
哺乳動物の扁桃体(扁桃核はその組織学的名称)は終脳の半球胞の腹側壁が側脳室の内腔に隆起状に発達した神経節丘の一部から生じる。側頭葉の発達・形成とともに神経節丘の後腹側が前方に移動し、扁桃体は側脳室下角の前端の前上部に位置するようになる。ヒトの扁桃体は側頭葉前部の海馬旁回鈎のすぐ下方にみられる。海馬は終脳の蓋板につづく半球内側面の部分が翼板の肥厚によって形成される“原始”皮質の一部で、発生初期には脳梁の背側に位置している。やがて脳梁に同伴して背後方に発達し、さらに腹前方に進み歯状回となり、また側脳室内に突出して海馬足(固有の海馬、アンモン角)を形成する。扁桃体と海馬は位置的に近接しており、"関係が深い" とよく言われるが、発生の過程からみると互いに独立している(図1)。
大脳皮質との関係でいえば、海馬体とは嗅内野・海馬台(ヒトでいう海馬旁回)を介して広汎に結合する連合線維(皮質-皮質結合)の系である。他方、扁桃体との関係は投射線維(皮質―皮質下結合)の系であり、側頭葉極や下部側頭葉皮質や前頭葉の腹側部および眼窩面皮質など、情動機能に関連する領域と相互に結合している。そのうえ、扁桃核は味や臭いや自律神経系の皮質下核と結びついている。後述するように、記憶変換器としての海馬体系と感情表出複合体としての扁桃体系の働きを結びつける主要な接点は嗅内野ないし海馬旁回にある。
本論文では、記憶、情動(喜怒哀楽の感情)、認知の基盤を支えている扁桃体について、形態・機能的な面から、1)動物(ラットやサル)実験で得られた知見が人間の精神機能を解釈する上で適用できるかどうか? 2) 総説的な記述ではなく、形態・機能を中心とした基礎的研究およびヒトの脳画像解析や臨床所見から得られる扁桃体に関わる構成(神経回路)の2点に焦点を合わせて考察を試みる。
扁桃核には、形態の大きさ、区分、細胞・線維構築などの差異が動物間に認められるが、大別して主核(外側基底核と基底核)とそれを囲む発生学的に古い亜核(背側に中心核、背内側に内側核、内側に皮質核)から成る。最近、扁桃体についての総説的文献[Aggleton (2000), Gloor(1997), LeDoux (1998, 2000), Pitkänen (2000), Sah ら(2003)]が多く出版されている。ヒトの扁桃体形態に関する記述は、DeOlmos (2004)の論文に詳しく、とくに参照されたい。
扁桃体には、味覚、嗅覚、内臓感覚、聴覚、視覚、体性感覚などあらゆる種類の刺激が、嗅球や脳幹から直接的に、そして視床核(視覚、聴覚などの特殊核近傍ニューロン群)を介して間接的に入力される。その他に、大脳皮質内で処理された情報および海馬からの情報が扁桃体に入ってくる(ちなみに、これらの領域間の情報の流れは両方向性である)。この後者の入力は梨状前皮質、嗅内野(28野)、海馬台(ヒトの海馬旁回近傍)、帯状回(とくに24野)、側頭葉、前頭前野から扁桃体の基底外側核に至るもので(Kawamura & Norita, 1980; Aggleton ら,1980)、前者の皮質下からの入力に比べて時間的に遅れて、より適正かつ精緻な情報が入力される。これら2種類の粗と精、「低位」と「高位」の経路を通過した情報が扁桃体で出会う。そこでは、環境に対して瞬間的、反射的に反応した生得的な生体反応は、皮質レベルで認知された「高次」の情報に基づいて、益(報酬性)か害(嫌悪性)であるかが環境適応的に判断され修正される。同時に、海馬は新皮質からの認知情報を受ける前に情動的情報を含んだ扁桃体からの入力を受ける。この海馬・扁桃体間の関係は相補的である。
なお、モノアミン系の入力として、1)ドーパミン系: 黒質緻密部(A9)および腹側被蓋野(A10)から起こる中脳辺縁皮質系の一部が、2)セロトニン系: 中脳背側縫線核(B7)および正中縫線核(B8)から起こる上行性セロトニン束の一部が、3)ノルエピネフリン(=ノルアドレナリン)系:青斑核(A6)から、4) コリン系:マイネルト基底核(Ch4)からの入力が扁桃体に送られてくる。
大別して3つを挙げる。すなわち、①扁桃体の中心核(および一部、内側皮質核)から起こり尾状核と視床の境界部に沿って背後部から前腹方へと走って中隔核、視床下部(前核、腹内側核、弓状核)、内側視索前核などへ終止する分界条(stria terminalis) という神経線維束、および②主として基底外側核を出て内側に走り、側坐核や外側視床下部から内側部にかけて分散状に分布する腹側投射系(ventral pathway、または腹側扁桃体遠心路ventral amygdalofugal projection)と呼ばれる投射路、さらに③広範囲の大脳領域へ終わる皮質投射がある。とくに側頭葉(TE野、Boninと Bailey, 1947,による略字命名、以下同様)梨状葉皮質、前帯状回、眼窩面皮質(OF野)との結びつきは強い。側頭葉については、扁桃体の内側基底核はTE野の腹側域とのみ、外側核はTE野全体と各々結びついている(Saleem, 2000)。
メダカの扁桃体に相当する領域とその近傍を両側性に破壊すると群集行動(群れの形成)ができなくなる。コミュニケーション障害の研究のモデルとして使用可能である(坪川ら、1999)。
両側の扁桃体が破壊されたサルは、対象物の生物学的意味認知が障害され、食べられないものでも手当たり次第に口に運んだり(精神盲、口唇傾向)、同性に対しても交尾行為を仕掛けたり(性行動の亢進)、以前恐れていたヘビやヒトに平気で近づく(情動反応の低下)ようになる。クリューバー・ビューシー症候群と呼ばれている[Klüver-Bucy,1939]。てんかん治療のために、両側側頭葉切除手術(海馬破壊)を行なった後に、扁桃体の破壊をも伴うために、また、この部位の変性疾患の際にも似たような症状がヒトにも現われる (後述)。
ラットに音を聞かせると同時に電気ショックを与えて聴覚条件づけ(恐怖条件づけ)を行なうと、刺激条件(音)だけで血圧上昇やすくみ反応が起こるようになる。扁桃体が破壊されると、この恐怖聴覚条件づけ学習は障害される。この恐怖条件づけによって成立する情動反応の発現は、動物一般にみられるもので、その成立の機制はヒトとラットで基本的に違いはなく、自律神経反応(心拍数の増加、過呼吸、血圧上昇、発汗、立毛、緊張感、不安感など)を伴う「恐怖症状」や「パニック発作」が観察される。
扁桃体は情動記憶の獲得・固定・保持にも関与している。感覚条件刺激や強化刺激に誘発された快・不快感あるいは嫌悪性の痛覚情報や報酬性の味覚情報が、条件づけにより、同期的に扁桃体に入力されて、長期記憶などの機序により感覚線維-扁桃体ニューロン間のシナプス結合が強化され、扁桃体ニューロンが条件刺激に応答するようになる。また、扁桃体は、中隔-海馬体系における学習性のシナプス応答の変化を増強させ、扁桃体で得られた情動記憶を長期記憶として大脳皮質に移行させ、海馬からあらゆる感覚刺激や環境状況に関する情報を受けて、それを価値評価し意味づけを行なっている(小野と上野、2003)。
さらに、条件反射第二信号系が発達したヒトの段階になると、その発現過程に認知機構が大きく関与する。たとえば前部帯状回皮質や海馬の障害(萎縮)との関連が注目されているPTSD(post-traumatic stress disorder)では、思い出したくない無意識的なフラッシュバック(再体験現象)のみならず、言葉による刺激によっても病的な情動反応が引き起こされる(Yamasue ら, 2003)。つまり、概念化された表象がトラウマ(心的外傷)となる。精神分析による治療法として、言葉を用いてこのトラウマを消去する方法をとるが、そこでは、意識的な洞察と評価が必要で、側頭葉の記憶系と前頭葉を含む皮質領域の認識系が共に働いて、意識的記憶を用いて大脳辺縁系が関わる情動の働きを制御している。
ウルバッハ‐ビーテ病(Urbach-Wiethe disease)はヒトの臨床例に稀にみられる、常染色体劣性の代謝異常の一つで、脂肪が体内組織に蓄積し、カルシウム沈着を皮膚や喉頭部に、さらに脳では両側扁桃核に起こす疾患である。患者は、他人の顔を認知することはできるが、他人の示す表情、とくに恐れの表情を読み取ることが困難となる(Cahillら, 1995, 1996)。32歳の女性S.M.の文献上の症例もそれで、「恐れ」を認知できず(他の感情表現は普通に読み取れる)、社会的危険に対して無防備であるから、すごく社交的で、無遠慮で騙され易い。つまり、社会的行動はその器質的感情障害の故に一見して前向きであった(Adolphs ら, 1994; Damasio,1999)。病因は異なるが両側扁桃体損傷例で「恐れ」に加えて軽度に「怒り」や「嫌悪」の表情をも認知し難かったという、似たような症例D.R.も報告されている(Young ら、1995)。また、健常者に幸せな表情(ポジティブ条件)と悲しみや怒りの表情(ネガティブ条件)を見せて脳活動状態を調べた脳画像研究で、ネガティブ条件下において扁桃体(とくに、なぜか左側)の活性化を示す血流の増加が観察されている(Morris ら, 1996)。
扁桃体主核(外側基底核と基底核)と側頭葉(とくに前方部)および前頭葉(とくに眼窩部皮質)の間は相互に結合されている。ほかに、側頭葉下部(紡錘(状)回)や中古皮質(帯状回)との結びつきが注目されている。大脳新皮質と辺縁系との相互連絡は動物が高等になるにつれて発達し、とくに側頭連合野・前頭前野と扁桃体、海馬を含む辺縁系構造物との間の線維連絡は密になる(川村、1977; Van Hoesen, 1982)。
側頭連合野に属するこれらTG野やTE野前方部は、鈎状束(fasciculus uncinatus)と呼ばれる両方向性の連合線維によって前頭葉腹側部および眼窩面皮質(OF野)と相互に強く結合しているのみならず[Kawamura & Naito, 1984; 未発表例Com80]、TG、TE、OF野の各皮質はいずれも扁桃核との密接な相互結合をもっている [Kawamura & Norita, 1980]。
扁桃体は外界からの情報を価値評価して、側頭葉に蓄積されている長期記憶のバッファーの中から適宜マッチするものを選ぶべく、情動に関連した刺激に注意を向けてそれを正しく認知する上での機能をたかめる。大脳新皮質には扁桃体からの入力の他に、覚醒を賦活する脳幹網様体からの上行性投射が存在し、さらに、大脳基底部(basal forebrain)からのアセチルコリン系、黒質緻密部からのドーパミン系、縫線核からのセロトニン系の皮質全体、とくに前頭前野、帯状回前部への強い投射がある。これらの神経終末から放出される神経伝達物質は皮質ニューロンに働き、脳を覚醒状態に保ち、刺激に対する感受性を高めて、外界からの情報を処理すべく脳内のシステム効果を有効に活用できる仕組みになっている。とくに扁桃体や海馬からの情動・認知・記憶・価値判断に関する情報が前頭前野へ入力されることによって皮質が活性化され、ワーキングメモリーが適正に実行される[Goldman-Rakic ら, 1984]。

扁桃体から起こり視床下部に終わる神経路には、分界条と腹側扁桃体遠心路があることは既に述べた。視床下部は呼吸・循環、摂食、性機能、水分代謝などの自律機能や脳下垂体の機能を促進または抑制する内分泌調節などを司る生命維持に関わる重要な領域で、外的または内的な誘因が存在するとき、視床下部への刺激が来たとき周囲への働きかけの動因となり、これによって動物は行動を起こすことになる。分界条を形成する軸索終末は視床下部の腹内側核(古典的「満腹中枢」)や外側野(古典的「摂食中枢」)を含む摂食関連領域のニューロンとシナプスを作る。これらの領域にあるニューロンはグルコースに反応するが、同時に身体の脂肪細胞から分泌される肥満因子レプチンに対して腹内側核には促進的に、外側野には抑制的に反応する受容体をもつものも存在する。海馬からは脳弓を経由して外側乳頭体核へ、網膜からは視交叉上核へ、さらに脳幹からは視床下部の広い領域へアミン系線維の投射が行われている。
視床下部の機能活動は、種々の脳部位、すなわち、①大脳皮質とくに前頭前野、②扁桃体、③海馬、④脳幹から入力されるさまざまな情報によって制御されている。外界の対象物(刺激)に対して、それらが自己(動物)にとって益になるか害になるかの価値判断を扁桃体が行なっている。益になると判断されるときには生体に快情動が起こり、反対に害と判断されると不快情動を伴う。
視床下部が司る自律機能は大脳皮質、とくに前頭葉からの直接的な影響下にある。すなわち、その特徴は、視床下部から前頭前野への上行性投射は広く分散しているが、逆方向の前頭前野から視床下部への投射は主として内側面皮質と眼窩面皮質から起こり、扁桃体からの線維束である分界条の終止域と重複している(Rempel-Clower と Barbas, 1998)。
こうして、情動と認知を基盤として、動物が判断し行動に移すまでの過程には視床下部求心路が関連し、判断結果を行動に移す形態基盤には、「運動に関連する視床下部遠心路」とくに脳幹・脊髄の運動神経細胞に軸索終末を送る視床下部・被蓋路、視床下部(室傍核や外側野から)・脊髄路、さらに網様体・脊髄路などの下行投射路が関与している(佐野、2003、参照)。
また、意識を支える機構として眺めると、視床下部は内臓感覚などの刺激を受けて直接大脳皮質を賦活すると同時に、非特殊性・広汎性の網様体賦活系にも興奮を伝えており、視床下部調節系と脳幹(上行性)網様体賦活系とは互いに関連しつつ、意識レベルを調整していることが知れる。意識への科学的アプローチはこれからである。
20年前に筆者は総説「大脳辺縁系の解剖学」(川村と小野、1987)を執筆したが、その中で海馬と扁桃体の関係を「図4:大脳辺縁系の見取図」に示した(川村:「脳と精神」、2006の 図32)。本誌に改変して図3として掲載する。その後、扁桃体と海馬の間の関係は、両者からの情報がヒトの脳では、嗅内野近傍、海馬旁回近傍( 動物実験で明らかにされた嗅内野、嗅周野皮質領域)で出会うという研究(Aggleton,1986; Saunders と Rosene, 1988; Saunders, Rosene と Van Hoesen,1988)が詳細になされ、さらにこの10年間には広範な総説や原著論文(Stefanacci ら, , 1996; Pitkänen ら2000; 2002; Majak と Pitkänen, 2003; Majak ら, 2004 ; Kishi ら, 2006)が発表された。この問題については、本特集号で執筆される小野論文を読まれたい。
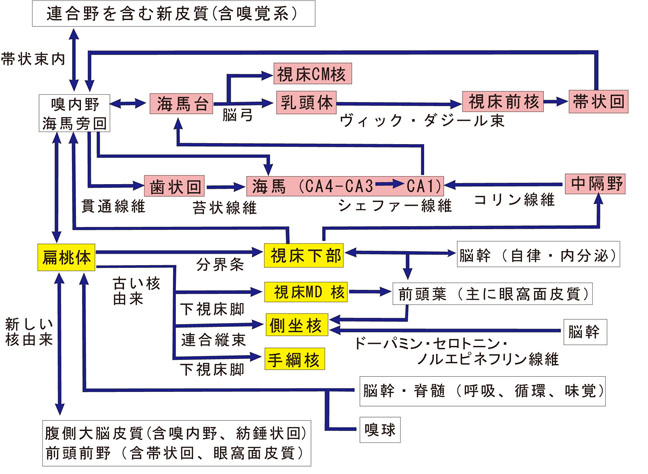
嗅覚や味覚の情報は視床経由で大脳皮質で分析されるほか、扁桃体でも処理される。すなわち、嗅覚刺激は嗅球から、味覚刺激は脊髄・脳幹レベルからともに直接、扁桃体の系統発生的に古い部分の皮質核・内側核群に入力されており[Norita & Kawamura, 1980] 、これらの感覚系の情動への関与も大きい。水棲動物では嗅覚も味覚も同じ化学物質が各感覚細胞に興奮を与える。これらの動物では、聴覚は、水圧や波動感覚や味覚と同様に、側線系がこれに相当する。このように、嗅覚や味覚は、とくに下等動物において、辺縁系/情動と深く関連している。また、脳幹部にはモノアミン、ペプチド、コリンなどを含む神経細胞や線維が存在しており、自律系・内分泌系・情動系の中枢部である視床下部/扁桃体に入力線維を与えている。
興味深いことに、種々の神経伝達物質、①ドーパミン系、②セロトニン系、③ノルエピネフリン系という生物学的基盤があるとして、社会的、行動的、心理的、各次元の総合である人格(パーソナリティー)のモデルの一つとして、各々、①新奇性探求(novelty seeking)、②危機回避(harm avoidance)、③報酬依存(reward dependence)と関連づけられて、一見単純な形で提唱されているものがある(Cloninger, 1987)。ところで「人格」とは、性格や性質とは異なり、感情/情緒を伴った自我意識をもって持続的に統一しており、しかも行動的特性の全体を表わすものである。従って、精神医学的にみた人格障害の生物学的見解を求められる時には、(思考・意志/行動決定・理性に関わる)皮質連合野や(動機づけ・モチベーション・情動行動に関わる)大脳辺縁系の機能を考慮して論じたいものである。
気質、病質、人格とその異常/障害に関する問題は、Schneider K (1923)に代表されるように、対象を平均基準からの偏倚かつ変異概念として捉えて、類型学的記述にとどまったが、現代の科学水準に立った精神医学が社会の要請にどのように応えられるか大きな課題の一つであろう。
大脳基底核は、扁桃体から情動系の入力(Wright ら, 1996)が、またさらに新奇物体に対する意欲的行為に関与する中脳の黒質からのドーパミン入力(Li ら, 2007)が、終止する場所である。とくにその腹側部には側坐核(=腹側線条体)と呼ばれる細胞集団が見られ、背外側の核部(core)と腹内側の殻部(shell)に区分されている。そして線維結合上、前者は運動に、後者は情動や自律系に関連が強いと考えられている。shellには豊富なドーパミン線維とドーパミン受容体(D4R)の強い発現が認められる。この領域のニューロンは、形象を(サル)自身にとって意味のある空間内に認知し、その方向に向かって意欲的に行動を開始したときに活動する。側坐核へは、脳幹・脊髄から自律神経系の刺激が、また終脳からは分析・処理された形の感覚刺激が伝わる。その他、扁桃核から分界条を経由する興奮性の情動入力が加わる。そしてこれらの入力刺激はドーパミン軸索終末からの放出を状況に応じて制御し、さらに腹側淡蒼球のグルタミン酸作動性ニューロンを介して間脳や脳幹の諸核に興奮を伝えている。このように腹側線条体は大脳辺縁系から情動に関する情報を受けるとともに、報酬を期待することによって動機づけを評価し、その結果を運動情報に変換して情動行動に関与している。近年、機能的核磁気共鳴画像(fMRI)を用いて意欲、感情、報酬(および薬物嗜癖)などとの関係が研究されているが(Tanaka et al, 2006; Hampton & O'doherty, 2007)、扁桃体—側坐核—腹側淡蒼球—視床背内側核—眼窩回・帯状回のループはその中核をなしている。
扁桃体は上に見たように、広範な大脳皮質領域から脳幹・脊髄までの中枢神経系(CNS)全体にわたり、神経回路網によって複雑に結びついている。これに加えるに、内分泌・自律神経系やペプチド・モノアミンなど液性伝達系をも含めると、CNS全体を対象としなければならない。断片的に概説するのではなく、動物一般の扁桃体の生理機能を総括的に論じることは重要ではあるが、本論文の目標とはなりにくい。
ここでは、高等動物とくに人間の扁桃体について考えてみたいので、ヒトで著しい発展を遂げた大脳皮質レベルに話題を絞りたい。改めて扁桃体は、前頭前野の眼窩面(orbitofrontal)、内側面(medial prefrontal area、とくに帯状運動皮質cingulate motor cortex)、側頭葉の前方域(temporal pole)、内側(hippocampal formation)、下面(とくに紡錘状回近傍fusiform gyrus)、と強く結合している。そして、これらの皮質域間の関連性も考慮しながら、異現同根で不可分の関係にある情動機能と認知機能の周辺に関心は払われる。
現象的トピックスとして以下の3点、すなわち、①「原始的感覚」としての嗅覚と情動と生命を支える自律機能、② 側頭葉の紡錘状回が関わる表情の認知、③ 前頭前野(内側面および外側面)が関わる能動性・「社会脳」機能、でまとめてみる。
動物は餌を捕り、敵から守るために、最初に嗅覚と味覚を発達させた。これらの感覚刺激は扁桃体に、そこからさらに視床下部へと伝達される。大脳皮質を持つ動物になると、扁桃体から皮質に終わる神経路、さらに皮質・視床下部路が形成されてくる。これらの線維の終始域は前頭葉内で、嗅覚皮質は眼窩面に、味覚皮質は弁蓋部(43野、一般動物では島皮質)にあり、視床経由のいわゆる上行性感覚伝導路もここに終わる。この事実から、これらの皮質領域が呼吸・循環を含む自律機能および「原始的感覚」の情動と認知・分析のはたらきに関与していることが言える(生理学的研究もこれを裏付けている)。
側頭葉前下面皮質(TE野)は扁桃核からの入力もうけ、視・聴覚をはじめ多くの情報を統合して情動および認知記憶など多様な機能を果している。また、TE野には上側頭溝周囲皮質とともに相貌認識に関与する顔選択性細胞(face-selective cell)が見出されている [Perrett et al., 1982] 。顔認識や表情処理の研究は社会性の適応障害の問題に直結する。ヒトの顔の表情を認知する領域としては、側頭葉下面で後頭葉に連続している内側後頭側頭回(=紡錘状回)が知られているが(後述)、英国精神医学研究所のフィリップスのグループは上側頭溝周囲の皮質領域を中心に、健常者および精神異常者を対象に情動に対応する顔の表情変化の際に起こる神経活動について研究している(Critchley et al. 2000)。
サルで軸索流法を用いた研究から、扁桃体と前頭前野眼窩面および内側面皮質との間には強力な結びつきが両方向性に、そして外側面皮質との間には弱い結びつきが一方向性に、認められる(Ghashghaei & Barbas, 2002)。また、扁桃体から強い投射が前部帯状運動皮質(M3)に、弱い投射が後部帯状運動皮質(M4)に行われている(Morecraftら, 2007)。さらに扁桃体と側頭葉、とくにその前方部、との間には強力な相互結合の存在も示され(Kawamura & Norita, 1980;Ghashghaei & Barbas, 2002)、ヒトにおいてもこれらの領域間の相互関連性が顔面表情認知の過程でfMRIを用いたイメージング(画像解析)の結果が明瞭に示されている(Iidaka ら, 2001)。
ところで、他人の心を推測し得る「内部モデル」なるものが幼児期の脳内に形成されるという。寸劇を見せるという「心の理論(Theory of Mind, ToM)」の課題遂行実験[Baron-Cohen ら, 1985]は、歌舞伎の「助六」にも桶を取り違える見せ場があるが、他者の心の内面すなわち心理状態を想定するもので、コミュニティー活動を支える精神的活動を反映する「社会脳機能」を診るもので、役者の心の中を4-5歳になると察し得ると言う。前頭前野・側頭葉・扁桃体の間のニューラルサーキットは上に述べたように互いに関連し、活動している。前頭前野、前頭葉の腹内側部および眼窩部、上側頭溝および回の領域、紡錘状回、扁桃体を結ぶこれらの神経ネットワークの構成は、この「社会脳」の認知・情動機能を支える生物学的基盤をなしており、その機能低下は社会性の獲得障害として自閉症との関連が検討されている(Baron-Cohen et al, 1999; Bachevalier, 2000)。
なお、臺(1979,1991)によって指摘された統合失調症にみられる「履歴現象」、すなわち想起される記憶内容が新たな情動反応と結びついて病状が再燃することは、この「社会脳」の主要な要素である扁桃体の機能的役割を再認識させる。
臨床的に単極性うつ病者で[Drevets ら,1992]、また健康人でも悲しみや喜びなどを表わす情動的な陳述記憶を思い浮かべた場合に、扁桃体で脳血流が増加していることがPETやfMRIを用いた研究で報告されている。情動記憶と扁桃体の関係から明らかなように、ヒトの場合、益か害かの判断は実生活上の実利的なものに限られるものではなく、周囲(社会)の情報を処理し、広く動因なるものを選定して、また芸術などに接した場合に知覚される高尚な快の情感、つまり精神的な意味で生体にとって有益な情感の判断もここに含められよう。
最近、ヒトを用いた脳画像解析研究により、紡錘状回の働きは静止顔面像の特徴を情報処理して、個人の相貌を認知する領域に相当することが明らかにされた [Kanwisher et al., 1997; Farah & Aguirre, 1999]。相貌認知は社会的コミュニケーションの上で大切な非言語性の要素であるが、自閉症患者でこの紡錘状回の活動が著しく低下していることが指摘されている[Schultz et al., 2000 ; Pierce et al., 2001]。
双極性障害を対象にMRIなどの脳画像研究もなされており、扁桃体の体積の増減について健常者と比較した報告が多くみられるが、症状の変化のためか、一定した所見が得られていない(鬼塚と神庭, 2006,参照)。この種の研究結果の解釈は病態の多様性と経過の変動などが常につきまとうので、病気のタイプによる診断別ではなく、病状経過のステージ別に注目した調べ方が望ましい(Tebartz van Elst ら, 2007)。ごく最近、早期発症(ハイリスク病態、初期エピソード)から慢性に経過した統合失調症までの症例を対象に、治療とその転帰を追跡して扁桃体と海馬の体積変化を調べたメルボルンのグループの研究(Velakoulis et al, 2006)がある。今後、早期治療や社会復帰と結びつけるべく進展することが期待される。
知覚連合野からの情報が前頭前野に伝達され、そこで処理される過程で、情報の価値評価/意味認知を行なう扁桃体を主体とする大脳辺縁系と、情動表出/発現を行なう視床下部を中心とする感情出力系の両システムを、大脳新皮質連合野が調節する機構が発達する。この制御・調節系が柔軟かつ適正に働くように形成されることが大切で、このコントロールシステムに総括的に関わる前頭前野が、関連する神経回路網の髄鞘化を含めて、成熟するのが思春期に当たる。この系における応変の可塑的応答性の崩れが精神疾患の根底に在る。
動物一般にみられる扁桃核は、味や臭いや自律機能と結びついて情動機構の発現に皮質下レベルで関わると同時に、皮質-扁桃核間の神経回路が働くことによって、賞罰、報酬などの意味づけ、動機づけなどの形成がなされる。人間には金銭欲、支配欲、名誉欲など前頭前野が関わる「社会・生物学的」レベルの欲求があるが、これも皮質連合野、扁桃体、視床下部が参加する「価値判断と能動性」機構によって支えられている。
ところで、情動と認知およびそれらの相互作用の基礎にある高次神経活動は、皮質にある二つの条件反射系(感覚系と言語系)と皮質下にある無条件反射系を含んでいる。観念は常に言語による抽象作用であるので、ヒトでは、言語信号系が認識の基礎をなす神経活動となる。他方、情動は感覚信号系と皮質下(扁桃体、中隔核、分界条底核、側坐核/腹側線条体を含む大脳辺縁系を構成する部分)の無条件反射系の相互作用を主とするものでる。こうして、認知の機構と情動の機構の基礎にある諸過程は、互いに関連している。人間にあっては、認識(観念)と情動表現は、実在を意識内に反映するものとして不可分に結びついている。その意味で、他の動物と違って言語中枢を備えている人間にとっては、情動は意識的に認識されて意味をもつものとなる。
改めて扁桃体を中心とする情動の機構について考えてみると、認知機構におけると同様に、無条件反射的に反応するサカナやカエルのレベルから、大脳皮質が関与する条件反射的(第一信号系の)低レベル認知が関与するネズミ・ネコ、そしてサルのレベル、さらに進んで第二信号系または言語信号系が関与するヒトのレベルにまで「ヒエラルキー」が存在する。ここに動物一般に共通する側面と、特定の動物に固有に備わった側面との両面をあわせみることができる。つまり一般的に新皮質よりも辺縁系の働きに大きく依存するとみられている情動機構においても、それが言語機能と結びついたとき、量・質の両面で本質的な差異が生じてくる。つまり、情動機能は高次の認識機構がその根底にあって情動の基盤をなすときに、たとえば美術、芸術などで高レベルの機能を発揮する。
貴重なご意見を賜った臺 弘先生(坂本医院)に、また作図して頂いた船戸和弥氏(慶應義塾大学)と高沖英二氏(メタ・コーポレーション・ジャパン)に感謝いたします。
1)Adolphs R, Tranel D、Damasio H et al : Impaired recognition of emotion in facial expressions following bilateral damage to the human amygdala. Nature 372 : 669-672, 1994
2)Aggleton JP : A description of the amygdalo-hippocampal interactions in the rhesus monkey. Exp Brain Res 64: 515-526, 1986
3)Aggleton JP :The Amygdala, A functional analysis (Ed), Second edition, Oxford Univ Press, 2000
4)Aggleton JP, Burton MJ, Passingham RE : Cortical and subcortical afferents to the amygdala of the rhesus monkey (Macaca mulatta). Brain Res 190:347-367, 1980
5)Bachevalier J: The amygdala, social cognition, and autism. pp. 509-543, In: The Amygdala Second edition, Oxford Univ Press, Ed by Aggleton JP, 2000
6)Baron-Cohen S, Leslie A, Frith U: Does the autistic child have a “theory of mind” ? Cognition 21 : 37-46, 1985
7)Baron-Cohen S, Ring HA, Wheelwright S et al: Social intelligence in the normal and autistic brain: an fMRI study. Eur J Neurosci 11 : 1891-1898, 1999
8)Bonin Gv, Bailey P : Neocortex of Macaca mulata. Univ Illinois Urbana, 1947
9)Cahill L, Babinsky R, Markowitsch HJ et al : The amygdala and emotional memory. Nature 377: 295-296, 1995
10)Cahill L, Haier RJ, Fallon J et al: Amygdala activity at encoding correlated with long-term, free recall of emotional information. Proc Natl Acad Sci USA 93 : 8016-8021, 1996
11)Cloninger CR : A systematic method for clinical description and classification of personality variants: A proposal. Arch Gen Psychiat 44: 573-588, 1987
12)Critchley H, Daly E, Phillips M et al; Explicit and implicit neural mechanisms for processing of social information from facial expressions: a functional magnetic resonance imaging study. Human Brain Mapping 9: 93-105, 2000
13)Damasio RD : The Feeling of What Happens. Harcourt Brace & Company, 1999 (無意識の脳 自己意識の脳、田中三彦訳 講談社 2003)
14)DeOlmos, JS : Amygdala of The Human Nervous System, Chapter 22 2nd Ed.,Ed. By Paxinos G, Mai JK, Elsevier, Academic Press,2004.
15)Drevets WC, Vidden TO, Price JL et al: A functional anatomical study of unipolar depression. J Neurosci 12 : 3628-3641, 1992
16)Farah MJ, Aguirre GK: Imaging visual recognition: PET and fMRI studies of the functional anatomy of human visual recognition. Trends Cogn.Sci. 3 : 179-186, 1999
17)Ghashghaei HT, Barbas H : Pathways for emotions: Interactions of prefrontal and anterior temporal pathways in the amygdala of the rhesus monkey. Neurosci 115: 1261-1279, 2002
18)Gloor P : The Temporal Lobe and Limbic System. Oxford Univ Press, 1997
19)Goldman-Rakic PS, Selemon LD, Schwartz ML : Dual pathways connecting the dorsolateral prefrontal cortex with the hippocampal formation and parahippocampal cortex in the rhesus monkey. Neurosci 12 : 719-743, 1984
20)Hampton AN, O'doherty JP : Decoding the neural substrates of reward-related decision making with functional MRI. Proc Natl Acad Sci USA 23 : 1377-1382, 2007
21)Iidaka T, Omori M, Muraata T et al: Neural interaction of the amygdala with the prefrontal and temporal cortices in the processing of facial expressions as revealed by fMRI. J Cogn Neurosci 13: 1035-1047, 2001
22)Kanwisher N, McDermott J, Chun MM: The fusiform face area: a module in human extrastriate cortex specialized for face perception. J Neurosci 17 : 4302-4311, 1997
23)川村光毅: 連合野"の線維結合(I)皮質間結合−−サルとネコの皮質間結合の比較と"連合野"の発達についての試論−−. 神経研究の進歩 21: 1085-1101, 1977
24)川村光毅: 脳と精神 ―生命の響き― 慶應義塾大学出版会、2006
25)Kawamura K, Naito M : Corticocortical projections to the prefrontal cortex in the rhesus monkey investigated with horseradish peroxidase techniques. Neurosci Res 1: 98-103, 1984
26)Kawamura K, Norita M : Corticoamygdaloid projections in the rhesus monkey. An HRP study. Iwate Med Ass 32 : 461-465, 1980
27)川村光毅 小野勝彦:大脳辺縁系の解剖学. Clinical Neuroscience 5: 132-138, 1987
28)Kishi T, Tsumori T, Yokota S et al: Topographical projection from the hippocampal formation to the amygdal: a combined anterograde and retrograde tracing study in the rat. J Comp Neurol 496 : 349-368, 2006
29)Klüver H, Bucy PC : Preliminary analysis of functions of the temporal lobe in monkeys. Arch. Neurol Psychiatr 42 : 979-1000, 1939
30)LeDoux J : The Emotional Brain. Weidenfeld & Nicolson, London, 1998 (エモーショナル・ブレイン、情動の脳科学、松本元、川村光毅ほか訳、東京大学出版会、2003)
31)LeDoux JE: Emotion circuits in the brain. Ann Rev Neurosci 23:155-184, 2000
32) Li HP, Homma A, Sango K et al: Regeneration of nigrostriatal dopaminergic axons by degradation of chondroitin sulfate is accompanied by elimination of the fibrotic scar and glia limitans in the lesion site. J Neurosci Res 85: 536-547, 2007
33)Majak K, Rönkko S, Kemppainen S et al: Projections from the amygdaloid complex to the piriform cortex: a PHA-L study in the rat. J Comp Neurol 476: 414-428, 2004
34)Majak K, Pitkänen A: Projections from the periamygdaloidcortex to the amygdaloid complex, the hippocampal formation, and the parahippocampal region: a PHA-L study in the rat. Hippocampus 13: 922-942, 2003
35)Morecraft RJ, McNeal DW, Stilwell-Morecraft KS et al : Amygdala interconnections with the cingulate motor corex in the rhesus monkey. J Comp Neurol 500: 134-165, 2007
36)Morris JS, Frilt CD, Perrett DI et al: A different neural response in the human amygdala is fearful and happy facial expressions. Nature 383 : 812-815, 1996
37)Norita M, Kawamura K : Subcortical afferents to the monkey amygdala: an HRP study. Brain Res 190 : 225-230, 1980
38)鬼塚俊明、神庭重信:双極性障害の脳画像. 臨床精神医学 35: 1433-1438, 2006
39)小野武年、上野照子:情動. 753-770頁, In: 脳神経科学、2003、三輪書店
40)Perrett DI, Rolls ET, Caan W: Visual neurons to faces in the monkey temporal cortex. Exp Brain Res 47: 329-342, 1982
41)Pierce K, Muller RA, Ambrose J et al : Face processing occurs outside the fusiform ‘ face area ’ in autism : evidence from functional MRI. Brain 124 : 2059-2073, 2001
42)Pitkänen A: Connectivity of the rat amygdaloid comples, In: The Amygdala, second edition, a functional analysis, Ed.by Aggleton, JR, pp 31-115, Oxford Univ Oress, 2000
43)Pitkänen A, Kelly JL, Amaral DG : Projections from the lateral, basal, and accessory basal nuclei of the amygdala to the entorhinal cortex in the rhesus monkey. Hippocampus 12 : 186-205, 2002
44)Pitkänen A, Pikkarainen M, Nurminen N et al: Reciprocal connections between the amygdala and the hippocampal formation, perirhinal cortex, and postrhinal cortex in rat. Ann NY Acad Sci 911: 369-91, 2000
45)Rempel-Clower, NL, Barbas, H : Topographic organization of connections between the hypothalamus and prefrontal cortex in the rhesus monkey. J Comp neurol 398: 393-419, 1998
46)Sah P, Faber ESL, De Armentia L et al : The amygdaloid complex: Anatomy and physiology. Physiol Rev 83: 803-834, 2003
47)Saleem KS, Suzuki W, Tanaka K et al: Connectiions between anterior inferotemporal cortex and superior temporal regions in the macaque monkey. J Neurosci 20: 5083-5101, 2000
48)佐野 豊:
視床下部、医学書院、東京、2003
49)Saunders RC, Rosene DL: A comparison of the efferents of the amygdala and the hippocampal formation in the rhesus monkey: I. Convergence in the entorhinal, prorhinal, and perirhinal cortices. J Comp Neurol 271: 153-184, 1988
50)Saunders RC, Rosene DL, Van Hoesen GW: Comparison of the efferents of the amygdala and the hippocampal formation in the rhesus monkey: II. Reciprocal and non-reciprocal connections. J Comp Neurol 271: 185-207, 1988
51)Schneider K : Die Psychopathischen Persönlichkeiten. Franz Deuticke, Wien, 1923, 精神病質人格、懸田克躬、鰭崎 轍 訳、みすず書房、1954
52)Schultz RT, Gauthier I, Klin A et al : Abnormal ventral temporal cortical activity during face discrimination among individuals with autism and Asperger syndrome. Arch Gen Psychiat 57 : 331-340, 2000
53)Stefanacci L, Suzuki WA, Amaral DG: Organizzation of connections between the amygdaloid complex and the perirhinal and parahippocampal cortices in macaque monkeys. J Comp Neurol 375: 552-582, 1996
54)Tanaka SC, Samejima K, Okada G et al : Brain mechanism of reward prediction under predictable and unpredictable environmental dynamics. Neural Netw 19: 1233-1241, 2006
55)Tebartz van Elst L, Ebert D, Hesslinger B: Amygdala volume status might reflect dominant mode of emotional information processing. Arch Gen Psychiatry 64 : 251-253, 2007
56)坪川達也: 社会行動と魚の脳. 慶応義塾大学日吉紀要自然科学 26 : 1-10, 1999
57)臺 弘:履歴現象と機能的切断症状群 - 精神分裂病の生物学的理解. 精神医学 21:453-463, 1979
58)臺 弘:分裂病の治療覚書. 創造出版 東京 1991
59)Van Hoesen GW : The parahippocampal gyrus. New observation regarding its cortical connections in the monkey. Trends Neurosci 5 : 345-353, 1982
60)Velakoulis D, Wood SJ, Wong MTH et al: Hippocampal and amygdala volumes according to psychosis stage and diagnosis. Arch Gen Psychiatry 63 : 139-149, 2006
61)Wright CI, Beijer AVJ, Groenewegen HJ: Basal amygdaloid complex afferents to the rat nucleus accumbens are compartmentally organized. J Neuroscience 16: 1877-1893, 1996
62)Yamasue H, Kasai K, Iwanami A et al: Voxel-based analysis of MRI reveals anterior cingulate gray-matter volume reduction in posttraumatic stress disorder due to terrorism. Proc Natl Acad Sci USA 100 : 9039-9043, 2003
63)Young AW, Aggleton JP, Hellawell DJ et al: Face processing impairments after amygdalotomy. Brain 118 : 15-24, 1995
臨床精神医学 36 (2007) 817-828 (アークメディア)より許可を得て転載。