川村光毅
サカナやトリにも認知と情動の機構は備わっていて、リズミカルに環境に反応して、互いに感情を交えたコミュニケーションをしています。イヌやサルやゴリラでは、声を変化させて音楽の一歩手前といってもいい音色を響かせて感情表現をします。犬の遠吠え、猿の奇声・嬌声、ゴリラのドラミングなどはかれらのライフスタイルが生みだした作品です。しかし、音楽と言う芸術にまで感性を高められる機能を彼らの脳はもっていません。なぜ? その秘密とは? まず、音についての認知、聴覚の話から始めましょう。
音の認知にはいろいろなレベルがあります。 以下に、1)音を聞く、声が聞こえる、 2)調べ(旋律)・言葉を聴く、 3)音楽・講演を傾聴する、4)楽器を奏でる・歌を歌う、の順に話を進めます。
音は空気中を音波(振動)として伝わり、聴覚の受容器である耳で捕らえられ、脳に伝えられて感知されます。音の振動は、耳の穴(外耳)に入り、中耳との境にある鼓膜を震わせます。鼓膜には3つの耳小骨という人体でもっとも小さな骨が結合していて、これを介して音の振動は内耳に伝わります(図1)。内耳では音の振動を電気的信号に変えて神経に伝えます。内耳には蝸牛(かぎゅう)という文字通りカタツムリの殻のようならせん状の構造があり、内部を覆う細胞には小さな毛が無数に生えています。これを有毛感覚細胞と言い、特定の高さの音は、特定の有毛感覚細胞を刺激します。その刺激は蝸牛神経(いわゆる聴覚神経)を通って脳に伝わります。
脳の内部には、無数の神経回路が存在しますが、耳からの刺激は特定の神経回路を通って、最終的には最高中枢である大脳皮質に送られます。この聴覚路と呼ばれる神経経路の途中には、いくつかの神経核(蝸牛核、上オリーブ核、外側毛帯核、下丘、内側膝状体など)とよばれる神経細胞(=ニューロン)の集まりがみられます。周波数の違いによって音の高低がきまりますが、おのおのの神経核内の場所表示は蝸牛の段階から大脳皮質のレベルまで保たれています。(注1)。
末梢レベルの蝸牛神経では音の時間パターンはニューロンの発火パターンの時間変化によって表わされますが、中枢レベルではこの時間パターンを統合した時間情報として示されます。われわれは、刻々と変化する響きの高さや強度を追って、その時間パターンの違いによって音のもつ意味を認知します。ニューロンの性質を調べてみると(周波数分析)、大脳皮質に近い上位の神経核のニューロンほどその応答野は鋭くなっており、下丘で「聴ニューロン」の応答周波数範囲が最も狭くなっています。周波数分析は中脳の下丘のレベルで完成しているといわれています。
これに対して、間脳と大脳皮質のレベルでは、スペクトルの弁別がなされます。間脳の視床にある内側膝状体のニューロンは、純音に対して音の始まりに発射するon反応、終わりに発射するoff反応、あるいは両方で発射するon-off反応を示します。そして波の形は多峰性で、下丘のニューロンの応答野よりも周波数範囲はむしろ広くなっています。大脳皮質ではこの傾向がよりはっきりしてきます。これらのことは、間脳より上位脳では、周波数分析とは異なった機能が主となり、統合作用がなされることを示しています。
内側膝状体から直接線維が終わる第一聴覚野(A1野)では音の基本的性質である音の大きさや高さ、それに音色などを認知しますが、それだけでは「音楽を聴く」ことはできません。音の一連の組み合わせを音楽として認識するためには、より高度な脳の機能が必要とされます。そのためには聴覚野のニューロンの興奮が大脳皮質の他の領域に伝えられ、しかるべき情報処理がなされなければなりません。こうして聴覚野に伝えられた情報は大脳皮質の側頭葉にある聴覚連合野に送られます。そこで、個々の音の情報が統合的に処理されます。(注2)。
以下にこれから数年後に、音大生と医学生が共同して研究できるようになったときに基礎知識となる聴覚研究の最先端のエッセンスを紹介します。
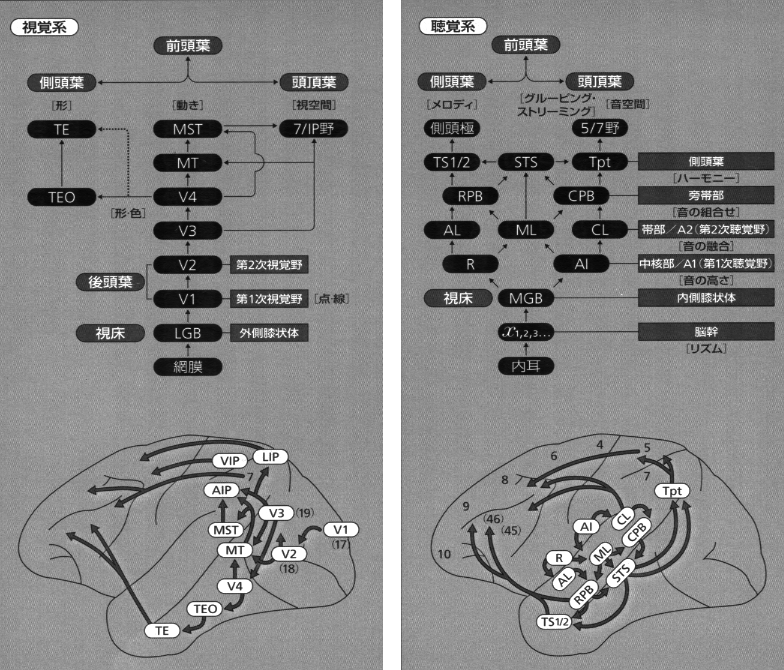
最近サルの聴覚領皮質で以下のことが明らかにされました(Romanski ら, 1999)。すなわち、Core領域(中核部、第一聴覚野;auditory core area, R; rostral core, RT; rostrotemporal core) は、各振動数に対応した単純な音をうけとり、中核部からmedial & lateral belts (内側および外側帯部、CL, ML, AL) に興奮が伝わり、さらに外側帯部から parabelt (旁帯部)へと進行する。この過程を通じて聴覚処理の内容が高められます。第一聴覚野(A1野)から belt へ軸索がのびている。そして、いくつかの音が「融合」する22野 (帯部と旁帯部) では、雑音に反応するニューロンが存在します。また、ネコに音を聞かせたとき、A1野および第二皮質聴覚野(A2野)でチューニングの幅に広狭が認められ、潜伏時間にも長短があるなどいろいろな性質をもったニューロンの存在が観察されています(He ら,1999)。
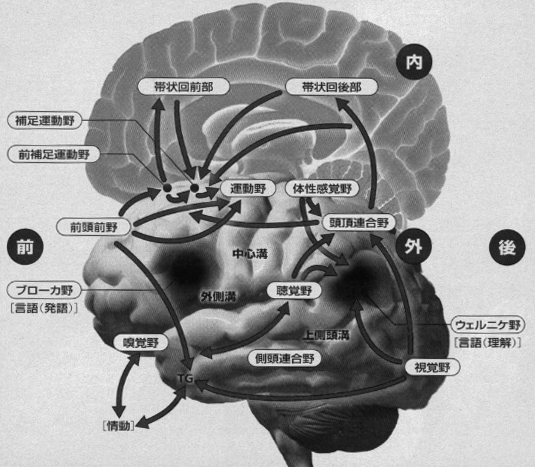
サルの側頭連合野に属するTG域やTE域前方部は鈎状束とよばれる連合線維によって、前頭葉眼窩面皮質(OF)と両方向性に強く結びついており(川村、1977)、これらの皮質 (TG,TE,OF) はともに扁桃核と相互に結合しています(Kawamura と Norita,1980)。そして、上側頭溝(STS)周囲の皮質域では、後部は言語系に、前部は情動系に関連し、中央部を含めて全域的に多種感覚性ニューロンが存在し、ここにさまざまな感覚の入力が集中して終わっています。また、ネコにも多種感覚入力が同様に終わる領域(上シルビウス溝周囲皮質)があります(Kawamura, 1973a,b,c, 1977, Kawamura とOtani, 1970)。ヒトの脳では、外側溝周囲皮質にヘッシェル回と呼ばれるA1野に属する領域があり、その後方に連続して存在する感覚性言語野(ウエルニッケ野)は、聴覚野からたくさんの線維を受けています。そして、ここはヒトに進化する過程で、サルのSTS後部皮質域から発達した皮質域と考えられます(川村、1977, 2006)。こうして、感覚性言語野と結びついた受身の情動表現は、運動性言語野(ブローカ野)と結びついた能動的な情動行動と深く関わり合っています。(注3)。
視覚系では脳内での情報処理の過程がかなり明らかになっています。目の網膜で捕らえた光の情報は視神経を伝わって、間脳の視床にある外側膝状体という部位に送られます、そしてそこから大脳皮質の後頭葉の視覚中枢に達します。ここで光情報は映像として再構成されますが、まだ映像の意味づけはできません。映像の情報は上下に別れて脳の前方へと送られ、最終的に前頭葉の前頭前野に達します。前頭葉は人間でよく発達した脳の領域で、理性、判断、学習などの高次機構を司っています。上の経路(背側路)は映像が「どこか(where)」を認識する機能(空間認知)に、下の経路(腹側路)は映像が 「何か(what)」を認識する機能(対象認知)に、それぞれ関係するといわれています(Goodale と Milner、1992)。対象認知は、個性や経験(個々の脳の成り立ち)に彩られた情動機能とも関連していて、例えば、火事を認知すると同時に恐怖や驚きを引き起こします。このように、視覚認知のなされ方は多様で、後連合野内での連合線維の働きによって、ニューロンレベルでの多種の組み合わせができ、調和した活動がなされます。
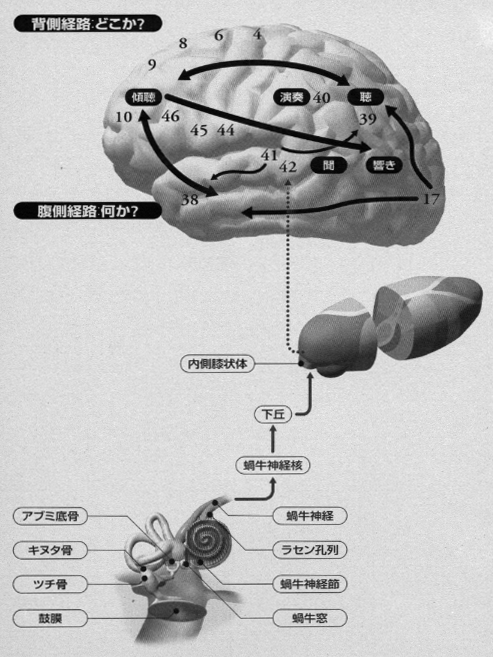
聴覚系でも後連合野から前頭前野への流れに上下(背側および腹側)の神経路があり、それぞれ、「どこか」と「何か」の機能が認められています(Romanski ら, 1999; Kaas と Hackett, 1999)。「背側の流れ」および「腹側の流れ」の概念は、もともと、後連合野内の興奮の伝達経路に対応して用いられたものですが、次のステップである前頭前野へ向かう経路をも含めてより広く考察することができます。さらに拡張すれば、前頭葉で「処理」された情報が、同じ背側と腹側の流れを通って逆方向に、前連合野から後連合野へと伝わる経路もこの言葉に含められます。視覚および聴覚の領野のブロック図と大脳皮質内の興奮の流れについては(図2)。
以下に、これまでの研究結果をまとめた考察をしてみます(図3)。
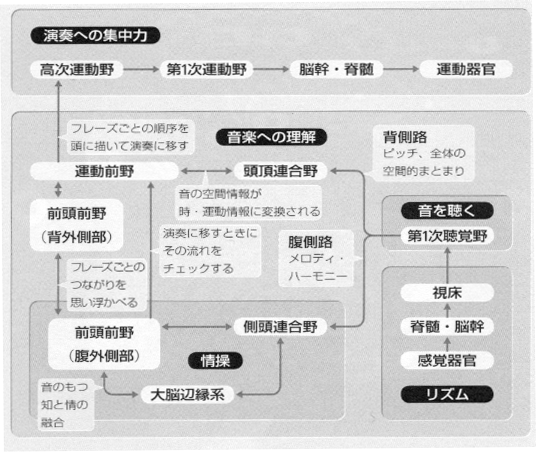
「背側経路」は知覚の空間情報を前頭葉に伝えています。頭頂葉と運動前野のニューロン活動を同時に調べてみると、似たような応答特性をもったニューロンが両域に認められ、視空間、音空間から運動空間への写像変換がなされると考えられます(Goodale と Milner, 1992)。最近、ヒトでPETを用いた研究でも検証され(Andersenら., 1997; Rauschecker, 1998; Bushara ら, 1999; Weeks ら, 1999)、背側経路の伝える空間情報は比較的 「ダイレクト」 に運動のプログラミングに利用されているようです(Wise ら, 1997)。また、短期的に保持されている認知記憶を運動に導くワーキングメモリー(作業記憶)の空間的課題の遂行にも関与しています。
「腹側経路」については、視・聴覚の情報が側頭葉前方域に伝わったのち、前頭前野の腹(外)側部に入力されます。この経路で運ばれる物の形、色彩、音色を含む情動にかかわる情報は、背側経路で運ばれる空間情報とくらべると運動との関係はより間接的に思われます。前頭前野(10野)のニューロンを発火させて、視・聴覚の認知情報を周囲の状況に応じて処理したのちに、運動系に伝達するのです。つまり、感覚野→後連合野→前連合野→高次運動野→第一次運動野という流れです。この高次運動野には、運動前野、補足運動野(内側面6野)などが含まれます(丹治、1999)。この背腹の二つの経路について、さらに(注4)。
個々の音の情報はこのように脳内でさまざまな方法で処理されて、単一の周波数からなる純音・基音から複合音へ、そしてピッチ(音高)、リズム(律動)、和音(ハーモニー)、メロディー(旋律)、さらに音の「複合流体」がもつフレーズやストリーミングなどを統合して私たちは「楽曲」と認識するわけです。(注5)。
ところで、それまでに一回も音楽を聴いた経験のない人が、クラシックのシンフォニーを初めて聴いて楽しめるでしょうか。「クラシックを初めて聴いてすぐに理解できた」という人がいるかもしれませんが、その人はそれまでの人生で、何度もクラシック音楽のいわば「断片」を聞いた経験を持っているはずです。例えばバイオリンの音色や短いメロディーを小さな頃から聞いていたに違いありません。人が成長すると同時に脳も発達します。その過程で、何度も何度も、さまざまな音を、歌を、楽曲を聴き、脳に少しづつ記憶させていきます。幼少の頃は母の子守歌や童謡を聴き、小学生になるとテレビのアニメの主題歌や歌手の歌を憶え、趣味が音楽であればクラシックやロックなどのCDを聞くようになる。このような経験が個人個人の音楽体験となって、その人の脳の中に音楽の記憶と神経回路を形成します。
今のような豊富な音楽媒体の存在しなかった昔には、伝承音楽や民族音楽、日本で言えば民謡が村の人たち皆が歌える音楽でした。他の種類の音楽に触れようもなかったので、皆がほとんど同じ音楽体験を共有していたわけです。現在では、音楽体験は人によって大きく異なっています。一般の音楽好きな人々は歌謡曲やポップスなどの比較的単純な繰り返しの多い曲を好んで聴きますし、音楽の専門家は複雑で高度な楽曲を聴くことが多いでしょう。個人個人の脳は生まれ持った素質(遺伝的要素)の他に、さまざまな後天的な音楽体験によって音楽的に成長していくのです。これがある程度個人の音楽に対する好き嫌いを決定しています。
脳の発生過程には、学習に適した臨界期(critical period)というものが存在します。例えば、生まれたばかりの仔ネコを縦縞ばかり見せて数か月育てると、その後、その目は横縞を感じることができなくなります。後頭葉の視覚野のニューロンが周りの刺激に適応して発達できないからです。人間でも10歳ころまでに外国語を習得すると、現地の人たちと同じような発音や聞き取りを身に付けられます。これも脳の語学能力に臨界期があるからです。音楽について言うと、世間で言う「絶対音感」(音高を厳密に聴き分ける能力)がこれにあたるかと思われます。大脳皮質の聴覚野のニューロンが絶対的な音感を習得できるのに臨界期があるからです。しかし、本当に重要なのは、或る音と他の音との関連性を認知してゆく「相対音感」のほうで、いわば音(楽)によって思想・感情を伝えようとするときに要(かなめ)となるもので、大人になってからでも練習によって身につけられるものです。一般的に、ニューロンの発達は幼い時に活発です。一説には、人間の大脳には100億個以上、脳全体では1000億個におよぶニューロンが存在し、毎日10万個づつ死んでいると言われます。生まれたての赤ん坊には大人の約2倍のニューロンが存在します。その後の成長に伴い、不要なニューロンは淘汰され、必要な神経回路が形成されます。外界からの刺激を受けるとそれぞれの機能に関係する神経回路も強化されます。運動選手が練習に励めば、運動機能に関係する神経回路が強化されますし、音楽の練習を重ねれば音楽に関する脳の領域が発達します。このような神経の可塑性(neural plasticity)は、大人になると低下していきます。しかし、最近流行の脳のトレーニングなどに見られるように、老人になっても刺激を与えることで脳のニューロンを活性化し、老化を遅らせることができるようになります。
最近、聴覚系神経経路に関して、認知と運動をつなぐ運動エネルギー論とその変換モデル論をめぐり、Nature Neuroscience 誌上で興味深い討論が交わされました(Belin と Zatorre、2000 ; Romanski ら, 2000)。ベリンとツアトール(2000) は聴覚性認知の処理過程について、ロマンスキーら(1999)やカースとハケット(1999) が提唱した what/where モデルを大筋で認めた上で、what/how モデルという考えを提示しました。すなわち、ベリンとツアトール(2000)は、腹側経路は音の源が何であるか(what)を同定することに関連するが、背側経路は複数個の音から成る響きが時間とともに経過していくとき、それらが如何なるもの(how)であるかを認知することに関連すると考えました。
そして、この後者(how 経路)はスペクトル運動の正確な分析に基づく「 “where-in-frequency” または ”how”」であるとし、このスペクトル運動は、周波数空間における聴覚性エネルギーの最大値の位置変化に対応すると論じています。この聴覚性のスペクトル運動を周波数の次元に沿った基底膜上の聴覚エネルギー運動と考えると、網膜上の光の運動と考えられる視覚性空間運動エネルギーに同類のものであり、共に感覚上皮に起こるエネルギー変化/変換の分布に対応すると主張しています。これらのモデルには実験による証明が必要でありますが、ベリンとツアトール(2000)は、言語認知においては、腹側経路は話者の同定(what)に、背側経路は聴覚性空間処理過程を通じて言語内容の抽出(how)に関与すると提言しています。それに対して、ロマンスキーら(2000)は上側頭回に位置するcaudal beltという領域は、聴覚性の運動過程におけるスペクトルと空間の両者の要素を結びつけているスイッチボードのような役割をもち、where 経路の起点となっており、ここから能動的話者としてのspeech(話し言葉)の過程が始まると考えると答えています。この音の連続というデジタル信号から言葉の内容というアナログ信号をつかむという脳内でなされる「変換事象」の研究は今後の重要な課題です。なお、腹側経路および背側経路という2つの大きな流れのほかに、聴覚認知の様々な面に対応するいくつかの小さなルートがあり、これらの全体的構成は今後解明されてくるでありましょう(Kawamura と Naito, 1984;Kaas と Hackett, 1999)。[この一節は加筆された]。
興味深いのは、自己を中心とする認知に関わりのある背側経路から一次運動野(4野)への直接投射はないものの、4野と強い結合関係をもつ運動前野(6野)に投射がなされ、対照的に、ゲシュタルトや情動の処理と関わりのある腹側経路からは4野や6野に直接投射はなく、その前段階として前頭前野(10野)に投射がみられる点です。視覚系においても、聴覚系においても、後連合野で受容され、処理された認知情報が、前頭前野で文脈(周囲の状況)に沿って行動学的に意味づけられ、組み換えられて、高次運動系に伝達され、演奏や彫刻といった目的行動がなされるのです。前頭前野は、それらのモジュール情報が能動的発現をするにさい、「仲介変換器」(インターフェイス)として重要な役割を演じています。
歌を歌い、楽器を演奏する時には、まずその曲を覚えなければなりません。そのためには何回も繰り返してその曲を聴き、脳に記憶させるのです。好きな曲を何回も聞いているうちに、知らないうちに細部まで覚えてしまうこともよく経験します。知らない曲でも繰り返し大脳皮質で処理されているうちに、音の属性が記憶され、音楽として認知されるようになります。とくに大脳辺縁系の海馬は短期記憶を形成するのに重要な部位です。「曲を聴いたときには旋律を繰り返せても、次の日には忘れている」のは、短期記憶だからです。海馬で形成された短期記憶のうち、とくに印象的なものは大脳皮質の側頭葉に長期記憶として蓄えられます。東南アジアの老人が子供の頃に憶えた日本の歌を歌えるのも長期記憶の成せる業(わざ)です。ただし、専門的に音楽教育を受けてない普通の人の脳は音楽の理解や処理のスピードが遅く、能力も低いので、歌ったり演奏したりできる曲の数は限られます。自分の好きな曲だけ記憶できるのです。これに対して音楽の専門家の場合には、曲の理解や記憶ははるかにスピーディーに行われます。彫刻家や画家は形象を具象化する前に内的形象のイメージを視空間内に位置づけており、作曲家や演奏家は楽曲の内的響きを聴くことによって、現実の日常的空間とは次元を異にする音空間を創造していると言えましょう。(注6)。
機能的核磁気共鳴映像装置(fMRI)で調べてみますと、演奏家の脳では楽譜をみたときに聴覚系の感覚性言語野が活動しており、記号の流れを意味のある音の流れとしてとらえていると推察されます。実験によれば視覚性の言語野の活動は低いようなので、楽譜を単に視覚的にイメージングするのではなく、一種の文脈として「音を読んでいる」ことになります。
演奏とは、このようにして形成される内的響きを運動エネルギーに転換することです。どのような運動によってどのような表現ができるか、演奏家の記憶にくみこまれた運動パターンから自動的に抽出される動きの組み合わせで、演奏が行われるともいえます。もちろん、自動的運動パターンはあくまで補助的なもので、「悲しみ」の表現も場所、時間、集まった人により変化して当然でしょう。
宮下グループ(Hasegawaら, 1999; Tomita ら, 1999)は、側頭葉に保存された記憶の想起にさいし、前頭葉から側頭葉に指示がなされることを明らかにしました。彼らは図形の連想学習をさせた後に脳梁の後半部を切断して、左右の大脳半球視覚野間の交流を絶ったサルを実験に用いて、図形連想時における側頭葉のニューロンの電気活動を調べた結果、側頭葉内記憶を検索する信号は前頭葉が制御していると結論づけました。聴覚系においても、音楽家は音(符)の順序付けをイメージする回路(連想→作曲→演奏→記憶)を前頭前野により活性化して、側頭葉に貯蔵されている記憶を想起し、内的「響き」と照合しながら練習を行っていると考えられます(川村、2006)。
石を削るミケランジェロが石塊を前にして心の内に削り出される形を予見するとき、「観念形成領域すなわち“言語野”」に形成されている内的イメージと、削り出されつつある実際の彫像形とが、絶えず照合されていくに違いありません。このような内的イメージの外化の過程が芸術における創作でありましょう。このときの形を響きに置換すれば、それは音楽の創造にも演奏にもなります。演奏家は、自分が響かせている音が正しいかどうかを自分の内なる響きとの絶えざる照合によってチェックしつつ、それに基づいて演奏とよばれる運動を修正していきます。作曲家や演奏家は作品という形で具象化しようとする内的響きを、音空間として形象化させ外化させています。専門家は訓練を重ねることによって、音が外的空間に鳴り響いていないときでも、音のイメージを大脳皮質連合野内に連続的に想起し、何らかの意味をもった形象=作品へとまとめあげる技(わざ)を身につけていきます。プロの演奏家はその楽譜から作曲家のイメージと響きのルーツをたどり、これを外的な響きとして再構築していきます。記憶に書き込まれ符号化された連続的な内音の響きを、小脳・大脳関連ループと大脳・基底核・視床関連ループのなかで学習された運動パターンを意識下(随意性でないという意味)で呼び覚ますことによって、身体運動により外化させて演奏を行ないます。聴覚のほかに、視覚、体性(皮膚および粘膜を含む)感覚、内臓感覚などの全感覚が総動員されると、感動が生まれます。
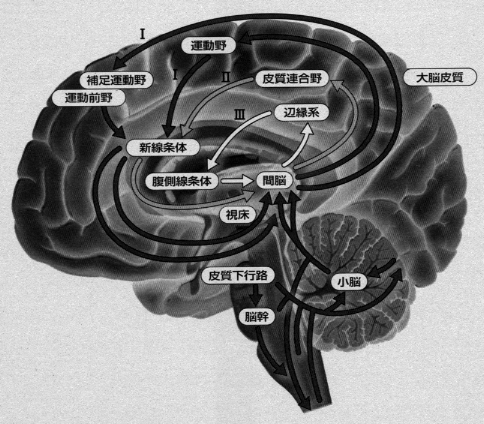
最後に、運動、すなわち、能動的行為についてまとめてみます。大脳皮質の一次運動野(4野)の興奮を脊髄の運動ニューロンに伝え、随意的に筋肉を動かしている神経伝導路に「錐体路系」があります。しかし、運動は「随意的」のものだけではありません。全身の運動を「自動的」に調和させて、円滑に統御する機構があり、それは系統発生的に古い「錐体外路系」といわれるシステムです。運動系の基本的な形態的基盤をなすもので、これには、小脳、大脳基底核、脳幹網様体や脊髄、を中心とした、少なくとも3つの「不随意性」運動に関わる神経回路があります(注7)。
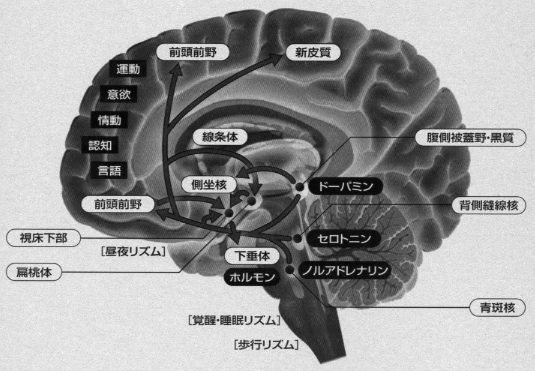
動物は進化の過程で、コミュニケーションや協同作業を通じて、環境へ能動的に働きかけることにより、ブローカの言語野を含む前頭前野を発達させてきました。環境条件を変えられたサルに、新たな解決策を考えださせて、成功したときにのみ報酬を与えるとういうパラダイムを施行させて、サルの脳内活動を分析してみますと、特異的に反応するニューロンが前頭葉内側面の帯状回でみいだされました(Shima と Tanji , 1999)。この部位は前頭前野と互いに連絡する領域で「意欲」の発現に関係する領域と考えられています。なお、帯状回や前頭前野には中脳腹側被蓋野(VTA)からのA10 ドーパミン含有線維が多数終わっていて、ここは意欲、情操、道徳など「高次精神」機能に関連する領域とされています。
能動的ロゴスの座である運動性言語野を含む前頭連合野と、受動的ロゴスの座である感覚性言語野は連合線維によって結びつけられており、各ロゴス野の近傍の前頭葉下部と側頭葉前方部はそれぞれパトスの座である大脳辺縁系(扁桃体や海馬)と投射線維系によって結びついています。このように、連合野と辺縁系とは密接に関連しています (川村、2007b)。こうした領野間のニューロンの働きは、いったん運動性言語中枢近傍の前頭前野でシナプスを形成し、この“仲介域”で「組み替え」を起こしたのちに、運動系皮質域に伝えられます。感覚性の認識が情動と結びつき、直接運動系を働かせるのではなく、前頭前野で情報処理をした後に、運動系を活性化させているのです。情動機能の発現は扁桃体や側坐核からの入力によって、また意欲の衝動は脳幹や帯状回からの入力によって維持されています。とくに、視床下部や中脳からのホルモン、ぺプチド、アミン(セロトニン、アドレナリン、ドーパミン)などの入力もあり、意識・注意集中、呼吸、循環の調和が保たれます。演奏表現は、こうした「広義の運動野」の活動によりなされることが了解されるでしよう。とくに重要な役割を演じているのは、小脳の学習機構とともに、線条体の運動調節統御、それに脳全体としてバランスを保った調和した機能でありましょう (図4)。
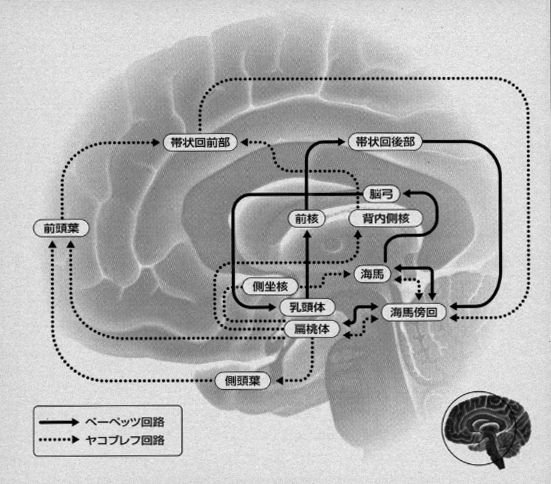
ここで注目したいのは、生物の生命現象は神経回路という一見硬いパターンのみでなされる営みではなく、体内を循環する血液成分、免疫成分、神経伝達物質などをも含む液性の(栄養)因子が大きく関わる全身的な営為であるということです(図5)。ギリシァの昔から、エンペドクレスの「四元素(火、水、空気、土)説」、ヒポクラテス「四液(血液、粘液、黒胆汁、黄胆汁)説」に連なる「四気質(多血質、粘液質、胆汁質、憂鬱質)説」が唱えられてきましたが、人間、とくに芸術家の個性(性格や体質)の形成についてはこれらの体内外からの影響が大きいと思います。
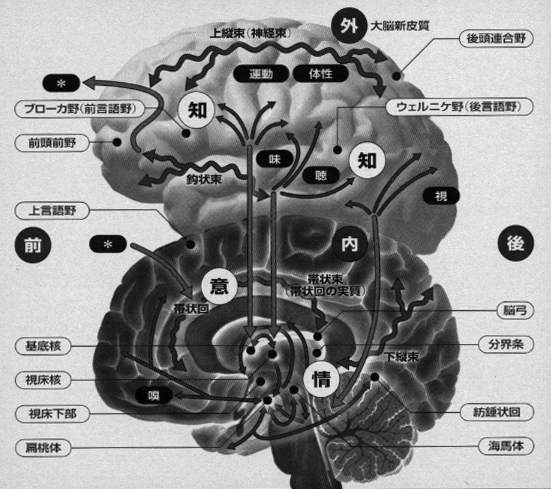
音楽の聴受と演奏において、高次認識系としての言語表現は液性因子が関与する感情表出系と不可分に結びついています(図6、図7)。すなわち、この時には、前頭前野→高次運動野→第一次運動野(一部が後連合野にフィードバック)として形成される神経回路が言語野および情動の回路と結びついて形成されます。歌詞の内容を識って聴く、オペラやミサ曲などやジャズ、ヴォーカルまで含めた種々の声楽曲において、知らずに聞く場合にくらべて情動中枢が強く刺激されることは日常経験するところです。芸術をエンジョイするに当たって、言語中枢(条件反射第二信号系、パブロフ)のはたらきが大きいことをあらためて意識させられます。(注8)(図8)。
昔から、素晴らしい音楽について語る時、しばしば、「神の啓示によって生まれた音楽」とか、「天賦の才能」という表現が使われてきましたが、人が生みだし、演奏する音楽は間違いなくすべて人間の脳(の活動)から生まれたものです。たとえば、モーツァルトの音楽が他の作曲家のものと区別されるのは、彼の脳にそれを特徴づける「音楽回路」 が作られているからに他なりません。このような独自の脳内に形成される回路は、文学や絵画など他の媒体を用いる芸術にも共通するものでしょう。モーツァルトの「音楽回路」は、幼いうちに父レオポルトから規則正しい音楽教育を受けて獲得したその独自性(個性)の上に形成されたものですが、もともと誰にでもある「ヒト固有」の遺伝子から創り上げられた「機能的」な神経回路なのです。ということは彼の幼少期のころに受けた密度の濃い音楽体験が脳内に記憶・蓄積・整理され、テオフィーリウス(神に愛されたもの)の能力として出力されるまでに成熟していったことを物語っています。
音楽は古くギリシァ・ローマ時代の教養人にとって、算術、幾何学、天文学と共にマテーマテカ(学ばれるべきことども、「四科」quadrivium)のひとつでありました。秩序立った世界=宇宙(コスモス)の響きは(数比例による)数学的構造をもち、それは和音という「ドグマ」(ピュタゴラス)の存在ないし発見でした。それに、個々の人間の発声器官はまさに音楽をつくる楽器(instrumentalium)そのもので、呼吸器系統さらに横隔膜・腹筋・骨盤など、そして皮膚・粘膜を含めてからだ全体が外からのすべての感覚刺激に反応します。これに体内を循環する液性因子が加わってはじめて可塑的性質をもったダイナミズムが生まれると言ってよいでしょう。これらを支配する自律神経の中枢が視床下部にあります。
モーツァルトに限らず、ヴェルディも、カザルスも、マリア・カラスも、ゲルギエフも、すべての音楽家、作曲家、演奏家、歌手、指揮者はそれぞれ個性をもっています。音楽家の個性は、よく技芸であると言われますが、そのおおもと(根元)はからだ全体を統御している脳にあります。その意味で「音楽するダイナミクス」は脳の機能と成り立ちに依存していると言えるでしょう。
畏友、川野仁先生(東京都神経研、発生形態部長)と神崎仁先生(慶應義塾大学名誉教授、耳鼻咽喉科)から貴重なご指摘を頂いたことを記して、感謝の意を表します。
注1:聴覚系では末梢から中枢の第一聴覚皮質(A1)まで特定の周波数によく反応する部分が順序よく並んでいます。蝸牛基底膜上の周波数の場所表示はその基部が高周波音、先端部が低周波音に対応しています。蝸牛核や上オリーブ複合体でも、周波数の場所表示が保たれています。下丘の中心核では同一の周波数に同調したニューロンが面状に並び、同心円状に層構造に配列しています。そして低周波音に応ずるニューロンは背側に、高周波音に応ずるニューロンは腹側に位置します。内側膝状体の腹側核は周波数ごとの層構造を示し、そのニューロンは下丘中心核から主たる入力をうけてA1に投射しています。
なお、外側毛帯核の背側核や下丘の中心核にある多くのニューロンには、抑制性神経伝達物質GABAが含まれ、周波数同調を先鋭化する構造となっています。
注2: 連合野について:聴覚野や視覚野や運動野のような特定の機能の中枢ではないと考えられていた領域。現在では脳内の情報処理に重要であることが分かってきました。後(部)連合野と前(部)連合野に分けられます。後連合野は後頭連合野、側頭連合野、頭頂連合野に分かれ、おのおの、視覚、聴覚、体性感覚に関する高次機能を司ります。前連合野は前頭連合野あるいは前頭前野とも言われ、高次の運動をふくむ能動性機能に関与する領域です。
注3: 言語野について:後連合野にある、言葉を理解することに関わっている感覚性言語野(=後言語野、ウェルニッケ野)と、前連合野にある、言葉を話すことに関わっている運動性言語野(前言語野、ブローカ野)が有名です。このほかに、内側面皮質にある上言語野と呼ばれる領域もあり、これらを含めていわゆる「古典的」言語野と言われます。というのは、現在では、言語野は大脳皮質にもっと広い領域的な広がりをもっていると考えられているからです。
注4: 以下は、最近明らかにされた重要な知見です。Matelli と Luppino(2000)によれば、5野からは背側運動前野へ、7野からは前頭前野の内の主溝を含んでその46野へ連合線維が送られます。また、頭頂間溝内のAIP からは腹側運動前野への投射がみられます。そして、独自の形態・機能的な特徴を備えています。すなわち、背側経路には主溝の背側域(9野)へのほかに運動前野(6野) という一次運動野(4野)の前方に隣接した運動関連域に直接投射する成分があります。他方、腹側経路は、前頭前野内の主溝の腹側域(10野および一部の45野と46野) に相当する非運動野に終わります。これら後連合野からの多種のモジュール(module←modus=measure, mode)がそのまま一次運動野に送られてしまうと、これらの流れが個々に備えている役割を一つの全体的機能にまとめ上げて、ゲシュタルトとして全体を認知することがむずかしくなるので、まとまった形での行為およびそれによってもたらされる形象は成立しなくなります。このように高次神経活動の機構をみると、課題ごとに処理する領域を異にするモジュール型になっています。
注5: 楽音の性質は、① 高さ(音高、ピッチ、周波数で表示)、② 強さ(振動エネルギー) ③ 音質/音色(波形)の3要素によって決まるといわれています。 また、西洋では、音楽の3要素として、①リズム(律動)、②ハーモニー(和声)、③メロディー(旋律)がとくに強調されております。
これらいろいろな音の性質の組み合せによって複雑性が生じます。それらの関係を整理してみますと、音の強弱を一定の決まりに従って繰り返したものが拍子(タクト)で、それに音の長短を結びつけたものがリズムです。たとえば、踊りのステップで分かるように、3拍子にもリズムの違いで、メヌエットやワルツやマズルカなどがあります。そして、勇ましい行進曲は決まって2拍子です。さらに音の高さを同時的に組み合わせるとハーモニーとなります。そして、これらの音の性質(強弱、長短、高低)を経時的に組み合わせていくとメロディーとなります。
それでは、これらの脳内機構は聴覚中枢組織のどこにあるのでしょうか?脳機能との関連で考えてみましょう。まず、楽音の3要素は脳幹レベルの機能で、周波数・音波の「物理的」性質が分析されます。ついで音楽の3要素という「心理的」要素が加わる段階になると、さらに高次レベルの小脳や大脳が関わってきます。歩行リズム(音のリズムの発生は歩行や踊りのリズムにあります)やメトリック(metric、リズムの周期的な反復)に関しては、音楽と直接に結びつけた研究ではありませんが、Moriら(2001)やSakaiら(1999) による仕事があります。しかし、メロディーやその背景をつくる和音(トニカ[T]、ドミナント[D]、サブドミナント[S])や、個々の和音が “あたかも言語の文法・構文のように” つながったカデンツ(TDT, TSDT, TST)、さらにこれらを組み合せてフレーズがつくられますが(phrasing)、これら音の流れ(=音楽)の神経科学的研究は音楽家と科学者の将来の共同課題となりましょう。これは大脳皮質レベルの第一次聴覚野(A1)および聴覚連合野における連続的に変化する複数の音の周波数分析と知覚とそれらの融合、および時間・空間的な統合の問題です。音の高低に関する場所表示の違う(すなわち周波数の異なる)A1の領域から、より“高次”の聴覚連合野へ収束と分散のパタンをもって聴覚情報が送られることが知られています(端川、2002)。それから先は、後連合野での認知・認識機能の次、すなわち、背側および腹側の経路を介した情報の伝達の段階、さらには前頭前野の機能が問題となります。つまり、大脳皮質全体のはたらきです。
一般に、知覚においては色々な要素がたがいに関係し、結合し、いくつかの単位に統合されていく機能にグルーピング(grouping)とよばれるものがあります。聴覚系の知覚・認知においても、複数の楽音のグルーピングがあるとき、それらがどの程度時間的に近接し似ていると知覚されるかによって、通常複数のストリーム(stream)が形成されます。そのスタートポイントとなる大脳皮質のA1領域での研究が行われるようになりました(Micheyl ら、2005, 2007; Snyder ら、2006)。
注6: 記憶について。一度利用したらすぐに忘れてしまう「短期記憶」と昔のことをよく覚えている「長期記憶」があります。また、学習や日常経験の思い出、エピソードなどで覚えている「認知記憶」と自転車乗りやピアノ弾きなど体を動かして身につけた、いわゆる“昔とった杵柄”と言われる「手続き記憶」があります。感情と一体になって刻み込まれた「情動記憶」などと呼ばれる記憶もあります。これらの仕組みはそれぞれに違っております。
注7:錐体外路系について。
大脳皮質の運動皮質でいちいち考えて脊髄、筋肉まで命令を下して動かす仕組みが随意運動で、錐体路運動系が関与しております。練習によってそれらの運動はぎこちなさが取れてスムースに滑らかな運動が半ば自動的にやれるようになります。錐体外路運動系といわれる仕組みで、小脳、線状体、黒質、脳幹網様体などにある神経回路(ニューラル・サーキット)が関与しています。図4を参照してください。
注8: トリやサルの脳の発達段階は、ヒトの脳のように言語機能をはたらかせて情動・認知機能を高次化させることはありません。トリの音の認知機能には哺乳類との共通点が見られますが(岡ノ谷、2003)、聴覚情報の処理能力に大きな差があります。われわれは同じメロディをバスやソプラノで歌えますが、このような「移調能力」はトリにはありません。トリのさえずりは、特有の高さに固定されていて、彼らの脳には、たとえば3度低い音程をイメージするというような概念を形成する能力は備わっておりません。音楽における言語中枢の重要性がここにあります。
Andersen, R.A., Snyder, L.H., Bradley, D.C., and Xing, J.
Multimodal representation of space in the posterior parietal cortex and its use in planning movements, Annu. Rev. neurosci. 20 (1997) 303-330.
Belin, P. and Zatorre, R.J.
‘What’, ‘where’ and ‘how’ in auditory cortex, Nature Neurosci. 3 (2000) 965-966.
Bushara, K.O., Weeks, R.A., Ishii, K., Catalan, M.-J., Tian, B., Rauschecker, J.P., and Hallett, M.
Modality-specific frontal and parietal areas for auditory and visual localization in humans, Nature Neurosci. 2 (1999) 759-766.
Goodale, M.A., and Milner, A.D.
Separate visual pathways for perception and action, TINS 15 (1992) 20-25.
Hasegawa, I., Hayashi, T., and Miyashita.,Y.
Memory retrieval under the control of the prefrontal cortex, Ann. Med. 31 (1999) 380-387.
端川勉
霊長類の聴覚野と聴覚関連皮質、神経進歩 46 (2002) 79-90.
He, J., Hashikawa T., Ojima, H., and Kinouchi, Y.
Temporal integration and duration tuning in the dorsal zone of cat auditory cortex, J. Neurosci. 17 (1997) 2615-2625.
Kaas J.H., and Hackett, T.A.
‘What’ and ‘where’ processing in auditory cortex, Nature neurosci. 2 (1999) 1045-1047.
Kawamura, K.
Corticocortical fiber connections of the cat cerebrum. I. The temporal region,II. The parietal region,III. The occipital region, Brain Res. 51 (1973a,b,c ) 1-22,23-40,41-60.
川村光毅
連合野"の線維結合(I)皮質間結合
−−サルとネコの皮質間結合の比較と"連合野"の発達についての試論−−、神経研究の進歩 21(1977)1085-1101.
川村光毅
脳と音楽、臨床精神医学 2001 増刊号 7-16.
川村光毅
認知機能の脳内基盤について ―視覚と聴覚―、 精神医学 44 (2002) 827-837.
川村光毅
脳と精神―生命の響き― pp. 550, 2006 慶應義塾大学出版会
川村光毅 緊張のしくみ、弾くとき脳はどう働く? CHOPIN No.277 (2007a,) 21-26
川村光毅
皮質連合野と小脳の高次精神機能、分子精神医学 7 (2007b) 27-36.
川村光毅
扁桃体の構成と機能、臨床精神医学 36 (2007c) 817-828.
Kawamura, K., and Naito, J.
Corticocortical preojections to the prefrontal cortex in the rhesus monkey investigated with horseradish peroxidase techniques, Neurosci. Res. 1 (1984) 89-103.
Kawamura, K., and Norita, M.
Corticoamygdaloid projections in the rhesus monkey. An HRP study, Iwate med. Ass. 32 (1980) 461-465.
Kawamura, K., and Otani, K.
Corticocortical fiber connections in the cat cerebrum: the frontal region, J. Comp. N
Neurol. 139 (1970) 423-448.
Matteli, M., and Luppino, G.
Parietofrontal circuits: parallel channels for sensory-motor integrations, Adv. Neurol. 84 (2000) 51-61.
Micheyl, C., Carlyon, R.P., Gutschalk, A., Melcher, J.R., Oxenham, A.J., Rauschecker, J.P., Tian, B., and Wilson, F.C.
The role of auditory cortex in the formation of auditory streams, Hearing Res. 229 (2007) 116-131.
Micheyl, C., Tian, B., Carlyon, R.P., and Rauschecker,J.P.
Perceptual organization of tone sequences in the auditory cortex of awake macaques, Neuron 48 (2005) 139-148.
Mori, S., Matsuyama, K., Mori, F., and Nakajima, K.
Supraspinal sites that induce locomotion in the vertebrate central nervous system, Advances in Neurology 87 (2001) 25-40.
岡ノ谷一夫
小鳥の歌からヒトの言葉へ 2003 岩波書店
Rauschecker, J.P.
Cortical processing of complex sounds, Curr. Opin. Neurobiol. 8 (1998) 516-521.
Romanski, L.M., Tian B., Fritz J.B., Mishkin M., Goldman-Rakic P.S., and Rauschecker, J.P.
Dual streams of auditory afferents target multiple domains in the primate prefrontal cortex, Nature Neurosci. 2 (1999) 1131-1136.
Romanski, L.M., Tian, B., Fritz, J.B., Mishkin, M., Goldman-Rakic, P.S., and Rauschecker, J.P.
Reply to ‘what’, ‘where’ and ‘how’ in auditory cortex, Nature Neurosci. 3 (2000) 966.
Sakai, K., Hikosaka, O., Miyauchi, S., Takino, R., Tamada, T., Iwata N.K., and Nielen, M.
Neural repressention of a rhythm depends on its interval ratio, J. Neurosci. 19 (1999a) 10074-10081.
Shima., K., and Tanji., J.
Role of cingulate moter area cells in voluntary movement selection based on reward, Science 282 (1998) 1335-1338.
Snyder, J.S., Alain, C., and Picton, T.W.
Efffects of attention on neuroelectric correlates of auditory stream segregation, J.Cogn.Neurosci. 18 (2006) 1-13.
丹治 順
脳と運動-アクションを実行させる脳 1999 共立出版。.
Tomita, H., Ohbayashi, M., Nakahara, K., Hasegawa, I., and Miyashita, Y.
Top-down signal from prefrontal cortex in executive control of memory retrieval, Nature 401 (1999) 699-703.
Ungerleider, L. G., and Mishkin, M.
In : Analysis of visual behavior, eds by Ingle, D. I., Goodale, M. A., and Mansfield, R. J. W. , 1982, pp.549-586, MIT Press, Cambridge, Massachusetts.
Weeks, R.A., Aziz-Sultan, A., Bushara, K.O., Tian,B., Wessinger, C.M., Dang, N., Rauschecker, J.P., and Hallett, M.
A PET study fo human auditory special processing, Neurosci. Lett. 262 (1999) 155-158.
Wise, S.P., Boussaoud, D., Johnson, P.B., and Caminiti, R.
Premotor and parietal cortex: corticocortical connectivity and combinatorial computations. Annu Rev Neurosci. 20 (1997) 25-42.
出典
編著者名: 小泉英明、
書名: 脳科学と芸術、
版元名: 工作舎
刊行年: 2008年11月3日
工作舎の許可を得て掲載。 一部文体を修正、補筆した。