内側・皮質核群はその他、視床下部、視床背内側(MD)核、側坐核、手網核にも繊維を送っており、視床下部やMD核を介して、前頭葉の主に眼窩面皮質とも密接な関係をもっている。
いわゆる固有の海馬 (hippocampus proper)と呼ばれるCA1~CA4の領域は最も新しい皮質である皮質連合野とは直接的には結合しておらず、嗅内野や海馬台といわれる領野(ヒトの海馬旁回(28野)に相当する所)を中継領域として、間接的に結びついている。さらに、この嗅内野は扁桃体と結びつくとともに、海馬台、乳頭体(脳弓を介して)、視床前核、帯状回と順々に連結し、いわゆる情動の回路(Papez,1937)内の重要な位置を占めている。
ここで海馬も扁桃体も共に記憶(memory)に関与していることを一言述べておきたい。これらの構造物が記憶の情報処理の中で果たす役割を理解するために、これら領域内のニューロンの性質をサルを用いて神経生理学的に分析することが最近可能となった(富山医科薬科大学・小野のグループにより,1992)。その結果海馬の領域内に空間や場所を認知する記憶ニューロンが存在すること、および扁桃体内に外的刺激の生物学的意味(報酬性か嫌悪性か)を判断する認知・記憶ニューロンが存在することが明らかにされた。つまり、海馬には、ある特定の場所に動物がおかれたときに反応するニューロンが存在する。筆者自身の経験でも、新しい職場に通勤することになったとき、道順や乗物を待つ場所や曲り角にある標識を認めたりする行動のパターンが決まり安定するまでの期間、かなり"疲れる"ものである。自己と環境との連続的に変化する位置関係を認知し、なかば無意識的な状況においても大きな間違いをすることなく動作するようになるまでのこの過程では、おそらく種々の部位の海馬ニューロンが海馬旁回を介して新皮質と照合されて活動しつづけると思われる。この段階がすぎると、新皮質優位の知的探索がスムースにできるようになる。
このように海馬は、環境への定位に関する'新しい'情報を、新皮質に蓄えてある'古い'保存情報と照合させながら、区別して記憶すべきものとして定めるという(一種の連想作用ともいうべき)重要な役割をもっている。これが行動と結びつくとき、探索(的)活動として現れてくる。Pavlov によれば、探索(または探求)活動とは定位(または詮索、指南)反射(「これはなんだ反射」の別名がある)を基礎にして形成された複雑な反射活動である。この定位反射(無条件反射と条件反射の二種類がある)に関与するニューロン集団の内には、海馬や前頭前野にみられる「選択的注意をしたときに新しい事象、物体、場所などに対応して反応する」ニューロン群も含まれると言えよう。
一方、扁桃体は、主として側頭連合野と前頭前野眼窩面皮質を含むかなり広範囲にわたる皮質連合野と相互に連絡しており、互いに情報を交換することによって、直面する外界刺激と情動とを連動させて意味づけをすることを可能にしている。つまり、'どろどろした'生活に根源的な事象、出来事、物体などの価値判断や評価を行っている。海馬からくる空間的出来事の連合記憶の情報は、扁桃体にもインプットされ、さらに、視床下部を中心とした情動性神経回路網の内に取り込まれる。このように扁桃体は、快、不快の情動の発現を惹き起こす要(かなめ)となっている。なお一言すれば、個人や家族や社会に関わるいわば'高尚な'人生の価値意識とか価値体系については、前頭連合野が関与しているが、繊維結合の上からみて、両者は視床下部で連結されているといえよう。
情動の脳内機構は複雑で不明な点が多い。ただここに、視床下部を中心とした領域が情動行動に深く関わっていることを示す研究がなされているので言及しておく。すなわち、情動には以下の二つの系が考えられている。
1 報酬系快接近行動positive
2 罰(痛みなどの嫌悪)系不快逃避/攻撃行動negative
報酬系は中脳被蓋の腹外側部から視床下部の外側部を上行する内側前脳束
(medial forebrain bundle )
に一致した領域で、この束の一部をノルエピネフリンや
ドパミンなどの
カテコールアミン上行経路が形成していることは興味深い。一方、罰系の方は中脳の背内側部から視床、視床下部、前脳基底部の内側部に連なる背側縦束
(Sch¨utz bundle)
の経路にほぼ一致した領域で、室周系を中心としている。
これらの領域と扁桃体が神経繊維によって結びついている。扁桃体と海馬との間の繊維連絡がサルで存在することが明らかにされており、価値、評価の判断をする役割が扁桃体で、それを支えている短期記憶系がおそらく海馬であろう。これらの出力が視床下部に入り(図5)、情動の表出が形成される。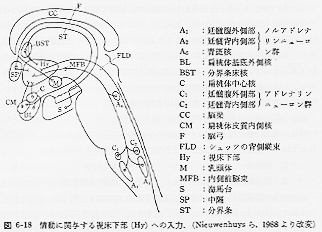
さらに一部が乳頭体・視床前核・帯状回・前頭前野・補足運動野から構成される神経回路をつうじて、意識化された意欲の形成ステップにもつながっている。この意味において、意志は、ある意味では、意識の機能の一つである感情の一種であるとも考えられ、知・情・意の連係はこのようにして脳内に形成されていく。 連合野から大脳辺縁系(皮質および皮質下)への投射が動物が高等化するにつれて増加することは、生存の基盤となる神経機構である大脳辺縁系に対して、この投射系が環境から得た最も組織化された情報を供給するメカニズムであることを考えると非常に興味深いものがある(図1)(6-14)。
このようにみてくると、われわれの認知機能というのは、感覚の要素的なものから、高度に意識化された抽象的認識の段階までの広いスペクトルをもっており、これらを、統括的にみてみると、言語機能(感覚性および運動性)領域を含めた広範な皮質連合野および扁桃体と海馬を含む大脳辺縁系とが相互的に結合することによって、現在の環境内の情報が評価され判断され、情動ないし感情に結びつき自己中心的空間(egocentric space)のなかで認識されるという仕組みが判かるような気がする。精神分裂病の認知機能の異常というものは明らかに感覚要素的なものではない。少くとも、これを中心とするものではない。Minkowski (1953) が分裂病者の認知障害の本質を突いて、「現実との生ける接触の喪失」と言って自閉症の本質ないし分裂病心性の基本的概念を提示したが、これは言葉をかえて言えば、「自分が」「現在」「比処に」「居る」という正しい時間、空間を環境の内で情緒的にも安定した自己として認知することが障害された結果であるとも言えないだろうか。
連合野の機能が発揮されるためには、後連合野と辺縁系皮質および前連合野との関連、さらには、前連合野と辺縁系皮質との関連が正しく機能していなくてはならない。これらの関連を生理学的に研究し、明確な結論を引き出すことは現在むづかしいが、解剖学的には、これらの領域は神経繊維によって互いに結合していることが明らかにされている。問題はその構成を霊長類で詳しく調べて、ヒトの脳の前脳機能の基盤を類推することを可能にし、さらに大事なことは、ヒトの非侵襲的検査のデータとつきあわせてその正常および異常(機能の乱れ)の状態を研究することができるようにすることである。
前頭葉の働きを大胆に表現すれば、"低次"から"高次"までを含む発展段階に相応した"運動関連系"の範疇に属するものの総体であると言ってもよいであろう。つまり、病的機転としては、運動麻痺も運動性言語障害も自発性減退もすべて能動的機能の低下または障害として把えられてよい。
基礎的理解として、前頭葉の構造上および機能的な部位局在が、これまた、質的に程度の差こそあれ、明らかに存在することは容認されよう。
Pavlov (1927) の大脳皮質構成に関する動的モザイク説(皮質領野内で、興奮領域と抑制領域とが静的に定常化しているのではなく、その部位と範囲が瞬間、瞬間にダイナミックな変化をするという説)が今やCT(computed tomography), rCBF(regional cerebral blood flow), PET(positron emission tomography), NMR(nuclear magnetic resonance)などの近代的武器を使うことによってヒトの脳を対象にして日常臨床の場で検証される時代に入っている。
ヒトの大脳皮質は、動物の脳と同様に、いや、それ以上に、複雑に走行する皮質間繊維によって結合されている。これらの連合繊維が離断されたときに現われる機能障害(ないし症状)は、それらの起始部位、障害部位、終止部位の少なくとも3つの脳部位に起因する機能障害がミックスされ修飾された形で症侯(群)として観察されるものであろう。この切断症状群 (disconnection syndrome,Geschwind,1965) の場合、神経学で問題とされるいわゆる器質的な切断のみではなく、機能的な切断をも広く含んでいる。これによる機能障害は、episodic に一過性に出現し、概して回復が可能な経過をとるものである。臺(1979)は、切断症侯群には「解離現象」と「照合障害」の二面を区別し得るとして、以下のごとき説明を加えている。すなわち、'解離とは、一連の機能系が他から隔離して支配的となり、他の機能系への転換可能性が制限されること'を言い、'照合障害とは、認知面では、外界からの知覚情報とイメージやシェマ(Piaget)との照合、注意の選択的関門、瀘過作用、記憶面では再認や想起、思考面では、論理的整合性や構想の検討、行動面では、企図と遂行のチェック、訂正など、総じて脳内の情報処理過程のうち、異同(= or ≠)の判別を明らかにする機能の障害'を指している。また、われわれは、"局在"の問題にしても、理論上でなく実際上の問題として、形態的局在から、機能的局在、さらには症候的局在に至るまでの異なる次元での問題を統括して考察できるレベルにまで脳の研究が進歩することを期待するものである。
Kraepelin 以来、精神分裂病は精神内界の失調 (intrapsychische Ataxie) であるとか、知・情・意の乖離(Spaltung)とか看做されてきた。'知'とは意識と言語活動を基盤としてはじめて成立する抽象的認識の上に形成されるものである。情'を根源的(一次または低次)なものと高等(高次)なものとに便宜的に分けられるとして、前者は間脳および辺縁系のレベル、後者は前頭連合野のレベルがその責任領域ということになろう。そして、この両者は視床背内側核、視床前核、帯状回を介して互いに結びついている。'意'の成立機序は不明の点が多いが、'情'と密接に関連している(上述)。帯状回前部-補足運動野・運動前野を中心とする前頭葉皮質全体の機能水準と関係するとの説もある。とすれば、diffuseに分布しているカテコールアミン系やインドールアミン系の分布の変化(病的機転でこの変化がみられるであろうことは充分に考えられる)にともなうシナプスレベルの考察も重要であろう。
いずれにせよ、認知・記憶ニューロンの機能的な集団(module)間の機能的な連絡が構造的に変化することによって、記憶、ひいては認知のパタンが変化し、一面高次になり、またそれが'乱れ'たときには認知障害を惹き起こす。これらを支えている神経連絡の構造的変化が、新しい神経回路の形成によるとは到底考えにくい。おそらく外界からの強い連続的刺激が加わったとき、シナプスを構成する要素の修飾が起こり、シナプスにおける伝達効率の持続的変化を惹き起こすと考えられる。このような修飾の様式として、伝達物質受容体の数の変化、受容体の親和性の変化や、シナプス後膜のイオンチャンネルの透過性の変化が起こることが、Aplysiaのような下等動物から哺乳動物にわたる中枢神経系で明らかにされている。受容体やイオンチャンネルの特性の持続的変化を惹き起こす分子機構の一つとして、蛋白質リン酸化酵素の活性化を介して、受容体やイオンチャンネルを構成する蛋白のリン酸化による立体構造の変化が誘導されることが示されている。この蛋白質リン酸化酵素の活性化は、神経興奮によってニューロン内に生成されるセカンドメッセンジャーや、流入するカルシウムイオンによって惹き起こされる。このようなプロセスが認知機構の適応性変化に対応するものであり、外界との適応性の獲得の基礎となっていると考えられる。そして、精神分裂病で認められる、線条体および側坐核におけるドーパミン受容体の数の増加は、病因であるかあるいは病態の結果であるかは現在不明であるとしても、シナプスにおける分子レベルでの異常が外界との適応性の破錠として反映される機序の解明へのいとぐちになると思われる。現在の神経科学は、繊維連絡とシナプス活動の分析的解析から、それらの総合的機能であるヒトの高次神経活動や、その破綻のメカニズムを解明できる地点までには、かなり遠くにあり、近くには到達していないように思われる。しかし、だからと言って、精神の異常と物質的基盤に立った形態・機能の異常とを乖離させるのは、自然科学の領域へ不可知論や観念論がしのびこむ余地を生ぜしむるものであり、正しい科学的発展を阻害するものとなろう。
嗅覚的表現で稿を終わるのは本意ではないが、分裂病の認知障害の本質にせまるために臭う相手を指定するとすれば、敵は辺縁系(海馬、扁桃体、視床下部)と連合野あたりにあるように思われる。根拠に乏しいが、筆者の嗅覚をとくに強く刺激するターゲットは扁桃体・視床下部投射系(分界条、stria terminalis はその一つ)であり、これらを視床下部の諸核の発達と対応させて研究することによって、何らかの手がかりが得られるような気がしてならない。もちろん、ここで視床下部の機能について論じる際、単に喜怒哀楽とか食欲、性欲などの、いわゆる動物性機能のみでなく、生体(有機体)の内部環境の恒常性を支配する植物性機能や液性伝達経路の機能についても言及することは、個体全体の統一性を考える上で欠くことのできない視点である。種々の理由で、この領域に踏み込めなかったが、いずれ視床下部を中心とした分野で仕事をする機会があれば、そのときに考えてみようと思っている。
神経科学の一分野である(生物学的)精神医学が自然科学の分野の成果を広くとりいれ、それを基盤にして観念論哲学や機械論的認識論の影響を排除し、ヒトの高次神経機能の異常にせまって行くことが現在の重要な課題であると思われる。
生物学的精神医学 Vol. 4,
[認知機能からみた精神分裂病](日本生物学的精神医学会
小島卓也・大熊輝雄 編)学会出版センター,1993 より許可を得て転載