| その1 | その2 | その3 | その4 | 目次 |
脳と精神は如何に出会うか(その1)
- 1] 序曲/プロローグ(Prologue)―脳と精神
- 2] 音の響きと聴受(音楽傾聴)
- 3] 聴覚と視覚-音楽と言語
- 4] 芸術(とくに音楽)における視聴覚合体論
- 5] 音楽の表現形態(演奏)と能動性皮質(前頭葉)について
- 6] 音と言葉と形象
- 7] Gestalt 認識の生理学
- 8] 情動表現と脳内機構
- 9] リズム/テンポとズレの生理学
- 10] 高次神経活動(=ヒトの精神活動)の要諦と精神医学
- 11] 条件反射と心の問題
- 12] 脳と精神の科学の発展と思春期における自我の形成と情操教育
- 13] エピローグ(Epilogue)
1]序曲/プロローグ(Prologue)―脳と精神
この標題は様々に解釈できる。何が共通のキーワードで、何が脳と精神(=心)の接点であるか。言葉であろうか?それとも情動であろうか?それともGestaltであろうか?これらの会合の仕方はどのようになされ、脳の何処で生起するのであろうか。とにかく冒頭の呼びかけとしてはあまりに遊戯的すぎるからもう少し姿勢を正さねばなるまい。
脳の機能について言えば、魚では脳幹レベル、両生類では中脳レベル、爬虫類では間脳・辺縁系レベル、下等哺乳類では大脳レベルという風に動物が高等になるにつれて、中枢神経系の尾方に宿っていた主要な神経系の作用が次第に頭方に移動することが知られている。神経作用頭端移動の法則Gesetz der kranialen Wanderung von nervösen Funktionen (Herric)と言われるものである。ところで、脳の高次機能は大別して、認知機能と情動機能に分けられる。サカナにもカエルにも認知/認識機構は当然備わっている。高等な動物がより高い認識能力を発揮できるのは、その動物がより高次の機能を発揮できる脳を持っていると言うことである。ヒトが高いレベルの認知機能を発揮できるのは、言葉(パブロフの条件反射第二次信号系)すなわち、信号の信号化を理解できる段階に達した脳を働かせていることに基づくものである。この段階に達しない、より低次元の感覚的認識(認知)は広く動物一般に認められるものである。すなわち、感覚器官を通じて入った外界からの刺激入力が大脳皮質の第一(次)知覚野(V-1、A-1、S-1とか)およびこれとほぼ同一のレベルで反応する発達段階のものが存在する。
同時に、大脳辺縁系、とくに扁桃体を中心とした情動の機構についても、単に無条件反射的に反応するレベル(魚、両生類)から大脳皮質が関与する条件反射的(第一次信号系の)低レベル認知が関与する下級~中等哺乳類レベルのものから、第二次信号系が関与するヒト(およびチンパンジー)に近い高等哺乳類レベルにまでいわゆるヒエラルキーが存在する。すなわち、認知機構同様にあらゆる動物に、当然のことながら、情動機構が備わっており、しかもその動物のもつ脳の発展段階に応じた発現の仕方がある。ここに一般共通性と特殊独自性が同時に存在する。認識機構において、言語機能の有無により決定的な、また、質的な相違がみられることは容易に理解できる。同様に、脳内構成として一般的に新皮質よりも“下層”の構造に位置する辺縁系の働きに大きく依存すると見られている情動機構においても、それが言語機能と結びついたとき、量・質の両面で本質的な差異が生じる。つまり、情動機能は高次の認識機構が基盤/根底にあったときに、芸術や文学の分野において格段に高いレベルの機能が発揮される。逆も亦然りである。
ここで冒頭に掲げた2つの高次機能、すなわち認知機能と情動機能が邂逅する。
ここで、脳研究者が考察できることは何か?ヒトの場合、後連合野の中にarea40、area39が大脳皮質内に形成されている。それぞれ縁上回、角回に相当し、頭頂連合野に属しているが、機能的には感覚性(受動的)言語野(ここの障害で感覚性失語症が現れる)とか、観念性行為構築(ここの障害で失行症、失認症が現れる)とか言われている領野である。統括的にあるいは概括的に全体としてまとまった形を造り上げようとする行為に関係する皮質域と言うことが出来る。即物的に言えば、図や絵画を描くときの全体的バランスの取り方がここに含まれ、視覚的には物体(絵画を含めて)を一個のまとまった形態 Gestalt として認知すること、聴覚的には、単音や複合音、高さの異なる2つ以上の音が同時に響くことによって合成された和音、それも二つ以上の音が一定の、簡潔な比例関係に基づいて和する協和音と、複雑な比例による不協和音、またこの協和・不協和音や旋律(メロディー)などの音が或る一つの音-主音・主和音-を中心に統一的にまとまりを形成している音組織としてのいわゆる調性、さらにはこれにリズムの加わった全体的響きとしての音楽(作品)の認知がこの領野においてなされる。また、言語活動における概念形成という面からみてもこの領野は大切な役割を果たしている。つまり、この領野の全体がうまく働き総合的に調和のとれた思考の路筋(Gedankengang)が施行されるに足るだけの能力が獲得されて初めて、人間社会においてコミュニケーションの内容が正しく伝えられるようになる。このように areas 39, 40 において認知される視覚や聴覚の全体的調和や概念形成、思考伝達までを含めると、広い意味での具象化されたもの、即ちこの感覚性言語野において、例えば音楽(作品)を構成する個々の要素と、それが総含されて形成される全体的形姿との相関的・同時的認知によってもたらされる一個の全体像=音楽(作品)を指して用いられる「形姿」「形態」= Gestalt の意味も了解されよう。Gestalt は決して外的な形や姿をいうのではなく、脳内の諸領野の働きが総合的に収斂されたところに浮かび上がる存在の形姿に他ならず、いわば概念化・形態化の働きを象徴するロゴス(言語・理性)と、存在の形姿の色合いや陰影を指していう情動・パトスの調和/融合と呼んで然るべきものであり、造語を用いればLogopathos/Pathologosの醸成が存在の形姿をとったものとしてGestaltの概念を捉えることが出来よう。
形態学的にいうと、ここ areas 39, 40 は頭頂葉(体性知覚の連合野がある)、後頭葉(視覚連合野がある)、側頭葉(聴覚および視覚連合野がある)が接し合いせめぎ合っている領域である。ここに受動的または知覚性の言語野が存在することは、感覚、知覚、認知、認識の問題を進化論的立場から論じる上で重要な視点である(詳しくは後述)。
この話の流れの中で、最も大事な点は、大脳辺縁系と密接に関連している場所、すなわち、下部(および前部)側頭葉と下部(および眼窩面)前頭葉-この2つの葉の部分が鈎状束という皮質・皮質間線維によって強く結合されていることは留意しておくべきである-と知覚性言語野が上縦束や下縦束という連合線維によって(恐らくpolysynapticな成分が多い)密接に結びつけられているということである。しかもヒトの脳内にはこの結合をもたらした形態的基盤の存在が認められる。このことは、非常に重要な意味をもつ。すなわち、ヒトとしての脳に発達するまでの間に条件反射の第二次信号系、すなわち、言語系条件反射が形成され得る条件が充分に熟していたということである。
以上、われわれは、受動的脳活動(passive brain activity)を担っている後連合領野を中心に論じてきた。ここで、さらに、能動的脳活動(active brain activity)の座である前(頭)連合野(前頭前野、前頭葉)に話の中心を移すと、創造活動と動的表現を含んだ動物(ここでは人間を主として考えている)と環境(人間と自然を含む)との関わり合いの領域に足を踏み入れることになる。
音楽表現について言えば、作曲家として、また、演奏家としての力量を示す芸術性資質が如何に作り出されるかということにもつながる。また、環境に働きかけるヒトの能動的活動性、創造性は何処に生まれるかという問いにも近づくことになる。さらに、精神分裂病患者の能動性欠如、自閉、これの機構の解明への第一歩ともなり得よう。
本稿において、1)音楽の表現形態と運動、2)精神病にみられる自発性障害と感情障害、という能動的な側面をもつ2つの大きな問題について拙速、大胆ではあるが考察してみたい。一部、受動的性格をもつ後連合野の機能に関わる音痴とか両価性の事象も含んで、前頭連合野/前頭葉の機能を中心に脳機能を改めて考察してみたい。
習作として、思索の断片を提示する方法を採用しているので、同類の対象に対して異なった視点から重複して考察することになろう。
私はここで、今日の研究状況からして、なお明解な説明を要求されないで済まされる領域を扱っているので、無理にテキストブック的表現を用いて読者に媚びるようなことはしない積もりである。
2]音の響きと聴受(音楽傾聴)
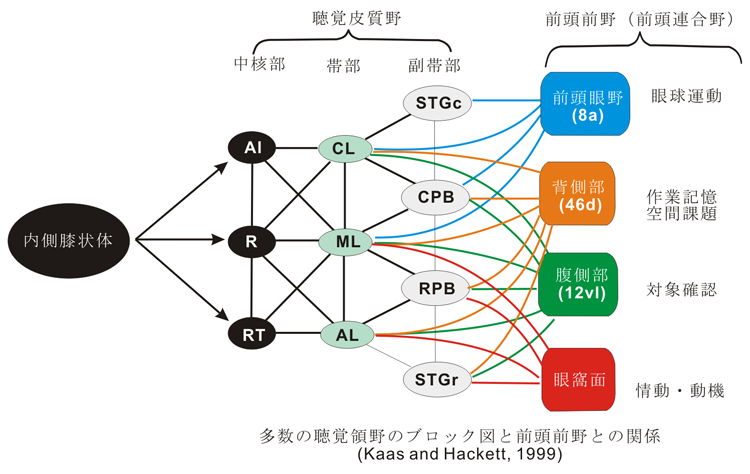
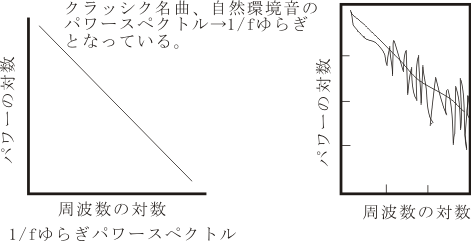
視覚系に比較して、研究が遅れているが、聴覚領も機能領域的に幾つかに区分される。高等哺乳動物の大脳皮質において、視覚系皮質がV-1, V-2, V-3, V-4, TEO, TE, MT, MST, ・・・と分かれているように、聴覚系皮質もA-1,A-2,Ep, あるいは、core(AI, R, RT), Belt(CL, ML, AL), Parabelt(STGc, CPB, RPB, STGr)・・・(cp. Nature, Neuroscience, Dec, ‘99)と区別される。 尤も、ニューロンレベルで実験的に検証するのはこれからで現在は仮説提唱の段階である。最近の研究によれば、丁度視覚系が点の表示と消滅、線分の認識、色の識別、運動方向選択性応答、四角形と三角形の弁別、対象物体の再認、ヒトの顔貌の認知およびその意味づけなど、一種のヒエラルキーを認識の範疇として持っているように、聴覚系にも単音から和音、協和音と不協和音、異なる調性と転調、フォルテとピアノ、それらの連続系としてのメロディーなど各々の質的に異なる性質をもった音の響きを認識する(神経科学の言葉でいえば、それぞれ個性のあるものに分化して特徴抽出された)ニューロン群が特定の皮質部位にまとまって(多くの場合、柱状 columnar に整列して)存在しているため、将来的には機能的、形態的に独立した(特徴のある)幾つかの領野に区分されることになると考えられる。これら多数からなるニューロン群は、皮質皮質間線維により互いに結合されており、この神経回路を活性化させることにより(passiveには聴受により、activeには演奏などにより)機能的にもその結合が強化される。そして、より高次の反応形態を持ったニューロン群に伝えられることにより統合されていく。このような反応の連続性の上に特徴を抽出されて確定された「個」が統合へと導かれていく。これらの過程(プロセス)が後連合野内で行われる。これらの脳内の形態的基盤が存在することにより、単なる音の羅列ではなく、一個の「表現」= Gestalt を形成すべく、意味づけられた連続的な響きとしての音の調べが、演奏者から聴衆に伝達される。その際に或る音表現が聴覚を通じて脳内のどのような領域に働きかけるかは音表現内容や響きの質によって異なる。おそらく、ゆらぎの多い音表現はPathosに働きかけるであろう。これは音楽家がそのように考え、響きを捉えてきたことからも特定し得るであろう (cf.Monteverdi魂のリズム。なお、「ゆらぎ」に関しては後述) 。
このように聴く側の立場として聴覚皮質域の階層性(この順序に従って大脳皮質内の興奮は伝達される)があり、ヒトの脳の後連合野内では聴覚連合野の後背部に隣接して知覚性言語野(受動性-与えられた言語を理解するという意味の-言語野)が存在している。また、この言語領野は視覚性連合野の前方域、体性知覚連合野の腹方域に発達している。このように音楽に関連する領域は脳内で言語機能領域と連合線維によって密接に結びつけられている。こうした観点からみて、ルネッサンス以降の言語研究を背景に、18世紀の、とりわけてドイツで体系化された「語る演奏/歌う演奏」とも言うべきいわゆる音楽の言語化という傾向は脳機能の面からみても、またこの機能による音楽=言語の結びつきの点からしても必然的であった(18世紀/19世紀にみられた言語への傾斜、クロプシュトック、フンボルト、ヘルダー、モーツァルトetc.)。言うまでもなく言語を用いることにより概念化が可能となり、その理解はヒト脳で初めて可能になったもので、パブロフの条件反射第二信号系に属する信号の信号化という脳の高次機能である。当然のことながら、聴受[あるいは音楽の享受(鑑賞)あるいは音楽傾聴]における脳機能の働きは、個人の当該領域の発達(遺伝的または先天的および後天的-教育、訓練、環境要因が大)によって差が生じてくるし、この脳の機能上の差は個人の音楽への理解の差として現れてくる。しかもひとつの言語、ひとつの響き、またその連続性、統一性に、例えば Mozart の作品にみられるように、響きは表面的には簡潔でありながら、その簡潔な響きの中に多層的意味が内在していればいるほど、脳の活動的機能の反映としてこの差は大きくなる。もっとも言語理解の多様性と結びついたこの音楽の多層的内在性のすべてが同時的に聴受されるわけではなく、その中から何等か特定の意味が聴受・認知されるわけであって、このように内的意味の多層性の中から各自によって異なる特定の響きが認知されるのは、これまで述べてきた新皮質レベルの活動に大脳辺縁系に属する古い皮質および終脳内の核集団(扁桃体や側坐核など)及び間脳(とくに視床下部)や中脳ドーパミン系・セロトニン系を含めた皮質下の情動機構の働きの差に起因するものと思われる。Pathologosを根底に持ったアフェクト/脳活動としての心は新皮質から辺縁系や脳幹までも含めて、人間に固有なアイデンティテーを規定する内在的なものであり、音楽の聴受において、ひとつの演奏から聴き手によって異なる響き・意味が認知される。その働きがつまりは各自の存在の特質、アイデンティテーを作り上げるのである。と同時に、こうした聴受・認知を構造的に捉えるなら、ほぼ次のようにそれを要約することができるであろう。
近代音楽、つまりルネサンス以降のヨーロッパ音楽の中で応用された、いわば共感・共鳴の考え方によって音楽が何等かの表現を伝え得るとされた。音楽にはその本質的働きとして、いわゆる“いやし”の効果が求められたが、それはconcentusつまり演奏者の魂と聴き手の魂とが共感し共鳴して同一の動きをとることによって果たされるものであるとされた。ルネサンス期イタリアの哲学者フィチーノを主たる源泉とするこの考え方によれば、人間の喜怒哀楽の情は音の動きによって聴き手の魂がゆり動かされることによって生起するものであり、そのためには演奏者自らがその魂の働きを音に託すべく技を身につけなければならず、そのような技によって作り出された音の働きが演奏者と聴き手の魂を同一の情調へと共感・共鳴させるわけで、ここにaffectus=アフェクト論の根幹をみることが出来る。ここでアフェクト、あるいは魂の動き、とされたのが要するに後連合野の働きになるものであると言い得るであろう。やがて17・18世紀への進展の中で音楽もその一面において概念化の傾向を強め、例えば特定のイメージ・内容を文学のように伝達すべき標題音楽や描写音楽も作り出された。また、それと併行して特にオペラのような声楽のジャンルで、歌われるべきテキスト・言葉に内在する情調を、歌声を伴奏する楽器が表現し、さらには声楽曲におけるこのような器楽の表現法が独立して器楽曲の中で用いられるようになることによって、例えばモーツァルトのオペラに聴かれるように、歌=言葉と器楽=情調が深く共鳴しつつ形成される響きの宇宙もまた可能にされたのである。この場合の歌=言葉は概念を、器楽=情調は上のアフェクトを各々に担うものであり、両者が不可分の関係に置かれているのが理解される。或る情調が特定の気分を結果させ、その気分が特定の表現、すなわち言語による規定を結果させてアフェクトの概念化が成立する。あわせて、こうして成立した概念=言語が再びアフェクトに働きかけてその陰影を逆に深めて、しばしば概念化された言葉=テキストによっては容易に表現し難い、いわば高次化された情調を音楽が響かせることになる。宗教音楽における「神」や「神聖」といった言葉と、それと結びついて響かせられる崇高な響きによるconcentusはその典型な事例である。これも、後連合野から前頭前野への脳機能のはたらきによって可能とされる情動→言語(概念化)のプロセスに一致する、芸術創造における高次化の現象である(なお詳しくは付論を参照)。
3]聴覚と視覚-音楽と言語
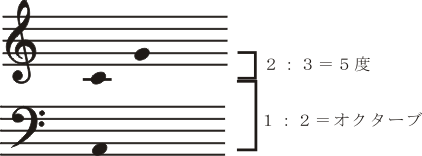
物体から生ずる空気の振動は内耳の有毛感覚細胞で電気的刺激に変換され、その興奮は延髄・橋・中脳・間脳内を順次ニューロンを替えて聴覚伝導路を上行し第一聴覚皮質領野(第一次聴覚野)に達し、ここで最初に音の性質が分析(感覚/知覚)される。その振動数 / 周波数に定められた特定の高さの音が知覚される。つまり、アナログの性質がデジタル化される。そしてその興奮は聴覚連合皮質内を段階的(stepwise)に、より「高次」化へと向かって進行する。ここで、このようにデジタル化されたものは再び高次のアナログを持つ性質に再構築される。上記のように音楽は単に音(響)の無秩序な集積ではない。旋律(メロディー)があり、和音があり、音の強弱(振幅の大小)や長短、すなわち、リズムがある。それぞれ複数の音の組織するパタンをもち、音のモジュールをもち、時間的構造をもっている。その和音(DO, MI, SOなど)は3つの周波数が調和している。調和しているとはその周波数が一定の比例関係になって共鳴していることである。これはギリシャ哲学および自然科学、とくにピュタゴラス(派)において、彼らに先立つ宇宙論との関連の中で知覚されて以来論じられてきている音と宇宙(コスモス)との間に存在する数的比例関係と秩序の対応性を物語るものである。ピュタゴラスの数的比例関係は音楽においてどのように、どこに現れるのだろうか?。ここちよい(angenehm)音,不快な(unangenehm)音とは何であろうか?語義的にはan-nehmbar「(自己へと来るものを)受容し得ること」で、そのようなものとして認知されるものを言う。これは聴覚連合野の機能に情動affectusが生じることにより高次の脳機能が発揮された状態である。また、同時に響かせて2音間の和音的音程において、感覚的に平穏で調和して響く協和音(程)、と音と音とがぶつかって反発しあって響く不協和音(程)、と言われるものがある。音の振動数に応じた知覚は空気、骨、内リンパ液を順次経た物質の振動がコルチ器官内の感覚細胞(有毛細胞)を最初に刺激し、脳内では、その興奮が蝸牛核、外側毛帯核、下丘、内側膝状体核とシナプスを代えて中枢神経系内を伝わり大脳聴覚領皮質areas 41、42(第一聴覚領first auditory area)で知覚される。これは丁度視野の特定部位が第一視覚領皮質(first visual area, area 17)の一定部位に対応した点として知覚される機構と同様である。
このように聴覚領皮質のcore部分(AI; auditory core area, R; rostral core, RT; rostrotemporal core)は、各振動数に対応した個々の単音を受容(receive)し、Romanski, Tian, Fritz, Goldman-Rakic and Rauschecker (1999, Dec. nature neuroscience)の最近の論文によれば、かなりの推論ではあるが、coreからmedial & lateral belts(CL, ML, AL)に興奮が伝わり、更にlateral beltからparabeltへ(STSwallを含め得る)とauditory pathwayが進行する。しかし、諸要素が融合したところにはじめて成立する音楽に対して、単純音から複雑音、和音、協和音、不協和音、メロディー、調性etc.へと単に階層的hierarchicに内容が複雑化すると解釈することには無理がある。聴覚領内における様々に分化したニューロン群の総合的相互干渉の結果として、皮質ニューロン構成が形態的に発展し、認識能力もこれと並行して進展してきたように思われる。受容(聴受)の場合は、おそらく音楽と呼ばれる総合的響き(和音+メロディー+リズムなど)の全体が脳に働きかけて情動 affectus を呼びさますと考えられる。(脳の瞬間的分析力にはどの程度のものがあるかは「聴音」(ソルフェージュ)の側からも検討してみる必要があろう)。この点、線分、曲線、単純な形、複雑な形象、顔貌(表情)、動き、色、配色、二次元から三次元への形象認知の発展など視覚系における階層的領域区分の機能的発展を見るが如くである。そして絵画や彫刻や陶芸などの場合、これまた、affectus(情)や美を結果させる。以上のように視覚芸術と聴覚芸術との類似性や対比を試みることは不自然ではあるまい。
最近聴覚皮質野の仕事は注目されるようになってきたが、視覚皮質野の仕事がHubel&Wieselの60年代からの詳細な仕事に比べて、Merzenich and Bruggeがmulti-unitのfield potentialを最初にauditory cortexで記録したのが1978年であるのをみても、10年以上も遅れていることになる。ついでに言えば、Mountcastleらの体性感覚野における研究も視覚野程には進んではいないが、auditory cortexのrecordingの仕事に比べれば可成り早くから施行されている。
このようにvisual, somatosensory, auditoryの皮質域内で細かく区分された低次から高次へと段階的に進行する機能域の存在が明らかになるにつれて、この3つの機能領野に囲まれ、そこに刺激/興奮の伝達が集中して終わる所にpolymodal(多種感覚様態)に反応するニューロンが(STS域を含んで、とくにそのcaudal partに)存在することが証明されていることは大きな意義がある。進化論的立場から見て、ここは知覚性言語領野(Wernicke)が存在するところで、この3つの領野のGestalt構成を動的に統括的に総合的にみてみると、この後連合領内に存在する感覚性言語領野内のニューロンは種々の感覚様態に反応する性質を備えており、高次に信号化された情報を処理する領野と考えられる。
音(楽)の比例関係をもった数的秩序や、遠近法画法にみる比例関係をもった空間的距離の尺度を視覚的形態的秩序として直覚することが出来るための脳内領域は視覚性および聴覚性の皮質連合野(auditory cortexでは、belt, parabelt, area22, visual cortexではparastriate cortex、area37,TE, TG野)に対応する。
AIからbeltへ軸索が伸びているが、音(楽)との関連で言えば、幾つかの音が混在するarea22では、雑音(white noise)に反応するニューロンが存在することが知られている。ネコにおいても聴覚皮質AI域に種々の性質をもったニューロンが存在することが知られている(Ju-Fangら,1999)。すなわち、音を聞かせたときの反応性をみて、tuningの幅がシャープなものから長いものまである。また潜伏時間(音を聞いてから反応するまでの時間)にも長短があり様々である。tuningの幅が広いもの、潜伏時間(latency)が長いものはAI域の背方域(DZ)に多くみられ、AII域には更にこのような特徴をもったニューロンが多数みられるようである。また、サルの実験で、white noiseに反応するneuronsはAI(primary auditory)には認められない。また、日本ザルの実験(当分confidential, from Tom)では同種のサルの(クウーという)鳴き声を聞かせたときAIでは多くのニューロンが反応する。そのテープを逆方向に回転させてサルにとって不思議な意味のない音声として聞かせるとarea22(belt, parabelt)の中央部から後部域にあるneuronsが反応する。聞いている音の性状を知ろうと努力して解析中なのであろうか。もとに戻して仲間のサルにここちよい鳴き声を聞かせると、area22の前方域で(TG dorsal)扁桃体と強い結合をもつ皮質域に多くのneuronsが反応する。音の快・不快を判断している状態なのであろうか。
次に重要な視点として言語、音楽の発生、とくに音楽の言語化について考えてみよう。ダーウィン以降の科学が明らかにしているように、サル→チンパンジー→猿人(アオストラロピテクス)→原人(ピテカントロプス)→旧人(ネアンデルタール)→現代人(クロマニヨン)へと発展してヒトへの進化を成し遂げた。サルの社会はボスを頂点とした、家父長的支配社会である。故に、それを引きついだ形で成立したヒトの原始社会を考えてみることはさほど不自然なことではないであろう。ヒト社会に移行する前のサル社会のコミュニケーションはどのようにして成立していたのか。その社会では、相手を攻めたり、あるいは、敵から身を守るような時には、単純音の不連続的な合図、指示、命令という信号としての原始言語(ことば)の形態であったと思われる。ここでは、集団社会を維持するという、いわゆる仲間に対する呼びかけとして、音の素材が生活と一体となって形成されてきた。ある音程をもった呼びかけが有効であったことは容易に想像できる。このように、語感、情感、感情が付帯され、リズムの響きが生じた。この原始(的)社会において、コミュニケーションの手段として、共通の合図、合意となる信号(言葉)が形成され、そして使用された。このようにみてくると、言葉の発生は、認識論的な理性の結果として生まれたと結論づけるよりは、むしろ、太鼓やラッパや笛を合図としてかき鳴らす情動的な背景と結びついて生まれてきたコミュニケーションの手段と考えた方が自然である。その背景は何かと考えてみると、ボスの側からみると集団を支配して他の集団に対抗する必要がある。そのためにはmind control(マインドコントロール)をする必要があるので仲間の交流(コミュニケーション)の一部が支配者と非支配者という上下関係に組織上の必要性から変貌した。快く聴覚系をなごませる音というよりは、むしろ、太鼓やラッパの音は皮膚感覚に訴える臨場感あふれる刺激的な音である。(突飛であるが、全身で刺激に反応するゾウリムシ-原生動物-の反応を思わせる)。その状況下で巫女、シャーマン様の抑揚ある独特なリズム音と踊りを伴う支配性や宗教性への利用が生まれた。最初期の人類の社会はこのようなサルの社会(まだ搾取社会にはなっていない)を引き継いだボス言語とリズム音楽が結合したものであったように思われる。これは、やがて発展する自由人と奴隷との区別が生じた奴隷社会であるエジプトやギリシャの社会にも原形として映しだされる。縄文時代、弥生時代のわれわれの祖先の集団社会も、同様な発展段階にあったと思われる。支配階級にみられるこのような音楽と言語の結びつきとは対照的に被支配階級にあたる一般庶民社会はどうであったか。上から与えられたものの他に、民衆自身のactiveな言語の音楽化ないし音楽の言語への融合現象をもっていた。それは祭りの時に歌われる労働の歌であり、かけごえの集団歌であり、収穫の歌に発展する原形であった。それらは民衆のコミュニケーションの場に、農地、魚池を中心とする地方の民謡として残っている。現在、遺跡が発掘されて猿人、原人の群集生活があきらかにされつつあるが、原型(Urgestalt)をみようとするとこうなるように思う。むしろ、これらの民衆音楽の中に言葉と情緒とが結びついた直接的発展としての音の響きやリズムを感じとることが出来る。
このように生物学的且つ社会学的にその発達過程から見てみると、音楽は1つの概念化した言語として捉えることが出来よう。そして、上にみたように、言語と音楽との間には同時性と階級制が存在する。たとえば、黒人社会が米国をはじめ先進諸国にあたえたspiritとしての黒人霊歌。ジャズ、ルンバ、タンゴの民衆性。開放された民衆のspiritがもつシャンソン。また、北欧のカレワラ民謡伝承歌、ハンガリーのジプシー民衆歌、ロシアのバラライカ、ブラジルのサンバの歌と踊り、大和の古代歌謡やアイヌ民族のユーカラの調べなどの神話や伝説の色彩の濃い原始民族時代の声楽(ついで古代楽器)の発達はこのようにして、発展してきたように思われる。これらはこのように民衆の間から生活を基盤にして、発達してきたものである。なお現在、古典音楽として親しまれているシベリウスやリストやグリークの独特な調べは、このような民族音楽の背景があって、その上に余暇をもつことの許された知識階級によって様式化され、楽理として理論化され、リズム、音階、和音などの“きまり”をもった(約束ごとをもった)文芸として発達したものと考えられよう。
それにしても、ピュタゴラス以降の音の数的概念で説明される自然界の秩序、数的比例を包含する音楽の聴受は、脳の構造や機能と如何なる関わりを持っているのであろうか。元来、音楽と言語、聴覚と視覚が深い関連を持つものであることは、現時点におけるヒトの脳の機能の解析によってかなりの程度にまで判明されるばかりでなく、そのような機能によって人間が作り出して来た伝達手段や芸術作品の歴史によっても裏付けられる。特に芸術の歴史にこのような関連性が如実に現れているのは、芸術作品そのものが基本的には多要素から成るものであり、かつまた脳の多機能の駆使によってもたらされるものであるからである。つまり、種々の芸術の源泉のひとつである古典古代、古代ギリシアにおいて、芸術・哲学・数学といったものがひとつの共通の問から派生して来たものであることが先ずここに問われている音楽と言語や聴覚と視覚の関連性を物語るといわねばならないのである。今日では音楽と言語、音楽と文学あるいは芸術と哲学と数学といったものは各々に独自の分野を形成して相関性を稀薄にしているが、本来これらのものは、広い意味で存在への問、在ることは何かに関わって成立・発展したものである。存在=在ることへの問は一方では宇宙・自然の科学的解明に向かい、また他方ではそれこそ存在論的に、形而上学的解明に向いながら、しかし両者共に在ることの根源を成す空間性に根ざして共鳴の輪を広げたに過ぎない。例えばプラトンの『ティマイオス』において展開された宇宙論は数学的であると同時に音楽の比例論に直接し、さらに音楽の比例論は詩、文芸、とりわけてギリシア悲劇に代表される韻律における言語の響きの数比例に不可分である。また『ティマイオス』の宇宙論は同じプラトンの『パイドロス』における魂の問題から眺められた宇宙の形而上的構造論や『法律』の多分に神話的・音楽的宇宙論と切り離されては存在し得ないのである。
音楽と言語が共に響きによるものである上、響きという、それ自体が空間を形成して在るものであることによって視覚の依拠する空間的機能と共通して「空間」を共有することは明らかである。但し、残念ながら、視覚的空間性=芸術に関して遺された古代ギリシアの文献は皆無に等しく、また音楽も実作品の伝承はこれも無きに等しいこと、あるいは、ギリシア劇における韻律の実際の響きがどのようであったかが不明である点等からして、古代ギリシアにおける音楽・視覚空間の関連性を捉えることはきわめて困難であり、類推の域を出ない程度において捉え得るに過ぎないと考えられる。そのような点からすれば、ギリシア・ローマの精神文化の遺産を継承し、これにキリスト教の考え方を加味してまとめられたヨーロッパの芸術の中に、それもルネサンス以降の芸術の中に、ギリシアにおける「在ること」に関わった芸術・学問の再生を、仮にそれがキリスト教的思考による再生ではあれ、認めることが出来る。例えば音楽のリズムを支えた言語のリズム・韻律は、西洋の詩の一源流であるギリシャの詩のもつリズムの種類すなわち、長短(長短の比率は1:1/2)の組合せ, iambosイアンボス 短長 v - ;trochaiosトロカイオス 長短- v;anapaistos アナパイストス 短短長v v - ;daktylosダクチュロス 長短長- v v;が詩脚として用いられた。ギリシャ・ラテンの詩はこの数的比例をベースとしたリズムをもつものとして表現されて来た。俗ラテン語(フランス語、イタリア語、スペイン語など)化され、もしくはドイツ語のような近代諸語の使用によって言葉のもつ長短リズムの規準は緩くなったものの、現代詩に至るまで(英語の詩も含めて)このrule/規準が生きている。漢詩も韻を踏み平仄の音律をもっているが、洋の東西を問わず、様式化された詩は一定の法則とリズムをもっている。そして、声楽の発展は器楽の発展に先んじており、原始時代の日本でも歌謡が中心であったと思われる(古事記、日本書紀に歌詞とその歌われた状況などの記載がある)。わが国の原始・古代の楽器にはコト(琴)、フエ(笛)、ツヅミ(鼓)、銅鐸などが知られているが、コトを弾く男子の埴輪像も発掘されている。また、一般民衆は石や桶をたたいて音やリズムを鳴らした。母音を長く引く歌い方(琴歌譜)が次第に五五調に、ついで五七調に歌謡の詞型が整理され、やがて五七五七七という短歌のリズムが確立し初期万葉集時代に受けつがれていった(但し、施頭歌、東歌をみよ)。この5世紀から8世紀にかけて大陸、特に百済(くだら)から音楽家も来日した。そしてインドに起源をもつ宗教(仏教)音楽(経典音楽)も伝来し、声の歌(音声言語による)から文字の歌(文字言語による)への転換/変革、または声(聴覚系)と文字(視覚系)との両面からの文化/文学の確立の第一歩という意義がある。音声言語を話すときは人は感情を表し、文字言語を用いて書くときには概念(観念)を表す(ルソー)という。これはものの程度を示す観点ではあるが、正しい面を含んでいる。そして後者の文学は遅れて特定の少数者から始められたことは心に留めておくべきことかと思われる。
西洋におけるルネサンス以降の近代芸術ではギリシャ悲劇の研究が重要な役割を果たし、上流知識階級の学問体系として、悲劇、すなわち、韻文による詩芸術の韻律から新しいタイプの音楽を出発させ、言語の抑揚が、ひいては音楽の抑揚、すなわち、メロディーの構造を規定するようになった。この関係が各国の音楽を支え、例えばドイツでは、やがて18世紀に、ルネサンス以来のドイツ詩文芸と結びつくことによって、バッハの宗教音楽にこうした詩=ドイツ語と音楽の融合のひとつの頂点を見出し、さらに18世紀の後半には、改めてドイツで行われたギリシャ悲劇研究を端初としてKlopstockらによって韻律法が発展させられ、ドイツの声楽曲の隆盛の一要因をなした。そして、モーツァルトやシューベルトの歌曲、ベートーヴェンの第九交響曲第4楽章のFreude合唱等を成立させた。また、19世紀におけるギリシャ悲劇の新たな音楽劇化であるワグナーの舞台作品、たとえば、「トリスタンとイゾルデ」などにみるように、音と言葉(声)と所作を一体化させた演劇的音楽の総合芸術であるオペラの分野で大きな発展をみせた。この流れは、Monteverdiを一頂点とする初期オペラを別にすれば、18世紀ドイツにおける、たとえばMozart から19世紀になってWeberから先記のWagnerに至るオペラの中にその成果をみせた。Verdi, Pucciniに代表されるイタリアのオペラも同様の流れの中で捉えられる。
ここで立ち戻って考えてみる。音楽の作品は単なる音の集合を越えた「何か」であり、音を組み合わせることによってその「何か」を伝えるべく作品を構築してゆくのが作曲であり、そこで構築された「何か」を響きとして具体的に聴衆に伝えるのが演奏である。演奏は上記の言語系の流れ・歴史の中で修辞法と結びつき、とりわけドイツの18世紀にその「何か」の目的とするaffectusの刺激を旨とする修辞的演奏法が起こった。この修辞的演奏法は、音楽におけるロゴスとパトスの関係について考える上でとくに重要な役割を果たす。また、その時、伝えるべき作品の「何か」によって形成される、形而上的時間が働いて演奏の種々の時間=テンポを規定する。これは演奏時の時間の流れ方の問題であるが、やがてこの音楽と言う存在の時間はallegro, andante, prestoなどの用語によって表象されることになった。この時間の表象化、すなわち言語化も、既述の言語=音楽の歴史の中で成立したものである。さらに音楽の「何か」は、演奏の具体的響きが形成する空間の形をとって聴覚に伝えられるが、この空間の形成には調性(ハ長調とかロ短調など)も重要な役割を果たす。空間の概念は本来視覚に関わるものであるが、上記のように音楽も各音各々の固有の響きや種々の調性の結合・組み合わせによる空間(いわゆる音空間)の聴受によって成立する。これはおそらく、脳において、聴覚系と視覚系が相互滲透することによって結果される、聴覚の視覚化によって可能になるものである。
ここで想起されるのは、視覚入力を聴覚皮質で受容するというごく最近発表された以下の実験である(Nature, Vol. 404, 20 April, 2000)。米国MITのSurらのグループは、生後1日のフェレット(ケナガイタチ)の視神経を両側性に実験手術して、聴覚伝導路の間脳における中継核である内側膝状体(MGB)につなぎ変えた。そして成熟動物にまで育ててから調べた。すると視覚情報を受けて育った結果、この動物の聴覚皮質はorientation column(特定の傾きをもった線分に特異的に反応するニューロン群が柱状に配列する)という視覚領に特徴的な「傾き地図」という構造を作るという視覚野に似た発生を遂げていたことが明らかになった。そしてこの領域のニューロンは、音には反応せず光に反応するようになっていた。このことは、遺伝的(DNA)レベルで聴覚情報を処理するように発達するように運命づけられていた皮質が、視覚情報を処理するように人為的に変えられた環境下で養育されると、構造的にも機能的にも新しい環境に適応(adapt)するようになったことを示している。このフェレットの脳は、本来“聞く”場所で“見て”いたのである。
アリストテレスは、ドラマにおいては、演ずべき役柄への、演者による共鳴conpassioは必要であるという。なぜなら、役者の“おそらくaffectusの働かせ方如何によって”作り出される演ずべき役柄とのconpassioの状態が役の仕上がりを左右するからである。ただし、演奏家の場合にはこのようなconpassioがaffectusそのものによるものであるかどうか疑問である。何故なら、演奏は一種の修辞・弁論術であり、ここでは、演奏者そのもののaffectusの表出は余り問われず、むしろ聴き手のaffectusを動かす方法そのものが問題となるからである。
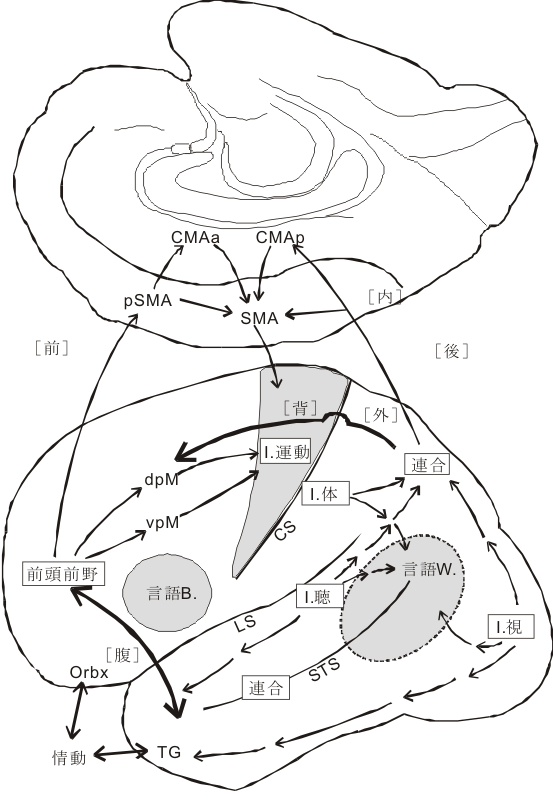
ところで、高度な信号である楽譜を読んで楽器を奏でて聴者のaffectusを刺激すべく連続的に音を響かせていくという知的過程は、単に視覚性符合認知と聴覚性音声変化とを結びつけることにとどまらない。そのほかに、頭頂葉の空間認知、側頭葉前方部(TG野)の情動の受動的認識、前頭葉の時間軸に沿った計量的認知、情動の能動的認識と演奏者の全身的 [筋肉的-cp.セイチェノフの用語。パブロフが師としたセーチェノフ(ベルナールに学んだ19世紀後半の生理学者)は、感覚受容から筋肉活動の間におこる脳内プロセスを彼の反射理論の中で“筋肉的局面”と呼んだ。且つ彼は情動を強化された”筋肉的反応”という言葉で説明した。] 情動表現に支えられて脳は全体として反応している状態であろう。
ここに、ある種の連続音をサルに聞かせて調べた最近の実験がある(未発表、personal communication)。その時はprimary auditory cortex (AI)で多くのニューロンが反応する。しかし、雑音に対してはAIのニューロンは反応せずarea22 (belt, parabelt) のニューロンが反応する。この時この連続音を途中で数秒間切り落して、その間だけ無音に細工したテープをサルに聞かせたとき、サルの聴覚領を調べてみると、AI neuronsの反応をみると数秒間断絶しているが、area22では雑音white noiseに反応するものの電気的興奮性は途切れてないと言う。この「雑音」のもつ意味、音楽を聞くときの意味をどのように解釈したらよいか。われわれ人間が音を断絶した避音室で、雑音のない状態下でヘッドホンを用いてCDを聴く場合と、自然の雑音が入る普通の部屋でCDを聴く状態(音楽会場での聴取はかなり良い条件であるが)とを比較してみよう。発音源(スピーカー)や楽器から発せられた個々の音は、その間に距離がある状態のとき、空気中で互いに共鳴し合うと考えられる。従って、離れて聴受した場合の方が、耳許のヘッドホンで聴く場合よりも、はるかに「調和」した響きを聴いていることになる。後者の状況におけるこの「調和」が、前者で聴受する以上の「何か」をもたらし得ると考えられる。もともと自然界に生存する生物の聴覚は、川のせせらぎや風のそよぎや蝉の鳴き声などの自然音になごんでおり、そこから発生した土俗的なものから作り出されており、ヒトにおいても高尚な道具-楽器-や理論の上に立った楽音を基にした均整のとれた“人工的音”が発音源に近く聴かれるより生な状態や、より器械的なヘッドホンのような場合に共鳴し難くなる。CDの音(楽)をヘッドホンで聞くいわゆる完成音(楽)では発現し難くなってしまっている、自然環境内に存在する“雑音”中に内在する調和比が音を伝達する媒体である空気を共鳴させてふくらんだ響きを作り出すことが出来ると考える。
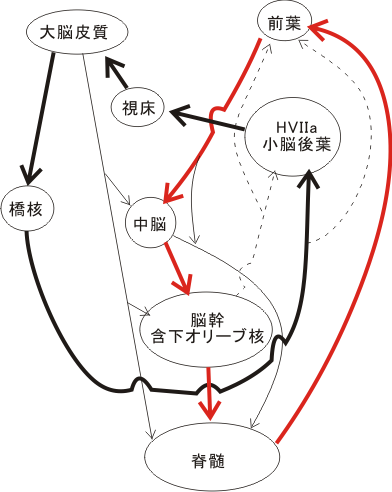
視覚系と聴覚系は中枢神経系内の種々の領域で重なり(overlap)ないし隣接(adjoin)している。1940~50年代(Snider, Stowell, Eldred; '44, '51)から皮質視覚領and/or皮質聴覚領を電気刺激したとき小脳虫部の中央部(虫部葉VIIAと虫部隆起VIIB)からteleceptive impulse(化学物質が直接の受容体に接触して刺激する体性感覚や味覚や嗅覚に対して、視覚や聴覚のように媒体を介して間接的に受容体に働くインパルスをいう)の(電気的)反応が獲得されることが知られている(Sniderらによりネコ、サルで)。これらの反応は小脳の室頂核の尾部でシナプスを変えて中脳視蓋部に伝えられる。(上丘の深部中間部と、そして聴覚系は主に下丘のexternal and peripheral nucleiへ)。これは小脳が関与するいわゆるvisuomotor(視覚性運動), audiomotor(聴覚性運動)の反応で動く視点や動く音源の同定に関連するシステムである。視聴覚系が動く視点や音源の同定に協同して作働している可能性がある(後述)。野生動物は生命維持(攻撃や逃避)のためにも、視覚的にも聴覚的にも動く対象に対して感覚・動作が発達しているものである。
他方、認識(recognition)系では、ヒト、サルにおいてもネコにおいても明瞭に区別された大脳皮質領域で視床からの各visual and auditoryの入力を受け二次、三次のニューロンへ伝達が進行したとき、深い脳溝(ネコではmidde suprasylvian sulcus, 中S上溝、MSS;サルではsuperior temporal sulcus, 上側頭溝、STS)で境されてその両壁皮質域に反応するニューロンが存在する。また、ネコのMSS後部域やサルのSTSのほとんど全域には、多種のsensationに反応するpolysensoric neurons(多種感覚反応性ニューロン)がみられる。更に、サルではSTS域を含めて後連合野から前頭連合野にbroadly localizedの投射があり、前頭前野の弓状溝(arcuate sulcus)の前方域の前頭眼野の前方に隣接して、主溝(principal sulcus)の周囲域(とくに背方域)に可成り広い範囲に視覚性の空間的位置の認定に関わるニューロンが認められる。これらの視覚性位置反応ニューロンの周辺には聴覚性位置反応(静止した音源の同定と思われる)するニューロンが同定されている(鈴木寿夫グループ)。因みに、光源、音源というものは、元来動的なものであり、後連合野で知覚される働きの対象を認知する機能とは性質の異なる、動く形や音の方向性を含めた同定の働きは脳幹、小脳レベルにあり、この問題は後に考察される。
これに連関して、たとえば、動く棒を見た時ある一定方向にものが動いた時に反応するが、軌跡は同じでも反対方向に棒が動いた時には全く反応しない視覚性ニューロンが視覚連合野に存在する。聴覚領内でも音が一定の周波数をもつ高音から一定の周波数の低音というベクトル方向が定まった連続音を聞いたときには反応するが、反対ベクトル方向の変化連続音に対しては全く反応しないという性質のニューロンがbelt, parabelt域に発見されている。
ところでV-4は(visualの)color responsive neuronが発見されている領域であるが、それに相応するauditory natureのニューロンとして何んなものが考えられるか。音の色彩というのは響きの彩りであろうか、などsemantisches Spielをしている場合ではないのだが、存在が認定されれば、「響き」と言う鍵概念に迫れるものとして注目されてくるに違いない。
次にinferotemporal(下部側頭回)の前方域を研究しているSaleemらのデータを考えてみる(一部未発表内容を含む)。37,TEO→TEとvisual processingが進むが、TE域は扁桃体と相互に結合している(皮質扁桃体投射ニューロンは皮質第6層にあり、その逆投射系の存在も確認されている)。TE域を背半部(dorsal)と腹半部(ventral)に分けて調べた結果では、扁桃体の内側基底核(mediobasal nucleus)は側頭葉TE域のventral域とのみ、外側核(lateral nucleus)はTE域全体(dorsal もventral域も共に)と結びついている(方向性?)。更にこの領域はventral striatum(側坐核と嗅結節)と結合している(に投射している)。側坐核や扁桃核は辺縁系に属し、また視床下部へは扁桃体から分界条(stria terminalis)という名称の線維束が投射しており、その他に、扁桃体腹側遠心路(ventral pathway)が分散性(diffuse)に側坐核や視床下部に投射している。前者は主として扁桃体の中心核から、後者は主として内側核と皮質核から出て、各々本能的行動誘発および意欲・情動の発現に関与するものと思われる。
詳しい研究がなされていないが、TG(temporal pole region, area 22の最前部)野は聴覚連合野に属しているが、amygdalaとの結合もみられることから聴覚系のemotional expressionに関連する領域と思われる。このサルのSTS周囲の皮質域は1970年代に小生(川村)が詳しく調べた研究領域で、後背部は言語系に、前腹部は情動系に、また、中央部を含めて全域的にpolymodal neuronsが存在し、数種の感覚様態の入力が集中convergeするところである。また、このsulcal areaは前頭前野にある程度のtopologicalな局在性をもって出力連合線維を与えている。さらに、STSの背側(または上側)壁には体性知覚や味覚からの興奮も入ってくる。hodologicalには、この領域は、areas 5,7 と、さらに興味深いことに島域(insula)との結合が証明されている。
前述したように、この側頭連合野に属するTG域やTE前方域皮質は鈎状束(fasciculus uncinatus)と呼ばれる両方向性の連合線維によって、眼窩面皮質(OF, orbitofrontal cortex)と相互に強く結び付けられている。これらの皮質 (TG,TE,OF) はともに扁桃核と密接に相互結合をもっている。つまり、後連合野のamygdala related cortical areaと前連合野の同類領域とが相互に結びついている(川村、車田)。実際の所、前連合野から後連合野へ向かう成分は、反対方向の成分に比べて少ないが、その殆ど全部がこの鈎状束を通るもので、この皮質皮質間の投射成分は無視できない重要な構成であると小生は常々考えてきた。最近、宮下 (1999-2000) は、特定の連合線維路に障害を加えるという実験を施行して、鈎状束を通る前連合野から後連合野への投射成分は記憶の想起に関係することを証明した。ここで注目したいことは、感覚性言語野と結びついたpassiveな emotional expression / recognitionが、運動性言語野と関わりをもつactive emotional actionと結びついて、その結果この前頭前野内で「組みかえ処理」がなされ、然る後に前補足運動野(pre-supple-mot.area)に伝えられ、意欲に関係のある帯状回(帯状皮質運動野前および後部, ant. & post. cingulate motor cortexがある)との結びつきをもった補足運動野(supplementary motor area)、腹側および背側運動前野(ventral and dorsal premotor area)に興奮がシナプスを代えつつ伝播する。このimpulseの一部は随意運動を司るarea4の運動領へ入力し、そして錐体路系を作働させる。他の大部分は皮質の広い範囲から(動物が高等になるにつれて運動系関連皮質以外のところからも)皮質下の運動系組織である線条体(striatum)へ投射する。この線条体運動系は他に小脳系、網様体系の”不随意運動系システム”をも含めて錐体外路系と呼ばれ、globalなバランスを統御・調節的に司るところである。言葉をかえて言えば、「運動系のGestalt機構」と言えるところであろうか。このように複雑かつ精緻なシステムによりaffectusとmotorusのダイナミズムが脳内で演じられる。
付言すれば、ピアノやヴァイオリンを学習するタイプのいわゆる「体で覚える技術的性質をもった」記憶の習得は小脳・脳幹を中心になされており、その学習の習熟のプロセスは視床運動核(VA, VL)を介して順次大脳皮質の運動関連皮質に伝えられるものである。複雑なことにVA/VL核には小脳からの情報の他に大脳基底核からの情報が淡蒼球を介して入力されて来る。すなわち、「体で記憶する, knowing how (Ryle, 1949),やり方の知識;手続き記憶、procedual memory」と言われる脳幹・小脳・視床・(大脳)の系とより知的綜合に関わる「頭で記憶する, knowing that (Ryle, 1949),事柄の知識;陳述記憶、declarative memory」と言われる大脳皮質・線条体・間脳/脳幹の神経回路網という2種の運動系が調和して働いているのである。因みに、手続き記憶や陳述記憶という用語は人工知能の分野で生まれた(対立する)概念である。
最近の興味深い仮説としてvisual系のdorsal and ventral pathwaysになぞらえて、auditory系でも後連合野から前頭前野へ興奮の流れにdorsal & ventral pathsを相当させて、各々、「where」と「what」の機能を推定している(Romanski, Rauschecker,それにMishkin, P. Goldman-Rakicらのグループにより)。すなわち、視覚系について言えば、外側膝状体から第一視覚領皮質に達した視覚性興奮は、area 17からareas 5 & 7 へと向かう背側路と、area 17 からTEO,TE,TG域へと向かう腹側路と二つの皮質内経路をとって段階的に進行することが知られている。空間知覚とかかわる視覚認知路(where course)と丸・三角やさらには顔貌の表情の認知などに関わる視覚情動路(what course)とも言えよう。Romanski らは、聴覚系皮質にも視覚領と同様な形態・機能上の構成を一つの仮説として提唱している (1999)。
最後に、この項でテーマにした、視覚と聴覚との類縁性を如実に示すfMRI(functional Magnetic Resornace Imaging, fMRI, 機能的核磁気共鳴映像診断装置)を用いたヒトの脳の研究を下に紹介する(新潟脳研、中田力)。
実験は以下の如くである。日英バイリンガルの人で、初見(楽譜)演奏のできる―音楽家にはごく普通の能力である―ピアニストが楽器なしで(つまり実際に楽器を奏して音を響かせることなしで)楽譜のみを見せられたときに、脳に生起するfMRIの記録である。もちろん、物を見ているので第1次視覚野(VI)は反応しているが、第1次聴覚野(AI)は反応していない。つまり、視床MGB→AIという聴覚伝導路は興奮を伝えていないのである。ところが、驚くべき事に、音を理解する領域である聴覚連合野が働いているという所見を得た。詳しく云うと、VI、聴覚連合野の他に、言語読解と共通する左脳の視覚連合野と音譜の読解に特異的な右脳の視覚連合野が働いていた。このようにして楽譜が読解された後に、言語を理解するときと同じ領域である左脳の側頭葉の聴覚連合野(その内にあるいわゆる側頭平面planum temporale,PT,に一致する)が働き、ここで楽譜を音としてとらえていると考えられた。側頭平面は、Geschwindとレヴィッキー、1968、により右半球のPTより左半球のPTが優位に大きく、言語機能に関係すると考えられてきたareaで恐らく狭義のWernicke野に一致する(中田)。このprocessの中で、同時に左側の頭頂葉が働きここで運動知覚に関する3次元空間の情報に変換されると考えられる。実際に演奏される場合は、短時間経過した後に、興奮は恐らく前頭前野(前頭連合野)を介して一次運動野に伝えられ、指を動かす領域が働きピアノを演奏することになる。
以上は、rCBF(regional cerebral blood flow)などで脳の検査をしたとき、音楽教育を受けたヒトが音譜をみているが、実際には音を聞いていない状態(AIが反応せず)においても、聴覚連合野のニューロンは活動していて、音を文字化(或いは文字を音化)して理解していると解釈される。これをsupportする、もっと判りやすい例が身近にある。それは、難聴者(耳の聞こえない人)が、手話会話ないし手話ニュースで視覚的に言語化された音の連続を理解している(勿論、信号の形象化が加わっているが)と考えられる。また、囲碁や将棋の上段者が三五歩、白4六ハネなどの進行読み手の声をきいただけで対局の盤面をありありと描くことが出来るのも、また、ソロバンの有段者が指先を動かしながらものすごい速さで読まれる数学の暗算が出来るのも、聴覚連合野が働いて音声を文字信号として認識しているように思われる。更に話は高尚になるが樂匠ベートーヴェンのように、長じて難聴/絶聴に襲われた音楽家が記憶された響き(内聴)によって作曲し、内聴された響きを音符という一種の文字記号によって楽譜化(文字化)し得たのも、ヒトの脳における言語(文字)=響き(音楽)認知の深い関連性と、その認知に必ずしも外聴( AI)が必然されないことを物語る事例であるといえる。なお、付言するならば、Beethovenの場合、作曲にスケッチ帖が用いられたことは広く知られているが、このこともまた文字=響き認知とAIの関係ついての示唆を与えるものである。無論、楽譜は作家における文字と同様の機能を持つものであるから、作家が一度書かれた文章を訂正していくいわゆる推敲の作業と同じく作曲家は一度書かれた楽譜を訂正し、加筆しながら作品に仕上げてゆく。この作業は原則としてどのような作曲家にもあてはまることであるが、Beethoven程にこの作業の初歩的段階、つまりごく簡潔な、殆ど無意味に思われる着想の段階からスケッチ帖を用いてこの楽想を記し、それを少しずつくり返して訂正・推敲していった思索の軌跡を記録した作曲家も稀有である。おそらくは一般に考えられる、耳の聴こえない=外聴の不可能な音楽家などは有り得ないであろうという判断とは全く逆に音楽家にとって第一次的に重要なことは内聴であり、内聴された響きが楽譜化され、楽譜化された内聴が、内聴された響きそのものを訂正し、また訂正された響きが音譜を書き直させるという、要するに響き=概念化を相互的に繰り返してゆくことによって、音楽(作品)は成立する。但し、それでもなお、失聴した後のBeethovenが自らピアノを弾きながら作曲・推敲し、自分の作品の練習に立ち会って、例えば自分の弦楽四重奏曲がシュパンツィヒの四重奏団で演奏されるのを聴き、弓の動きから響きを感知したり、第九交響曲のオーケストラでの練習時に改めて楽譜に改訂・訂正の筆を加えたことは、演奏家の身体の動きを捉える視覚野によっても響き=文字が認知されることを物語っているのであろう。
なお、絶対音感は上のような音=文字機能について検討する際の必要条件であるとは考え難い。音の高さを認知する場合に音感には二種あり、何等か外聴された音の絶対的高さ(振動数的音高)を判断し得る絶対音感と、或る音と他の音との関連性を認知してゆく相対的音感がそれであり、重要なのは前者ではなく後者である。前者は単に音高を識別する能力に過ぎないのに対して、いわば音による思想の伝達を旨とする音楽(作品)を成り立たせるのは後者だからである。音感には音高の識別の他に強弱やテンポ(感)、響きの陰影等の認知が含まれ、音楽のいわゆる表現は相対音感によるものである。従って絶対音感ではなく相対音感の優劣が音楽家としての能力を決定することになる。このことは言語について考える上でも重要で、言語もまた相対音感によって認知される。特に詩に象徴されるように、文学は本来書かれるものではなく語られ聴かれるものであることを、優れた詩がしばしば優れて音楽的であることは注意されて然るべきである。
4]芸術(とくに音楽)における視聴覚合体論
ファウスト(F)とメフィスト(M)の対話
M:青息吐息の御様子ですね。御邪魔でしょうか。
F:胡散臭い奴だ。勝手にしろ。
M:私にとっては、聴覚系の方が面白そうですね。人間は視覚で物事を判断しすぎる。
F:認識・判断系のことを問題にしているのではない。そちらは大脳の出番で、今の勤めは小脳だ。
M:小脳の機能として、視覚運動反射とか聴覚運動反射とか、また、光源や音源の定位に関連するなど御託宣のようですが、大脳や脳幹網様体との関連も禄に顧みないでよくも言えたもんだ。さっぱり実体的イメージが湧きませんね。
F:小癪な。暇とアイデアを呉れ。小天地にそれがあるのなら連れて行け。
M:いつもの御性急がまたぞろ出てきた。あちらでは、前庭機能の考察も不可欠ですよ。これは聴覚機能と関係がある。どちらも第八脳神経でしょう。
F:昔の生成論は知らないが、前庭系と聴覚系との間には血筋の違いが明瞭なのだ。一方は古く羊水中で釣合いをとる時分から発達し、小脳への最初の闖入者だ。他方は、新参者だ。小脳界隈にはすでに強固な縄張がつくられてしまっていたので前庭組のように大挙して直接入りこむことがむずかしかったのだ。代りに延髄の蝸牛核あたりで擬似小脳組織を作っているんだろう。和衷協同という訳にはいかぬ。
M:独善がりもいいとこだ。ともかく、視覚系も新しい。視覚系人間などという言葉も耳にする。
F:その通りだ。しかし、聴覚系も同様に、いや、それ以上に新しい。
M:オランダ人は視覚的でドイツ人は聴覚的だ。レンブラントとベートーヴェンが象徴的だ。同じくゲルマン系だが。
F:いい加減なことを言って、話を逸らして貰っては困る。それに、皮相な現象論をふリまわす輩なんぞに用はない。
M:大変なお冠だ。とにかく、視覚と聴覚とは別物。視覚は間脳に入り、聴覚は延髄に入る。それに一方は粒子と波の折衷混体で、他方は単に波として源(みなもと)が説明される。
F:似た処もあるではないか。小脳でも橋核でも、縄張りがほとんど完全に重なっている。視蓋・小脳投射系に於いては、上丘も下丘も同声異俗。物質の最高段階にまで発展した形態などといばり腐っている大脳皮質にだって完全独立王国といえる場所は純粋に認識・分析系に属する所だけのようで、後連合野にも前頭前野にも重複したり隣接している部位がいくつかある。
M:視聴覚合体論か。我田引水とはこのことか。ところで、相補性とか多重感覚性とかおっしゃっているようですが矛盾を感じませんか。
F:唯我独尊的中枢は脳の中には一つもない。対立物は相互に浸透し、それらは高い段階で統一される。
M:物の臭みが強すぎて流行(はやり)ませんな。
F:意識が物質より根元的だと考えるような干からびた尨犬の脳髄からは、実りある発展はあり得ない。
M:羊皮紙綴りの知識は灰色だ。実際に験することだ。豊かな人生はみどり色だ。言い残してはございませんか。
F:耳目を属(しょく)すという言葉がある。眼を開いて物事に刮目するなら、萬物流転の位置を確と定めることも左程苦にもなるまい。眼を閉じてもそれができるだろうか。こう考えてみると、視覚と聴覚の連続性という意味が少しははっきりする。前方は網膜にまかせられるとしても、側方・後方はコルチ器官の助けが要る。これが相補性だ。脳研究者は、生理学的・生化学的言葉で説明ができない限り迷うものだ。
M:Cogito, ergo sum. Nihil sub sole novum.
以上のdialogueは私が15年近く前に唱えた視聴覚合体論である。これは脳幹の小脳-四丘体領域での重複を中心に動く物体の視覚的認識と動く音源の聴覚的認識に主眼を置いて論じたものである。
また、文学的遊びと言われるかもしれないが、視覚性と聴覚性が融合した叙景的な世界が美しく描き出され抒情性が与えられているものが、万葉集にも数多くみられる。
例えば、
ぬば玉の夜の更けゆけばひさぎお久木生ふる清き河原に千鳥しば鳴く(巻6、925) 山部赤人
春の野に霞たなびき うらがなし この夕かげに鶯鳴くも(巻19、4090) 大伴家持
この4]項では3]項の「聴覚と視覚-音楽と言語-」で論じたことを解説的に、やや内容に味を付け(られるものなら付け)て論じてみたい。舞台は大脳皮質である。メロディーや楽曲など動く音(それぞれが個々に特徴をもった音素は断続したものである)の連続性を時間的に保持させるメカニズムが聴覚皮質において、あるいは、聴覚系神経組織のなかで、どのように働いているのか。また、物体(視覚対象)の空間的に切れた線分や切れた絵画の要素を空間的に連続的にさせるメカニズムが視覚系神経組織においてどのように働いているのか。
なぜなら単なる音素の結合や単なる線や点の結合だけでは音楽たり得ず、また、絵画たり得ない。鑑賞者(すなわち受動者)の場合でも音楽や絵画の行為者(すなわち能動者)でも音(素)や形質(素)を相対的にゲシュタルト的に組み上げてまとめ上げる過程(プロセス)こそが芸術を作る”原始的”とも言えるsine qua nonである。
先ず、聴覚皮質における音の形質とその発展的プロセスについて考えてみる。ところで、前項に触れたように聴覚領皮質に達したインパルスは、AI/core(AI,R,RT)→medial & lateral belt(CL, ML, AL)→parabelt(STGc, CPB, RPB, STGr)と進行し、後連合野皮質内で、ニューロン活動に対応して音の質(テンポ、音色、ピッチ、和音、協和音etc)が形成され、変革され、消去され、また、再構成されるというように変化すると考えられる。これが鑑賞ないし聴受の際に脳内に起こるプロセスではないだろうか。
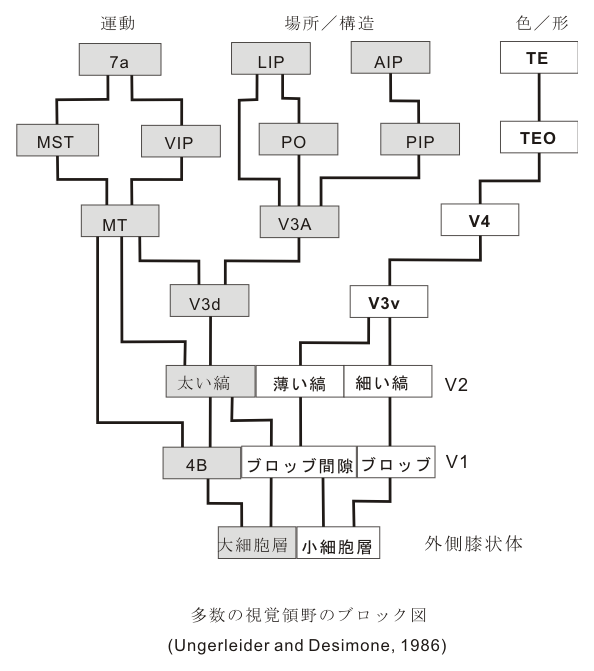
これに対して、より詳しく研究されてきた視覚系の方はどうであろうか?区分としては、VI(第一視覚野、視床のLGMからの入力が入る所)、ついでV2,V3,V4,MTその他にTEO,TE,更にPG(これは頭頂葉皮質に属する)などに区分される。PG野は本来体性感覚連合野として知られている領野で視覚系の背側路(dorsal route)が前頭前野に向かう領域である。また、ここは体性知覚と視覚とがdynamicに結合した立体的認知(この立体的という言葉は両眼による立体視という意味ではなく、物体の表面を斜めに傾けた時に生ずる奥行きの感覚などを言う)に関係する。視覚性特徴抽出ニューロンの種類としては(またはvisual cuesとしては)、色、動き、texture(物体表面の粗、緻などの感じや、手触りなど)、明るさの性質、また、点、縁、隅などの形態に反応するものが存在する。V-4領域のニューロンは色の認知に関係する。また運動の方向性シグナルはVIで最初に得られるが、MTニューロン(middle temporal visual area)の90%が刺激の方向と速さに著しい選択性を持っている。MTはPGに,更に前頭前野(主に背側運動前野)へとdorsal streamを送る。
また、V3A(static disprity, 静的視差、stereopsis) →CIA(手の形、向き、方位) →AIP;
MT(MST) →7a,VIP,LIP の流れがある。
そしてこれらの間にはヒエラルキー(階層性)がみられる。MilnerとGoodale(1995)らは、dorsalの流れは視覚-空間visual-spacial ("where" 何処) (後にGarwi らにより“how”, 如何に、vision for action, とされた)に関連し、ventralの流れは対象認知object recognition ("what" 何) (vision for perception)に関連するとの見解を持っている。そして更に情動emotionと関係をもつ。これら後連合野におこる視覚認知・認識の多様性は驚く程のバライエティーに富んでおり、連合線維の働きによってニューロンレベルで多様の組合せが出来て調和した活動が起こったとき、object(人物、風景)が描かれる下地が準備されたということになる。
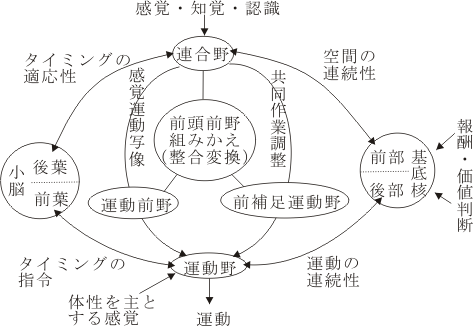
後連合野から前連合野への皮質皮質間投射について大きくまとめてみると、5野からは弓状溝上枝に隣接した背側域(dorsal premotor域)へ、PG (7a proper) からは前頭前野の内の主溝を含んでその背側域、46野)へTE(下部側頭野、IT)からは前頭前野の腹側域(12野)へ連合線維(皮質皮質間線維)を送る。また、頭頂間溝壁(IPS wall)内のAIP からはventral premotor 域へ投射する。それぞれの投射系はそれぞれ機能的な特徴を備えている。すなわち、dorsal stream は、背側運動前野(area 6) という一次運動野(area 4) 前方に隣接した運動関連域に直接投射するが、ventral strea の終止域は area 10 ( 一部area 45) に相当する非運動野である。 (この領域については、後に考察の対象になるであろう)。これら後連合野からの多種のmodules がそのまま能動的性格を有する一次運動野に送られてしまうとすると、おそらく個々の行為がばらばらとなってしまい、まとまった形の行為(action)は成立しないであろう。しばしば論じられているように、前頭前野を介する神経回路を経て運動系に高次神経活動が伝達されると言うことは、それらのmodule/informationが能動的発現をするに際して、その前段階で前頭前野で組みかえられ、”reduction”を受け、行為(行動)への意義(意味)づけをする上に必要な converter の役割を演じているのではないかということを想像させる。
聴覚情報も視覚情報も同様に背側経路と腹側経路を介して後連合野から前(頭)連合野(又は前頭前野)に段階的に形成された感覚情報を送っている。とすれば、この異種感覚間の統合や連合は前頭前野で生起しているのであろうか?「yes」という報告が、2000年5月にNature(Vol. 405, 18 May, 2000, 347-351)に発表された。この論文(Mustersら)は異種感覚情報を連合して時間的に処理するという前頭皮質ニューロンの特徴を見事に捉えた研究である。すなわち、サルにある特定の周波数の音を10秒間聞かせて覚えさせ、その後特定の色を選ぶという課題を与えた(sound-color matching task)。この課題を遂行中、前頭前野(背外側野で記録)のニューロンは音に対して選択的に反応し、その多くが音に結びついた色に対しても反応した。この聴覚と視覚の相関は、サルが課題(task)を上手く遂行できなかった時にはみられなかった。このことは何を意味するのか? 行動学的に意味のある視・聴覚間の連合が表現されるときには、前頭前野のニューロンを含む神経回路が存在し、その活動が時間差を統合・処理するという形で起こっていることを示している。つまりこの神経回路を構成するニューロンの正しい活動によって、上のtaskにみられる passive (聴受)から active (選択)への移行に際して、active (=運動)の方向性を規定し、またどのような手順を踏んで運動がなされるべきであるかの運動の順番=時間的流れが整えられるばかりでなく、この活動には言語(認知)、空間を含む認知、情動表現を包含しており、つづいて起こる運動表現の前のいわば“仲介”の段階にみられる現象であることを考慮した場合に、高等動物、とくにヒトにおいて、順序ある行動や言語の表現、すなわち、意味ある行為や思考を形成する上できわめて重要であると考えられる。
われわれが絵画や彫刻を鑑賞するとき、そこに表現された内容や概念/思想を、それらが具象的にしろ抽象的にしろ、理解しようとする。この際の脳内活動は、視覚性言語、つまり文字(信号の信号)の認知で、主として角回(gyrus angularis)で行われる。更にむずかしい解釈であるが、高度な視覚信号の”音楽的”認識(音階、音程、ピッチなど)である楽譜の読みというものがある。これは視覚と聴覚との接点である縁上回(gyrus supramarginalis)を中心とするWernicke areaでなされると思われる。
このように、受動的鑑賞に限っても、音楽的性格と絵画的性格とは相似かよった点があると思う。脳の機構の上でも視覚系同様聴覚系にもdorsal routeとventral routeの存在が最近注目されている。これらは上側頭回(superior temporal gyrus)から直接に、または上側頭溝(superior temporal sulcus)を介して間接的にprefrontal cortexに投射しており、dorsal routeは眼球運動やワーキングメモリー(作業記憶または作動記憶と訳されているが、目的志向的な課題や作業の遂行に関わるactiveな記憶を言う。すなわち、状況に応じて適切な情報を(意識的、意図的に)取捨選択して一時的に保持して操作して柔軟に処理する認識機能を言い、運動-行動-に導くもの。コンピューター上での作業をするために、一時的に内容を保存しておく場所をworking areaと言うが、この動作の類似からこの名称が用いられている。)(後述)の空間タスクに関与し、ventral routeは物の同定や情動に関与しているという。そして視覚系の後連合野から前頭前野への背側および腹側の2系列の投射がみられると同様に、聴覚系にもdorsal & ventral pathsが存在する。この際可成り視覚と聴覚のpathsは似かよった皮質・皮質間投射系を持っているように思われる。そして聞きことばの認識、すなわち、原始感覚性言語野Wernickeも包含されて、dorsal streamは主溝より背側へventral streamは主溝より腹側に投射される(サル)。
脳内の構成上または機構上に存在すると推量される視聴覚間の類似性と補充性は、以上のように、小脳運動系にも大脳認知系にも認められる。これまでの論旨の道筋から結果されるのは、音楽と美術/建築との間には、深い関連性がみられるということであり、音(音楽)・色や形(美術)・言語(文学)がニューロンの働きによって相関的な関係に置かれ、最終的には一体化されてゆくということである。このような脳内における異質感覚領野の相互的通約性は、これらの異質感覚が各々の領野において異なる対象の内に passive に認知し、かつ人間が自らの手によって active に作り出す対象の内に自覚的・無自覚的に投影してきた“表現”そのものに内在する共通性によっても逆証明されるであろう。美術・建築の内に、また音楽の内に記述され、発音されて響く言葉のリズム秩序と同一の数比例が存在するからであり、しかもここにみられる視覚に映る調和=聴き取られる調和という関係は、究極的には人間を包み込む宇宙=自然そのものに内在している数的比例の響きの調和と一致するものとして捉えられるものである。
ひるがえって考えてみれば、視覚、聴覚をはじめ多くの感覚様態を連合野のニューロン活動の連鎖回路により階層性の高いレベルに総合ないし統合していく仕組みが脳の機構として存在している。そこでは、受け取りつつある感覚(情報)が、現段階ではわれわれの理解を超えた「何か」-それはニューロン活動の総和としての産物(所産)として説明される性質のものではあろうが-、が加味されて、大脳辺縁系の活動を中核とする情動(パトス)と大脳新皮質活動の最高の産物である言語(ロゴス)の活動と結びついた時、高い芸術的受用の段階に達する。更にそれが、音楽にしろ、絵画にしろ、書道にしろ、彫刻にしろ、芸術家の手にかかって表現される時の前頭葉の働きはどんなものか。それが演奏なり創作として表現されるとき脳内の運動系はどのように働いているのであろうか。(脳幹および小脳レベルの事象を含めて後述されよう)。
5]音楽の表現形態(演奏)と能動性皮質(前頭葉)について
ここでは、能動的受用すなわち、前頭前野が関連する運動系/能動系発現の問題を取り扱う。すなわち、絵画を描き、音楽を演奏する際の運動系の働きである。視覚系において、演劇を観ること→ものの動きをみること、と、聴覚系において、音楽を聴くこと→音の流れ、動きを聞くことは、どう関係づけられるだろうか。演劇または観劇(ドラマ)や音の響きとして捉えられる運動に内在する秩序が存在し、これらの運動を支配している「調和」のとれた秩序を人は何故に心に思い留めることが出来るのであろうか。「本能」と稱するものが、この「調和」を感じることができるのか。然りとすれば、それは脳の作用であり、それを感知し得る能力の構造的基盤が必ず脳に存在するからに他ならない。
視覚系も聴覚系も恐らく体性感覚(触、圧、痛、温、冷覚)系も、各々の視床特殊核から入力をうける第一感覚野から後(皮質)連合野内(各々、後頭連合野内、側頭連合野内、頭頂連合野内)を段階的に興奮が進行したのち(形態的には連合線維により)、背側の流れ(dorsal stream)と腹側の流れ(ventral stream)-小生はmiddle streamとしてSTs域への入力、更にこの合流点は知覚性言語野に発展する、を加えて考察している――を作って前頭葉に興奮を送っている。
前頭葉はしばしば触れたように、能動的活動を発揮させる領域で、広義の運動系すなわち、アクション、行為、自発性を計画し、命令し、実行させる脳の大切な領域である。
Charvis & Pandya(1976)は、視覚(VI,VII,VIII)、聴覚(AI,AII,AIII)、体性感覚(SI,SII,SIII)-この分け方自体の妥当性を考察することなしに論をすすめるとして――から前頭葉への投射域をサルで調べた。そして第1連合野(VI,AI,SI)、第2連合野(VII,AII,SII)、第3連合野(VIII,AIII,SIII)からの終止域を弓状溝および主溝の周囲部で詳細に調べた。その結果、以下のような興味あるデータを得た。すなわち第1連合野からは弓状溝の前後の領域にまとまって終わり、第2連合野からの投射は弓状溝の腹側部領域に顕著に重なり合って終止するが、第3連合野からの終止域はもっと前頭前野の広い範囲の領域に分散して余り重複せずに認められる。この意味づけは難しいが、この三つの主要感覚系の後連合野から前連合野への入力、すなわち、受用からアクションへの変換の第一ステップを集合と分散の要素として見てみると面白いかもしれない。
「ここに、坂上論文考察のこと」
それにしてもareas 4, 6の運動系領域への直接投射は全くみられないことは注目に価する。以下にみるように、後連合野からの直接入力が終止する前頭葉域から、幾つかの段階を経て、(pure) voluntary area (area4)に興奮が伝えられるのである。一方、サルの大脳皮質―線条体結合をまとめたPercheron et al. (1984)によれば、主溝を中心としたその周囲領域からは専ら尾状核に、その背側(area6, 前頭前野)や眼窩面に近い腹側野は尾状核と被殻と両方に投射しており、第1運動野、体性感覚野を中心とする領野は専ら被殻に投射する。その他に頭頂葉(尾状核と被殻両者へ)や側頭葉(特に尾状核へ)の広い領野からも線条体に投射することが明らかになっている。
ここで昔から言われている“古典的”運動の概念、すなわち、2種類の運動について触れておく。一方は、第一次運動野からのみ皮質下の運動領野(脳幹、脊髄を含めて)へ興奮を送る随意的、意識的、計画された、運動、いわゆる錐体路系の関与する狭義の運動で、他方は錐体外路系が関与する運動で、この皮質・線条体投射系は、第一次視覚領(area 17)を除く大脳皮質の広範囲の領域から投射がみられる調和・調節的、且つ、全体的にバランスをとろうとする広義の運動である。
線条体については電顕を用いた微細構造の研究などはかなりの成果が挙げられていたにもかかわらず、10数年まえまでは、神経回路や機能的な研究、まして発生学的研究、については遅れていた。すなわち、大脳皮質や黒質から入力をうけ、視床や淡蒼球・黒質に出力を与える前脳内の構造物で運動の統合に関係しているという類の大雑把なものであった。線条体は細胞構築上、不均一でモザイク状を示し、アセチルコリンエステラーゼ(AchE)に強く染まる部分と弱く染まる部分からなっている。前者がmatrix compartment、後者がpatch compartmentとよばれる区画区分である。patchはstriosomesとも呼ばれ、matrixにより糸巻き状に囲まれた複雑な配列をしている。霊長類ではpatch/matrixの境界/区別は明瞭でNissl標本でも分かるが(サル)、鳥類や爬虫類では不明瞭である。発生学的にはラットでは、胎生13~15日(E13~E15)でpatchが形成され、E18~E20でmatrixが形成される。黒質からのドーパミン含有入力が最初のpatch形成に強く関与するといわれている。入力としては、古い辺縁皮質関連野からは主にpatchに、新皮質からは主にmatrixに、また、ドーパミン投射に関しては、腹側被蓋野(VTA)、黒質緻密質(SNc)の背側部からはmatrixに、SNcの腹側部と黒質網様部からはpatchに投射している。なお、腹側線条体に投射するVTAのドーパミン細胞はコレチストキニン(CCK)、ニューロテンシン(NT)を共有する。また、神経伝達物質や受容体の分布にもpatchとmatrixとでは差が認められる。すなわち、 patchには、サブスタンスP(SP)、ニューロテンシン(NT)、エンケファリン(ENK)、ダイノルフィン(DYN)、グルタミン酸脱炭酸酵素(GAD)、グルタミン酸(Glu)などの免疫反応が強く、D1ドーパミン受容体、M1ムスカリン様コリン受容体、μ-オピエート受容体などが高濃度にみられている。一方matrixにはソマトスタチン(SOM)、NT受容体、AchE、コリンアセチルトランスフェラーゼ(ChAT)、チロシン水酸化酵素(TH)、ニコチンアミド・アデニン・ジヌクレオチド・フォスフェート(NADPH)-diaphorase、パルブアルブミン(Pv)、calbindin-D28K(Calb-D28)などの免疫反応が強く、D2ドーパミン受容体も高濃度に存在している。
以上の研究成果をわれわれは持っているが、現段階でこれらのpatch/matrix構造および伝達物質、含有ペプチドや受容体に分布や関連などについて意味づけをすることは困難である。高次神経活動という知覚認識、情動、運動が有機的に結びつく領域として、この線条体を含む大脳基底核の分野の研究は益々重要性を帯びてきている。基礎的研究のみならず、パーキンソン病、舞踏病、精神分裂病などを含む神経科、精神科領域の難治性疾患の原因究明のためにも非常に重要である。
さらに議論を敷衍させれば、この後連合野からの多種感覚性入力重複皮質域は運動性の言語野及びその周辺に発展する領域と考えられる。言語表現能力を備えた領域自体ないしその近傍域でいわゆる音や形象の「組みかえ」がなされ、音楽の演奏となり、絵画の創作表現として発展するのであろう。別の表現で言えば音や形の内容が思考され解釈され、その結果が自発性の発露となって表現されてくるのである。
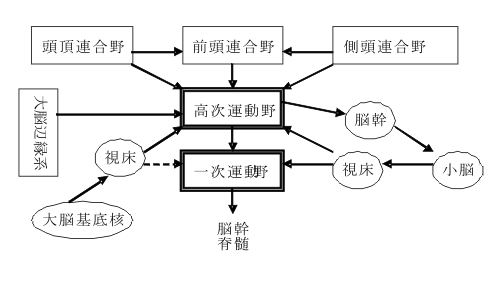
フローチャート式に書けば、感覚野→後連合野→前連合野→高次運動野→第一次運動野という図式になる。この初出の高次運動野とは、この十数年来Evertzや丹治らにより研究されている領域で、運動前野(area 6もっと詳しく云えば6DR, 6DC, 4C, 6Va, 6Vbを指す)、補足運動野(内側面6野)、帯状皮質運動領が含まれる。
運動前野(premotor cortex)は、area 6に相当する領野で背側と腹側に2分され少しずつ違った働きをすると考えられている。Area 6の前方のarea 8は前頭前野(frontal eye field)と呼ばれ、眼球運動の発現と調節に関与する。内側面の帯状溝より上(背側)にある部分でarea6に相当する領野は補足運動野(supplementary motor area)が、またその前方に前補足運動野(presupplementary motor area)がある。またこれらとは別に帯状溝に埋まった領域に帯状皮質運動野(cingulate motor area)が存在する。これらを総称して高次運動野と呼んでいる。すなわち第一次運動野(area4)以外の運動野をこのように呼ぶ。
現在、サルの実験から種々の運動領について明らかにされていることを簡潔に下にまとめておく。
○前頭前野:入力は頭頂葉からと前頭前野。物体の認知→動作への変換→運動のサブプログラム形式→筋活動の出力司令。
○補足運動野:PenfieldとWelch(1949)によって発見された。領域的に区分され、下肢、体幹、上肢、顔の動きを電気刺激で誘発できる。欠落徴候として、強制把握とか、動作の順序の組み立て障害がある。
○前補足運動野は前頭前野から入力をうけるが補足運動野はその入力をうけない。
以下に丹治順氏の著「脳と運動」より引用させていただく。
『このように、運動野が多数存在するのはなぜだろうか。その謎を解く鍵は、運動の誘因と目的性ということ、そして運動を行う状況・局面への対応ということにある。運動を行うきっかけは多岐にわたるが、生体をとりまく外界の情報や生体自身の身体情報、そしてすぐに脳に収められている記憶情報が、時に応じ、さまざまな組み合せで運動の発現を促すことになろう。次にそれらの情報を使って、行おうとする一連の運動の目標を設定し、そのための動作の手順や種類を選択し、それらをどのような時間的・空間的パターンで構成するかという過程に至る。このように、外界や体内の情報および記憶情報は、運動を行おうとする意図の発現のためにも、運動の選択・企画・構成のためにも必要なので、運動の高位中枢へと常に送られる必要がある。それらの情報は、個体にとって意味のある情報として利用ができる程度にまとめられている必要があるが、大脳皮質の連合野に求めることができる。ところが連合野は一次運動野とは直接には結びつきが少なく、密接なつながりは高次運動野を中継して行われているのである。したがって、周囲の状況を認知し、その中での自己の位置づけや全身状態を統合的に認知し、さらに運動の目標設定や組み立てに関連した記憶情報を参照するために、連合野の情報がまず高次運動野に送られ、そしてそこを介して一次運動野へと送られることになる。
高次運動野の脳内の位置づけを模式化して図示すると、下図
のようになる。前述のように高次運動野は大脳連合野から広汎に入力を受ける。他方、大脳基底核と小脳からも、運動の組み立て、構成や調節に必要な情報を受けている。このような脳内の結びつきに基づいて大略的に位置づけをするならば、高次運動野は運動発現・調節のための情報入力と、運動出力の情報形式の橋渡しをする、インターフェースを構成すると表現して良かろう。』
音楽作品に内在する「表現」は、それを演奏者が“運動”(すること)によって成立する能動的表現(特に量プラス速さの組合せによって、また呼吸によって変化する瞬間的力によって)と、それを聴くことによって伝達される受動的表現(聴受におけるパトス/アフェクトウス)との2つの面に分けて考えられる。後者の受動的表現については、前項で多少論じた。ここでは聴受におけるアフェクトに対比されるものとして、前者の問題、すなわち、演奏におけるアフェクトの問題に主たる焦点をあてて考えてみたい。
音楽における演奏によって左右される表現としての聴き手の心の状態(聴受されるものによって作り出される状態、パトス/アフェクトウス)は、演奏(者)の情動/感性(パトス/アフェクトウス)に基づく運動(これについては以下に更に考察する)と無関係ではない。厳しい見解であるが、少なくとも音楽における運動=演奏は、自己の運動がもたらす結果の如何に対する判断力によって統御される。この際、その統御=判断の根拠となるものは美的センスである。
ここで、①情動(pathos/affectus)と運動(随意運動・不随意運動、自律性運動など広義のものとして当然に理解されなければならない)との関連について、また、②そもそも脳の機能として、運動とは何を包含するものか、という2つの問題について考えておきたい。
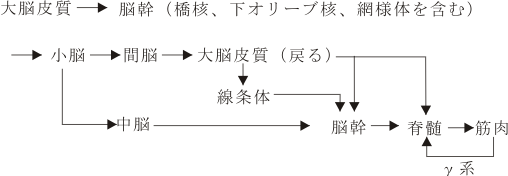
そもそも運動(motorus)とは何であろうか。大脳皮質area 4の運動領野が興奮することにより、そこにある第5層の大型錐体細胞からの神経突起内の電気的伝導が下行性投射神経路を通って、脳幹にある脳神経運動核ニューロンおよび脊髄にある前角運動神経細胞を興奮させ、その活動が身体の筋肉を随意的に動かすといういわゆる「錐体路系」の神経投射路である。勿論これだけの理解にとどまっていたのでは医学生の解答ととしては不十分で不合格である。人はいつも随意的な意図的な運動にだけ支えられているわけではない。身体全体の運動系を調和させ、統御し、運動を円滑ならしめる機構が動物にどのような形で備わっており、機能するのであろうか?前脳(終脳と間脳)が未発達の動物にもすでに、環境に働きかける上で必須の基本的なシステムが能動的作用として備わっている。それはいわゆる「錐体外路系」といわれるシステムで、あるものは小脳を中心とした回路、他のものは大脳基底核を中心とした回路(網)、またあるものは脳幹網様体や脊髄を中心とした運動系の回路と言われるものがある。この「錐体外路系」は動物一般に基本的な形態的基盤を提供するもので、「錐体路系」よりも系統発生的に古く、強固なシステムである。
さらに、呼吸、循環、発汗、消化、食欲、性欲などいわゆる植物性機能に関係する自律神経性の運動がある。この自律神経系は交感系と副交感系という互いに拮抗する機能をもつ二つの系統から成っており、これまた動物に本来的に備わっている基本的なシステムである。そして、大脳辺縁系を中心とする情動系回路と形態的にも機能的にも密接に結びついている。下等動物を含めた動物一般として脳幹レベルにみられる呼吸、循環、唾液分泌、性興奮、意識、睡眠に関係する自律中枢および視床下部、下垂体(ホルモン系の元締め)、さらには扁桃核、海馬を連絡する情動・記憶の神経系や副腎皮質、甲状腺、性腺を含むホルモン調節系が厳然として存在する。さらにより高等な動物になると、前脳が発達するにつれて、大脳基底核、大脳皮質、とくに新皮質、その中でもとくに前頭葉皮質(この中に前頭連合野あるいは前頭前野を含む)が著しく発達してくる。
このように、動物の発達史に沿ってみると、脳幹から大脳辺縁系さらに大脳基底核(線条体、黒質など)ついで大脳新皮質というように脳機能は高次化していく。
大脳新皮質において、種々の感覚(体性、聴、視、味、嗅など)が知覚・認知され、認識機構が後連合野で施行される。発達する皮質連合野について言えば、聴受にあずかるpassiveな後連合野がはじめに機能を発揮し、その次の段階で、動物間のコミュニケーション、協同作業を中心とする労働、それに加えて、道具を使用することを可能にした環境(自然)への能動的働きかけなどの協同社会体制の中で、前頭葉、とくに運動性(能動性)言語野を含む前頭前野が発達してくる。内側面には中古皮質に属する帯状回(とくに前方域area24)がある。サルが環境を変えられたとき、自分で新たに適切な課題解決を考え出した時のみ報酬を与えるとういうパラダイムを施行させて、「意欲」に関する研究を行った(丹治ら、1999, Science)。その際に特異的に反応(応答)するニューロンが、帯状回前部で見出されている。ここは外側面の前頭前野域との相互連絡を有する領域で意欲に関係する領域と言われている。なお、帯状回や前頭前野はA-10 Dopamin投射(中脳VTA域から)が密に存在する。このように前頭葉にも意欲、情操、道徳、運動などに関係する異なる機能をもった皮質域区分があり、この領域が障害されると運動性麻痺、運動性失語、自発性(あるいは運動性)低下、情緒不安症、などいわゆる能動性機能の障害に相当する種々の症状が現れる。
ここで視床から大脳皮質第一感覚野に到達した電気的な神経細胞の興奮が運動領に達して運動(=能動的行為)を起こすまでの過程を簡潔に整理しておく。動物の感覚器官として嗅覚と味覚は重要で考察を欠くことができないが、-とくに嗅覚刺激は扁桃体に入力し、海馬も含めた大脳辺縁系と深い関わりをもっている- 視覚系と聴覚系に焦点を合わせて論じたい。眼の網膜の錐体細胞と杆体細胞および内耳にあるコルチ氏器官の有毛細胞という感覚細胞が各々外界からの視覚的および聴覚的刺激を受容し、電気的興奮が生じ、いくつかのシナップスを替えたのちに間脳の外側および内側膝状体に伝達される。そこから視・聴覚の興奮は同側性に視床皮質線維を通って第一視覚領(V-1, primary visual area)および第一聴覚領(A-1, Primary auditory area)に伝わり、この大脳皮質の分析器(パブロフ)で最初の処理(認知作用の第一歩)が行われる。
上述のように、視覚領においても、聴覚領においても、そこに存在するニューロン群は特徴ある個性をもって居り、多数の細かい領野に区分されている。このように皮質レベルで形態学的に区分されていても、必ずしも充分に機能的に解明されているわけではない。とくに視覚領では現時点で解明された部分が可成り多いが皮質聴覚領野の亜域について視・聴覚系間の類似性を比較し、類推する興味ある見解がルネサンス期、啓蒙主義時代の知識人、自然科学者、哲学者らによって披瀝されている(後述する機会があればよいが本質的かどうか不明也)。われわれは現在ニューロン活動を実験的に調べる手段を得ているので、その機能類推を安易に受け容れることなく、科学的に検討し、実験データの上に立って考察することが要求されている。
この両感覚系(視覚系と聴覚系)には、①頭頂連合野を経由して前頭葉の背側運動前野(area 6)に向かう興奮の流れ(dorsal pathway,背側経路)と、②側頭葉前部~極部(temporal pole)を経由して前頭前野の主溝に接する腹側部域(area10)に向かう興奮の流れ(ventral pathway, 腹側経路)が霊長類(サルで調べられた)の脳で存在し、視覚情報はこれらの2つの経路にわかれて処理されることが識られている(Ungerleider and Mushkin, 1982)。背側経路は自己を中心として(egocentric)物体(視覚的又は聴覚的対象)がどこに(where)、どのように(how)配置されているかという、視覚刺激の空間的位置や動きの情報を処理することに関与している。他方、腹側経路は、視覚的または聴覚的事象(events)がGestalt(形や色;旋律や音色、別項で詳述する)としてどんなもので(what)、どんな意味ないし評価(evaluate emotionally)がなされるかということに関与している。勿論これに単純な説明を下すことには相当な無理があろうだろう。今後の研究が必要である。このプロセスを通じて視覚系では、単純な形(点や線)から複雑な形象を認知し、色彩や動きなども加味されて情動的な視覚反応も示すようになる。視覚系のヒエラルキーの機能現象論としておく。同様に、聴覚系にも振動数に対応した(単)純音、基音から、複合音、(協)和音、不(協)和音、メロディー、ハーモニー、さらに、リズム、ピッチ(これは、脳幹、小脳レベルの事象が大きく関わるようである)の問題も入ってくる。音の長さ、音と音との間の間歇(休符)、音階を伴った動的働き、リズム等々。これらは、小脳の半球後部が関係しているという(後述)。付言すると、小脳虫部中央部は視覚・聴覚の動く物体の認知に関係していることが1940年代からSniderらより知られている。
興味あることは、egocentric perceptionに関わりのある背側経路が、第一次運動野(4野)への直接投射はないが、4野と強い結合関係をもっている運動前野(6野)に投射を持っていることとと対照的に、Gestaltや情動の処理と関わりのある腹側経路は4野にも6野にも直接投射することなく、その前段階として前頭前野(皮質)の10野に投射していることである。
視覚系についても、聴覚系についても、後連合野で受容receive(ないし受用geniessen)し、処理された上述の情報が、目的行動(演奏するとか彫刻するとか)に進行されるためには、適応的な行動(運動、動作)をする必要がある。その意味で視覚および聴覚の後連合野から前頭葉への2つ投射経路(dorsal and ventral paths)を考察することは重要で興味あることである。
まず、「背側経路」について。頭頂葉と運動前野のニューロンを調べてみると、類似した応答特性をもったニューロンが両域で存在していることが知られているいる。従って、背側経路の空間情報は比較的”ダイレクト”に運動の企画(プログラミング)に利用されているようである(Wiseら、1997)。視空間、音空間から運動空間への写像変換(conversion)がこの経路を用いて比較的容易に施行される。その運動を行うための視覚情報処理経路と考えられている(Goodale and Milner, 1992)。
次に、「腹側経路」ついて述べる。この経路は、上述の如く、視覚および聴覚領内を腹前方に流れ、すなわち、側頭葉前方域に興奮が伝わったのち、鈎状束を通って前頭葉の(少なくともサルの段階では)主に前頭前野の腹側部あるいは腹外側部に投射する。この経路で運ばれる"Gestalt"(形象、全体的ハーモニー、色彩、音色も含めてこの用語を使用することにする)の情報は、背側経路で運ばれる空間情報と比べると運動情報との関係は直接的ではなく、比較的独立しているようにみえる。視聴覚刺激が運動情報に変換される前に、前頭前野の前頭葉腹(外)側部(10野)のニューロンを発火させて、情報を処理し、それを運動系に伝達させるという方式を脳はとっているのである。
以上数行は既述されているか? 要検討
視覚系でこの問題を研究している坂上によればこの前頭前野のニューロンの約半数は、視覚刺激に対して短い潜時(100ミリ秒前後)で発火活動を変化させるという。しかし、その応答は、色や形といった視覚刺激の物理的な特徴ではなく、視覚刺激がどのような行動を指示しているかを反映するものである(筋運動そのものには応答しない)。このような情報を”behavioral significance (connotation)"(行動的意味)と呼ぶ。感覚情報でも運動命令でもない、その間を媒介する「意味の表象」である。前頭前野腹外側部においては、この「意味の表象」は、少なくとも視覚情報との関係では、「腹側経路」の出力に基づいたものとなる(Wilson et al (1993): Sakagami & Tsutsui (1999))。色や形といった視覚情報は、空間情報と異なり、特定の筋運動と結びつく必然性はない。逆にいえば、特定の情報を行動に結びつけるために、学習が必要となる。事実、前頭前野の「行動的意味」をコードするニューロンは、学習によってその活動を、より適切な刺激―反応関係を反映させる方向に変化させる(Niki et al. (1990): Asaad et al. (1998))。よって、前頭前野腹外側部からの出力は、もはや感覚情報ではなく、運動プログラミングに利用可能な翻訳された行動情報として、運動前野や補足運動野に送られる。
以上述べてきた2つの視覚情報処理の経路と前頭葉の関係をまとめると、下図のようになる。長期にわたる学習の結果、すばやく感覚―運動の情報変換が行われるようになった頭頂葉-運動前野系によって準備された運動の候補を、下部側頭葉-前頭前野腹外側部系が、その場その場の状況に即した形で取捨選択していくことにより、より適応的な行動選択を可能にする、という図式である。最近、この図式が視覚以外のモダリティー、すなわち、聴覚のモダリティーにも当てはまるという研究もでてきた(Romanski et al. (1999))。 本稿では時期尚早であるという批判をうけることを承知の上で、視覚と聴覚の類似性について大胆な推論を展開している。
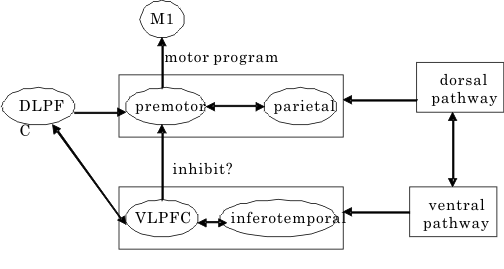
図1 2つの視覚系路と前頭葉 M1:一次運動野、DLPFC:前頭前野背外側部、VLPFC:前頭前野腹外側部。
坂上らは、ニホンザルに、異なる色や形の組合せからなる複合視覚刺激を使った、go/no-go反応タイプ(ある状況(条件)下では行動をgoせよ(行なえ)、また他の状況(条件)下ではno-go(行なうな)というタイプの実験パラダイム)の選択的注意課題を学習させ、その課題遂行中に前頭前野のニューロン活動の記録を行った(Sakagami & Niki (1994))。この課題では、サルは、色に注目するときには、形を無視して、たとえば、緑色が提示されたときには、go反応を、赤色が提示されたときにはno-go反応を行う。形に注目する場合には、○でgo,+でno-go反応を行う。組み合せによっては、たとえば赤い○は、色に注目するときには、no-go反応を意味する刺激になるが、形に注目するときには、go反応を意味する刺激となる。前頭前野腹外側部の「行動的意味」をコードするニューロン、たとえば、go反応を意味する刺激に強く応答するニューロンは、この刺激に対して、サルが形に注目しているときには、発火活動を上昇させるが、注意を色の次元に切り替えると、即座に発火の上昇は見られなくなる。発火活動の変化は、明らかに視覚刺激の呈示に同期しているのに、注意の文脈の変化で同じ刺激に対する応答が逆転してしまう。このことは、前頭前野腹外側部のニューロンが、単に個々の刺激-反応関係の「行動学的意味」をコードしているだけでなく、複数の刺激-反応関係を包含・制御するルールを実現するようなネットワークを形成していることを示唆する。
以上のように脳の機能としてみたとき、狭義の運動は随意的なものであるが、広義の運動は情動や意欲を含めた能動性機能と見なすことが出来る。この視点に立って演奏における運動とaffectの問題をみてみることにする。
ここで、次のことを指摘しておきたい。すなわち、演奏者によって、音譜の中に読み取られ、同時に聴き取られた「響き」に内在する運動とは何か?このように聴き=読み取られる「響き」は次のように聴き取られる。
1)聴き手のaffectusに働きかけるために必要な、要するに人間のaffectusを動かすための響きとして、
2)音の自律的法則として、 ――或る音と別な或る音との響きの関係、バランス、多くの音の関係が形成するバランス(音体系)=コスモス、
3)上の法則の中核をなす(二重の意味での)数的秩序を意識的に無意識的に、
4)秩序へと、もしくは超越的、存在の根拠=第一の数的秩序の源泉へと聴き手を最終的に導くために必要なものが、どのような響きであるかを考慮しつつ、
おそらく上の4)において、いわゆる「精神」もしくはきわめて鋭い感受能力を働かせて実際の響きが上の1)-4)の条件を満たすものであるかどうかをロゴス的に判断しつつ、演奏=表現=音楽の全体的支配・統御(Pathologos)がおこなわれると考えられる。
能動的(activeな)logosの座であるBroca言語野を含む前頭前野と、受動的(passiveな)logosの座であるWernicke言語野は連合線維により結び付けられており、各logos野の近傍の前頭前野下部(眼窩面皮質)と側頭葉下部及び極部はそれぞれ投射線維系により辺縁系(扁桃体、海馬傍回)と結びついている。このように、passive及びactiveなlogosとpathosの座である大脳辺縁系とその近傍の連合野が相互に密な関連性を形態的に(連合線維の結合という点からみても)有していることは注目すべき事である。ここで、興味ある重要な点は、後連合野で高度に処理された興奮(=脳の活動)が、狭義の運動野であるarea 4,(6)に直接に伝達されるのではないことである。皮質皮質間結合の研究の結果は、一旦、motorisches Sprach-Zentrum近傍の前頭葉皮質においてシナプスを形成する(ニューロンを替える)。すなわち、この前頭前野における仲介域で「組みかえ」(PathosとLogosとの質的向上を含んだ相互浸透)を起こしたのちに、運動系皮質域(補足運動野→前運動野→運動野)に伝達されるのである。感覚性の認識が情動と結びつき、その融合体が直接的に有機体の運動系を働かせるという方式をとらずに、生物体の最高機能をもつといわれる前頭前野を介して間接的に運動系に投射するという方式をとっている。情動のaffectusはamygdala又はaccumbensからの入力により、意欲のimpetusは前頭前野と強い相互結合をもった帯状回前方域(anterior cingulate area )からの入力(出力もある)により維持されている。この際、後述する視床下部からの入力やDopamin系からの入力が加味される。この経路を通った後にmotor areasが活動してはじめて演奏におけるアフェクトの関係が了解される。この際、重要な役割を演じているのは、先にみた小脳の学習機構と共に、線条体の運動調節統御、バランス的・全体的・ゲシュタルト的な能動的な役割であろう。
この際注目したいことは、ネコあたりのレベルでは、皮質線条体(尾状核と被殻)路の起始細胞の存在域は前頭葉の運動関連領域(主としてareas 4 & 6)に限局されているが、サルの段階になると前頭葉の主溝周囲および腹側部や頭頂葉、側頭葉、17野を除く後頭葉、さらに内側面では帯状回前部(’70-’80年代の研究で明らかにされている)を含んで広範囲の皮質領域にわたっていることである。前にも述べたように、この皮質線条体投射は単なる純粋な運動に関係しているのではなく、能動性の総合的、協調的な運動機能を動物が発揮する上で重要なシステムであると考えられてきた。そしてヒトになると、おそらく、質的相違が加味されて、ほとんど全皮質域から線条体への投射が発達してくると考えられる。
先に大脳基底核、とくに、線条体の細胞構築と線維連絡の概略について述べたが、ここで少し視点を変えて、大脳基底核・視床・大脳皮質を神経回路のシステムとしてみた時、どのような意味が付加されてくるかについて考えてみよう。Alexander, DeLong, Strick(Ann. Rev. Neurosci. 9: 357-381, 1986)は、以下に述べるように複雑な神経回路網を整理して、このシステム回路は、形態、機能ごとに並列的チャンネルを作っており、個別的、並列的な情報処理(parallel processing)をする場であるという概念(parallel channeling)を呈示した。この大脳基底核-視床-皮質系の線維連絡は、閉鎖回路を形成し、運動系、連合系および辺縁系の3ループに分けられる。また、線条体という構造物を主役に考えれば、線条体は線維連絡および機能的に①感覚運動線条体、②連合線条体、③辺縁線条体(尾状核頭の腹側部で主に側坐核と嗅結節の深層部にあたる。免疫組織化学的に中心部 core ―運動系と連絡し、辺縁系のインターフェイスとしても働く―と周辺部 shell ―視床下部、扁桃体など辺縁系と連絡する―に分けられる)に分類される。以下に簡単に説明する。
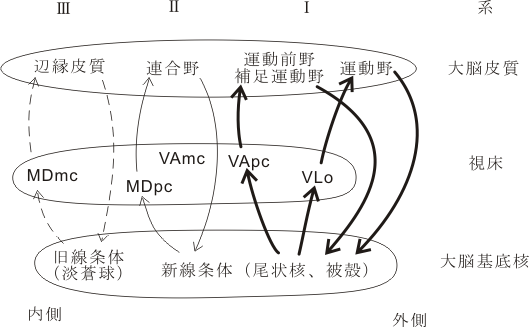
Ⅰ.運動系ループ
1)運動感覚系ループ(この系は運動の高次機能に関与する)
知覚運動野→被殻(運動系線条体)→淡蒼球外節/内節(GPe/GPi)(外側部、運動系-淡蒼球))→VLo→運動野
2)固有補足運動野(SMA-proper)系ループ
SMA-proper→被殻→GPe/GPiの中間部(補足運動野関連淡蒼球)→VLo内側部→SMA-proper
3)前補足運動野(pre-SMA)系ループ
pre-SMA→尾状核(CN)の外側部→GPe/Gpiの中間部→VApcの外側部→pre-SMA(pre-SMAはヒエラルキーが最も上位にあり、このループは運動のプログラム、準備に深く関与する)
4)運動前野(PM)系ループ
PM→CNの外側部→GPe/GPiの背内側部→VApcの内側部→PM
5)眼球運動系ループ
前頭眼野(FEF)/補足前野眼野(SFEF)→CNの中央部→黒質網様部(SNr)の外側部→VAmc,MDpl→FEF/SFEF
Ⅱ.連合系ループ(この系は認知などの高次の脳機能に関与する)
前頭連合野/頭頂連合野→CNの外側と腹内側を除く大部分及び被殻の前部(連合線条体)→SNr及びGPe/GPiの背内側部(連合系淡蒼球)→MDpcの中央部と一部VAmc→連合野
Ⅲ.辺縁系グループ(この系は動機づけ、情動行動に関与する)
辺縁皮質・扁桃体・海馬→辺縁(または腹側)線条体→腹側淡蒼球→MDmc内側部→辺縁皮質
ここで、述べておきたいことは、脳の運動性(能動性)機能として最近の注目されているワーキングメモリー(作業記憶)と演奏との関連についてである。ワーキングメモリーとは、情報の処理と保持の並列処理を可能とするシステムであると考えられており、操作によって記憶の内容が書き換えられるような短期記憶を言う。当初の意識的コントロール下で譜面を見て手の指を動かす随意運動主導の反応から、旋律に対応した運動系のコントロールが練習を繰り返す(小脳学習)ことにより自動化されて、演奏をする段階に働く機能である。この時の脳はどのように働いているのであろうか。記憶の貯蔵庫から、その情報(旋律)を取り出し、一時的に(でも)保持し、それに対応した運動反応に変換する(運動系プログラムへの変換)。同時に並列的に、次の旋律を長期記憶からほぼ自動的に検索する。このような、運動制御を行いつつ並列的に次の旋律を検索するという音楽演奏におけるワーキングメモリーの脳内局在はサルの実験では前頭連合野が関連していると言われている(Goldman-Rakic, 1992)が、ヒトの脳でPET(positron emission tomography, 陽電子放出断層撮影装置)やMEG(magnetoencephalography)を用いて調べてみると、前頭前野に限局しているものでなく、広範囲に及ぶ活性化が認められるという。概念化(Begriff)する能力を備えたヒトの脳は他の動物とは質的に異なる情報処理、課題遂行を統一的に施行することができるのであろう。この際memoryが動的に関与する高等動物において忘れてならないのは、言語系、第二次信号系(パブロフ)である。この言語系はsensory, motor, supplementary と3つの位の領域しか考えられていなかったが、最近のrCBFやfMRIの検査の結果はことばを発する時には大脳皮質領域内の広い範囲で多数の領域のニューロン群が反応していることをわれわれに教えている。この多両域言語 (logos) 反応ニューロンの分布をみると、言語と結びついたヒエラルキーの高い情動との結びつきを考えても、皮質内情動反応ニューロンの分布も側頭葉前部や前頭葉腹側部を含む広い範囲に出現するというようなデータが見い出されて然るべきという感じがする。音楽と言葉の響きの関係を考察(解釈)する上で大切な視点と考える。いずれにせよ、ヒトのような言語活動可能の高等動物においては、言語野と前頭連合野(前頭前野)の発達により、高度なaffectusを発揮/発現させることが可能となった。そして、高度な信号である音譜を解読し、形成されたPathologosの基盤の上に立って演奏(作曲)という能動的活動(運動)がなされる。そのゲシュタルト的響きは数的比例の秩序に支配されており、聴受者の(また演奏家の)聴覚系機構内で判断される。これが、具象化されたPathologosの吟味であり、これが再び前頭連合野を介して統御され演奏(運動)にフィードバックされる。以上を支える脳内の神経回路は形態的に皮質皮質間結合が①後連合野内で、②前連合野内で、③後連合野と前連合野との間で、存在するという形で証明されている。この点は強調されるに価する。
6]音と言葉と形象
先に演奏家が楽譜を見たとき、しかも初見のとき、音を聴かずともその音を理解/認知していることを述べた。すなわち、第一聴覚領(AI)は興奮しないが聴覚連合野と言語の領野(Wernickeの一部)は活動しているという所見である。このことは、何を意味しているのだろうか?また、これから、どのようなことが考察できるだろうか。そのためには、どのような実験を組み立てたらよいか。たとえ明白に証明することができなくとも、ここに隠されている重要な事実を解く鍵を得るべく研究を前進させたい。
ここで音楽と音声言語について考えてみる。空気、鼓膜、耳小骨、そして内リンパ液の振動として、気相、固相、液相内を伝播した波の振幅が増強されて、内耳のコルチ氏器官内で振動エネルギーが電気エネルギーに変換され、聴神経を通って脳内に伝達される。形象的には、いわばアナログ信号のデジタル化として捉えられる変換である。 [デジタル化について調べてから後日加筆すべし] !!!!中枢神経系、つまり脳と脊髄は、ニューロン(神経元)という細胞単位が、一定の規則をもって、シナプス結合を介して連結し構成されている。神経突起(=軸索)内を信号が送られるときは電気信号、すなわち、電子の流れとして迅速に伝えられる。音の場合も、脳内を蝸牛神経核、外側毛帯核、下丘、内側膝状体、聴覚(領)皮質へと聴覚系の諸核(ニューロン群)を段階的にシナプスを介して伝達される。
音楽も音声信号も視床から興奮入力をうける第一聴覚領(AI)のレベルでは、音素という同じデジタル化された電気信号をうけていると言える。中田グループや酒井グループ、その他の研究者たちが、近年、人で言語の意味処理や音韻処理をさせたり、連続音(メロディー)などを聞かせたとき、fMRI法、PET法、光トポグラフィー法などを用いて脳の機能局在を調べている。その結果、頭頂葉や前頭葉の言語域(聴覚系、視覚系ともに)を含む領野で機能領域の解離が認められると報告されている。この際の言語域にはWernicke areaやBroca areaも含まれ、聴覚性および視覚性言語の意味処理と音韻処理の選別がなされることが明らかにされている。すなわち、中田らは、ヒトに言葉を含む音楽を聴かせてfMRIを用いて調べたとき、いわゆる聴覚連合野に活動が現れると同時に、右側後頭葉前部と左側島域に活動を認めている。そして、前者を音楽の要素、後者を言語の要素と解釈されるデータを発表している(? 再検討すること)。また、酒井ら(2000a (PRONAS)& b (Neuroimage); 23会日本神経科学大会2000c)は、種々の脳機能画像法(機能的核磁気共鳴法、fMRI;経頭蓋的低頻度磁気刺激法、TMS;光トポグラフィー)を用いて後連合野および前連合野における言語情報処理について研究し、以下のように幾つかの点を明らかにしている。すなわち、①会話文を文節単位で提示して文章を全体として理解するという課題を行ったとき、後連合野(ウェルニッケ野)の角回(gyrus angularis)下部においては、音声による提示(聴覚条件)でも文字による提示(視覚条件)でも同時に活動を示したが、前頭葉のブローカ野においては、聴覚条件ではその上部域が、視覚条件では下部域が、というように部位選択的に活動していた。この結果は、聴覚連合野および視覚連合野からブローカ野へ至る経路は各々独立しおり、ブローカ野では文の情報を分離して処理していることを示唆している。② ブローカ野において統語処理を調べた結果、(i)代名詞が名詞を示し得るか、(ii) 2つの名詞が同じ文法的関係(主語・目的語)にあるか、を判断するという課題で、(i)ではブローカ野の背側部に、(ii)では腹側部に部位選択的に活動するというように、異なる統語処理モジュールに関係する領域が互いに解離していることを示唆する結果を得た。③前頭葉の中央部において、意味処理(名詞句と動詞句の意味の整合性を判断させる)と音韻処理(動詞句のアクセントの位置が正しいか誤りかを判断させる)とが独立して行われるということを示唆する有意な皮質活動が得られた。
これらの事実は、少なくとも聴覚系の場合、言語であれ音楽であれ「聞き取られたもの」は電気信号として第一聴覚領皮質(AI)に到達した後に聴覚連合野/ウェルニッケ野へと進行し、“ウェルニッケ”野以降に「言語/ことば」と「音楽/おと」に選別されることを示唆している。つまり、聴覚連合野/ウェルニッケ野で受け取られて、そこで、電気信号の種類別を認知し、各々の信号に応じてその後の機能(活動)領野を選択し、そこへ送信する、そのような機構の存在を考えさせられる。それはどのような形で存在するのだろうか?音楽も言語も基本的には音素から構成される音の響きを持っており、これがいわゆる広義の聴覚連合野(あいまいな表現として“ウェルニッケ野”)で認知される。このように考えると、従来からの臨床上の欠落症状を基に漠然と大脳皮質の広い範囲を採って定義された“ウェルニッケ野”と称されている領域は狭い限局された意味での“言語野”ではなく、むしろ、“響きを感受し選別する場所”として捉えた方が事実に即しているように思われる。すなわち、ここで種々の響きの違いが、おそらくデジタル信号の組み合わせの違いに基づいて読み取られ、或る形態のものは言語として、また別のものは音楽として選別されていくように思われる。そしてこの選別は、先に述べた後連合野から前連合野に至る背側経路(dorsal route)と腹側経路(ventral route)を介して伝達され、前頭葉の活動に反映されることになる。ここで考えておかねばならないことは、前頭葉の働きというのは、他の大脳皮質部位と大きく異なっているという点である。つまり、ここには、視覚や聴覚などの特定の感覚様相に個別に関わって特定の処理形式にのみをおこなうといった局在性はみられず、個別の諸機能の上に立って、その全体を統括的にコントロールするような役割を持っていると考えられる。
視覚および聴覚による(味覚、体性感覚、嗅覚が多少加わるにせよ)形象Gestaltや空間(Raum)の認知情報は、ヒトの場合、言語認識という高度に抽象化された記憶概念として、一方は情動が加味されて側頭葉からventral route(解剖学的には鉤状束)を通り、他方は自己の状態を位置的に時間的に同定するという空間認知として、頭頂葉からdorsal route(解剖学的には上縦束)を通って前頭葉に伝達される。いわゆる二重の認知機構である。この際、後連合野には、音と音声言語(語りことば、voise)、色/形と文字言語(letters; sentences)という選別機構が存在するように思われる。
一般に動物において、情動と認知―自己の認知をふくめて―の原始的(primitive)、受動的(passive)な環境からの情報はこのように後連合野で処理されたものが前頭葉に伝達される。そしてはじめて、その有機体すなわち動物がもつ最高次の神経活動として、能動的(active)且つ共働的に外界に対して働きかける精神活動を発揮することができるようになる。先の酒井らによるヒト前頭葉連合野における言語処理の研究結果が如実に示すように、視覚的にも聴覚的にも高度の言語法則の処理がブローカ野およびその近傍で行われていると考えられる。具体的な実証を待たなければならばいが、この言語情報処理の研究で見出された高次の規則・文法・法則の処理は、言語のみならず、別記の脳磁計(magnetoencephalography; MEG)による解析結果からしても、これと同様のレベルで音楽や美術の領域における響きや数比例を対象とした(情報)処理にもあてはまると判断し得る。大いに関心のもたれる所である。
わが国の研究者によっても、MEGや脳波を用いて前頭連合野(=前頭前野)の機能について調べられている。すなわち、佐々木ら(1994,1996,1999)は注意を集中して計算をさせたり、音楽を心の中に思い浮かべさせたりしたとき(musical imagination, play music in mind)、被験者の両側前頭葉にfrontal midline theta rhythm (Fmθ、 5-7 HZ前頭葉の割に広い範囲に発生することが後にMEGによって明らかにされたこのθ活動は、最初脳波(EEG)で前頭正中部に最大値となる活動が認められた、Yamaguchi, 1981をみよ、のでこの "midline"という名が付けられたが、"mental"と読み替えた方が適当と思われる)といわれる信号を見い出して左右の前頭葉-右側が多少より強い-が活動していることを観察している。音楽を聴いた時には、このθ波がとくに右前頭葉が活動していた。また、Asada, Fukudaら(1999,NSL 274, 29-32)も最近、MEGとEEGと同時併用して、2分間集中して行われるこのメンタルテストの際にみられるFmθの起源は、帯状回前部を含む内側前頭前野皮質であろうと推察した。また佐々木ら(1996)は、ポリフォニー音楽(モノフォニー音楽では有効でなかった)を心に思い浮かべたときとか、禅僧の瞑想時にこのFmθが出現すると報告している。
最も高次に発達している前頭連合野の最前方部(Frontal -pole)においての研究は困難でヒトにおいてこれ以上の明らかなデータは得られていない。われわれが現在計画している音楽の聴受または内聴という実験の結果、この領域において、上述に関連する活動が得られるならば(その可能性は大なりと考えて実験を行おうとしている)、昔から、真・善・美とか、道徳・倫理・宗教・審美・正義などの高次の機能に関連する領域として捉えられてきた哲学的概念が、脳の高次機能として自然科学的にメスが入れられることになるであろう。人間が古来より、星空を眺めて瞑想し、宇宙空間の法則性を無自覚的にせよ認知するとき、JS Bachの受難曲の響きを天上からの絶対美を瞬間的判断力で認知するとき、もしもそうなれば、これらの異なる形象が本来一つのものとしてある種の記憶の概念とマッチしていると言えることになろう。形と音と色と数の本質的対応と昇華は前頭前野でなされる。建築は凍れる音楽である( )、音楽は魂が無自覚的に行なう算数(数学)である(ライプニッツ)、といった表現は、こうした脳の高次機能に媒介される能動的な精神活動をいみじくも鋭く言い当てていたことになる。
そして、その場合、総合的に音楽は右脳優位に、言語は左脳優位に送信されることが推察される。しかし、脳の認知活動は、多様性に富んでおり、しかも全体的であり、また種々の概念形成段階に質的差違がみられる。このことから考えて、音楽家や演奏家を被検者に選んで彼らの音声認知活動における脳内活動の流れをたとえば画像解析することにより、貴重な資料を得ることができよう。その際、音楽(メロディーのみ、言葉が含まれる)や音声言語の音韻処理、意味処理、概念化のレベル(日常生活レベルか思想的、哲学的に高次のレベルか)の区分などを含めて、データを脳の機能局在(後連合野か前頭前野かなど)と時間的連続性などに留意して解析することが出来れば、大きな収穫が得られることは確実である。将に音楽家と自然科学者の共同でのみ遂行される作業である。
このような研究を行うにあたって考慮すべき点であるが、「音楽」と「言語」が電気信号によって区別されるためには、この二つの事象の違いが明らかな形で信号化されている必要があろう。例えば、言語と音楽の響きの違いの手がかりとして、音楽には存在せずに言語に存在するものに注目してみる。この相違は「言語」における特有な現象である「子音」の使用に深く関わっていると考えられる。「子音」は持続する「母音」の響きを切断し、いわば言語のモールス信号化を行なう。これに対応するものとして、音楽の場合には休符、もしくは最初から持続の長さを決定し、一定の時間の経過の後に必然的に空白/切断が行なわれるべく定められた音符(の持続・時間性)による切断をあげることができる。従って、「子音」つまり本質的に「音」を持たない切断する「音」は音楽には存在しない。一方、言語とは逆に、音楽に存在して言語に無いもの、もしくは乏しいものは音の高低を法則に則して決定する、数比例関係によって捉えられる振動数(比)の関係である。
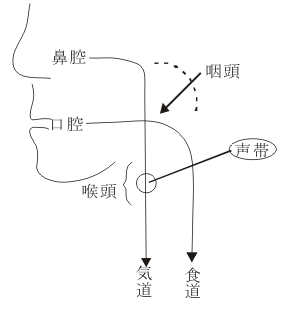
一歩しりぞいて考えてみて、一般のサルやチンパンジーやヒトの音声はどのようにして作られるのであろうか。これらの“高等な”動物では、空気の通り道である喉頭の内腔にある声帯ヒダ(この内に迷走神経支配の声帯筋や声帯靱帯が入っている)が振動して声が生じるという機構になっている。声帯ヒダは前庭ヒダとともに喉頭の内腔の壁に両側に張り出しており、その間隙/声門裂(=声門glottis)を空気が流れ、迷走神経の働きにより声帯の緊張や声帯裂の幅を変化させ、声帯ヒダを振動させ、ここで作られた空気の振動を咽頭を通って口腔と鼻腔などの付属共鳴腔で共鳴させることにより音声が生まれる。実は上述のこの機構はヒトにおいてのみ認められものでサルやチンパンジーでは様子が異なっている。すなわち、人間以外の哺乳動物では、喉頭の軟骨が咽頭内に高くとび出して鼻腔の後方にはまり込んでいるために、①鼻腔―(空気の道)―喉頭と ②口腔―(食物の道)―食道では、独立した立体交差路となっているので、声帯で作られた空気の振動を口腔内で共鳴させることがほとんど不可能である。すなわち、動物では声帯で作られた空気の振動は専ら鼻で共鳴されることになる。因みにヒトでは、喉頭の位置は低く、普段は咽頭の中にわずかに飛び出ているだけで開口しており、食物を飲みこむときだけ持ち上がって、喉頭蓋によって蓋をされるという風な信号付き交差点方式になっている。
ところで咽頭筋、喉頭筋は舌咽神経(IX)と迷走神経(X)により支配されており、舌筋は舌下神経(XII)により、また、口腔や鼻腔の感覚や運動は三叉神経(V)と顔面神経(VII)により支配されている。これらの神経核(神経細胞/ニューロンの集団)は脳幹にあり、これらの諸核には、大脳皮質運動野に広い領野を占める舌や(音声)発生共鳴腔器官の代表野(representation field)からの神経線維が投射して、神経活動を調節している。ヒトでは1歳頃までに母音を主とした発声/会話が可能になり、2~3歳頃には子音の発声も殆んどできるようになり、言語野皮質も発達して、意味処理可能の会話が出来るようになる。すなわち、脳を解剖生理学的に音声言語の観点からみれば、皮質代表部-脳神経諸核-支配筋およびこれらのfeed-backメカニズムが発達・成熟したということになる。
発声に限らずさまざまな機能が形成される胎児期、新生児期、幼児期は重要で、特定の機能の発達にとって、重要な意味をもつ、臨界期といわれる「その時期に有効な刺激が必要な適切な時期」がある。この分野の研究は視覚機能についてとくに詳しい。第一次視覚野には左右の目からの情報が分離して縞模様に並ぶ眼球優位コラムというものが存在するが、ネコの実験によれば生後2-6週間の「臨界期」に、このコラムが形成される。また、臨界期のネコに縦縞だけを見せて育てると、大人になっても横線をうまく認識できない。大脳皮質視覚野における神経細胞の反応を調べてみると、縦方向の線に反応する神経細胞が多くなり、逆に、横線によく反応する細胞が少なくなっていることが明らかになっている。他の機能についても同様の発達経過があり、各々の臨界期に、特定の機能を獲得するために関連する神経系の形態形成に必要な化学物質を動員する機構が生体内に存在すると考えられる。ヒトの青年期(思春期)における精神発達の重要性について12]項で考察されるが、Freud の精神分析の理論を待つまでもなく幼児期、児童期以前の身体的および精神的発達は医学的観点からも教育的見地からも重要である。
Bloomfield(「言語」)によれば、母音(vowels)とは、音声に舌あるいは両唇の閉鎖・摩擦・接触を含まない音であり、その範囲内の変更を与えること《modifications》により、母音には数個の異なった音素(a, e, i, o, u, etc)が区別される。しかし、これらの音素の間の差違は主として舌の位置の差違にあるという。母音に対照して、その他の音、すなわち、閉鎖音<stops>・ふるえ音<trills>・摩擦音<spirants>・鼻音<nasals>・側音<laterals>は子音(consonant)と呼ばれている。また、彼は、鼻音(m,n,η)、側音(l,r)および母音をまとめて、楽音(musical sounds)とし、閉鎖音、ふるえ音、摩擦音をまとめて、噪音(noise-sounds)としている。もっともこの点は、各国語の音韻構造や発音、表現法によって異なる見解を成立させると考え得るので、とくに楽音としての規定には注意が要求されるであろう。
サルやチンパンジーやヒトの赤ん坊の世界は「アー」とか「ウー」とかの母音の世界で、脳、発声、共鳴器官が発達するに従って音の断絶化が可能になり、子音を使用して声音による伝達、すなわち、音声言語の世界が入ってくる。
(以下メモ)
①音声のオシロスコープによる波型
②幼児の成長過程における言語の獲得=子音の獲得。
この2点は問題の検討に役立つと判断される。
音楽に素養のある人が音符の連続としての楽曲を黙読して「聴いている」ときは、分析「音」を視覚的に捉えているのであり、小説(の文章)を理解しながら読んでいるときのように文字化しているのである。音楽家(作曲家や演奏家)は自分の認識と経験の深さに応じて楽譜を見たとき“頭”の中に楽譜から読み取った響きを持つことが出来る。高い水準に達した演奏家とは、ある楽譜を“理解”したとき自己の脳内にそれに適する“響き”を形成することが出来、その響きにできるだけ一致させて自らの外的響き=演奏を作り上げるart, techneを持っているヒトのことを言う。この場合、原初的な、たとえば作曲家が聴き取った音の記号化である音符・楽譜を響きとして、しかも言語野において読み取りつつ、音の響きを文字記号として認識しているのである。一見極端に聞こえるが、音楽を聴受し演奏することは、高いレベルで、知的会話をやりとりしているように抽象的認識の過程、即ち概念化、の作用が基盤にあって成立しているのである。更に音の流れ、調べの中に意味や思想をもたせることが出来るが、これにもレベルの相異が認められ、比較的にレベルの低い、たとえば直接的に感情に訴えかけるものから高度の思惟に関わりつつ、たとえば神的・超越的世界を問うことが出来る(バッハのマタイ伝の二つのアリア、第39番と第49番を参照)。このうちの、とりわけ後者の世界は完全に高い次元のロゴスの世界であり、感情をこめて演奏するパトス、アフェクトゥスの世界を越えて昇華された内容に立ちいたっている。これは後連合野での実際の響き(演奏)に直接的に結びついている感覚、感性的認識の段階が、知覚性言語領域の機能と結びついて、自己を中心とした空間認知(identityの設定)作用を通じてのdorsal pathwayと、形象的、情動的認知(affectusの設定)作用を通じてのventral pathwayが、前頭葉に興奮を伝えることによって、はじめて可能になる。なお、この場合の空間認知は、哲学的に言えば、自己(存在)の世界内における定位、すなわち自己の在ることの深い自覚であり、神学的に言えば、神との対峙によって、神の働きの内に在る自己の確認ということになる。上に述べた演奏→響きの認知→ロゴスの次元への昇華もしくは高次化=形而上化とはこのことを指す。(なお、「マタイ」における第39番、40番、二つのアリアの神学的・音楽的表現(法)の具体的説明は附論を参照のこと)。
このときに背側経路の方ではsensoryが写像的にmotorに速く(または早く)伝わるが、腹側経路の方では”情”が加味されて前頭前野で組みかえ(conversion)られた結果が、運動司令・実行の系に遅れて伝達される。Affectusに訴え共鳴された音楽の演奏が“空間に響きがふくらむ”ものを持っていると表現されるのはこのことを言う。良い響きとは、人間存在を包み込む何か知られざる宇宙の秩序に合致した空間秩序の認識へとヒトのロゴスの働きを促して、かって古代ギリシャにおいてプラトンがイデアのミメーシスとして捉えた哲学的コスモス=宇宙空間を現成させると言ったらよいであろうか。この最高次の認識はわれわれが現在研究対象とすることが非常に困難な前頭前野の最前部と能動性言語野が相互に干渉して機能するものであろうか。昔、Economoが審美領域や倫理領域の機能域を前頭葉の最前方部に想定した苦心の作図を思い出す。
以上の聴覚系(音楽に関する)の考察は、視覚系(絵画や文学)にもあてはまるように思われる。とくに、視覚=色という光波の問題は、聴覚の音波の問題と同様に捉えうるように思われる。形・色の要素と楔形文字や象形文字の段階を経て信号化・概念化可能な高次信号文字言語という問題を、脳の働きとその進化・発展の歴史性から考察することが出来よう。この際、聴覚空間よりもより明白で顕著な視覚空間について、自己の世界(外界)内定位との関連において人間の精神の発達過程を考察することもできよう。ところで、視覚表現においても高度に抽象化された概念を表現することができる。或る図象においてそこに描かれている図柄とは必ずしもイコールで結び得ない、異なる存在をアレゴリーとして認知させるのがそれで、たとえばヘビの図は知恵を示し、リンゴは原罪を表すなどこのアレゴリーの手法は中世までの宗教画に色濃く反映されている。また、形と音ないし色と音色との結びつきを強く内面に感じとることの出来るヒトは芸術家に多い(cf. 絵をシンボルとしてみたとき、エコノロジーEconology)。
メモ:
{# 音と文字(言葉)と形の変化・行為の三位一体の芸術(=ドラマ)
声楽、宗教音楽、ギリシャ悲劇、オペラ}
メモ:
以下に、フルトヴェングラー、Wilhelm Furtwängler, 1886-1954) 芳賀訳(新潮社版)よりの抜粋を載せる。
「すべて偉大なものは単純である」これは芸術家のための箴言である、と言うのは、何よりまずその「単純」という言葉が、「全体」という概念を前提としているからである。ここで言う「単純」さとは、「すべてを見通して」「突如としてこの一挙に」正しくその「全体」をつかむ、という意味である。この言葉における「全体」とは、決してただそれ自体のために分離した世界の一部である、というだけではない。一部分にはちがいないが、それはこの世界をその「全様態」において反映する部分なのである。「全体の中に、魔神的なものが存在している」と芸術家ゲーテは言っている。非有機体的な世界はこの種の「全体」を知らない。非有機的な世界は限界というものを持たず、ただ無限にひろがろうとする。ここで言っているような意味においての「全体」という概念を持ちうるというのは、我々自身人間として、有機的な生命に所属しているからなのである。私たちは有機的に思索し、有機的に感覚する。すべての有機体、すべての植物、すべての動物はこの意味において、私たちにとって一つの「全体」を形作っている。
7]Gestalt 認識の生理学
形あるものを部分の集合とみて各々の位置(Stelle)に配置して(sto, stellen立てる、置く、据える)全体として統一的に組み立てられたものが形態(Gestalt)である。心理学ではそれは感覚されたものの総体であるとか、あるいは、感覚の概念を「形態」とか「それを構成する部分の総和以上のものである全体」とか一見「謎」めいたことを言う。しかし、最近の脳の科学の研究は、視覚系認識についても、さらには聴覚系認識についても、可成りの説明が可能なところまで進展しているように思われる。たとえば、視覚的認識とは、物体からの反射光が網膜を刺激し、(詳しく言えば錐体細胞(cone)及び杆体細胞(rod)→双極細胞→網膜神経節細胞の順に興奮が伝わり)、それが視床の外側膝状体に伝わり、シナプスを代えて、大脳皮質の第1視覚野(area17)に伝達される。つまり大脳皮質のこの領野での反応が感覚として認知される第一歩である。次に、area18,area19、area37、…→と後頭連合野おいて伝達され、その過程の中で、点→線→単純な形(丸や三角形など)→複雑な形(図形、景色、ヒトの顔など)へと知覚作用としての認知/認識が深まっていく。その形態的基盤は、その部位にある特殊に分化した神経細胞の反応を反映する形で、いわば階層的に低次から高次へと質的発展をとげつつその認識過程が進行する。画像(図形)の認知、その連続性の認識である図形の連想(あることを思い出したときに、その記憶に関連した別のことを思い出す現象)作用も側頭葉にある視覚性連合野のニューロンが機能の一端を担っているとして理解されている。すなわち、サルを用いた宮下らの実験で、図形を2個ずつペアにして学習させる。そしてそのサルの側頭葉ニューロンの活動状況をその一方の図形を提示した後に、他方をみせて調べた。すると、時間の経過と共に活動性が徐々に高まってくるというのである。
以上みてきたように、stepwiseに進むヒエラルキーを持った図形(画像)認識のGestalt構成は視覚系領域で起こる脳機能の反映であるが、主たる脳内活動の場が聴覚領に置かれている、音を素材とする芸術である音楽のGestalt機構も同様にみなすことができないだろうか。すなわち、視覚の場合は第1視覚野で感覚された切れた(不連続な)図形要素が空間的近接度を基準として、つながり(連続性)と切れ目(不連続性)を(再)構成する過程で、つまりフレージングによって輪郭を構築し、また再編していく(視覚のヒエラルキー構造をtranssynapticに視覚性刺激が進行する経過の中で)。同時に聴覚の場合は高低や強弱の差違に関わりなく時間的に隣接している音がつながり、音の流れ(stream)、すなわち旋律ないし何等かの音形が形成される。たとえば、一つの楽曲は一つの形態あるいは構造をもっているが、しかしこの構造、つまり幾つかの、それ自体としては単なる音以上の意味を持ち得ない音符によって構成される構造そのものが音楽「作品」たり得ているのかどうかは疑問であり、むしろそれ自体は単なる音である音符が、何等かの「意味」を形成しつつ結び合わされてゆくときに、はじめて音楽「作品」は成立するわけで、ここに一個の構造として形成された「意味」、別の言葉で言えば「表現」こそが、いわゆる音以上の「何か」を成すのであり、この構造としての「意味」・「表現」すなわち一個の音楽的「存在」そのものをここではGestalt と呼ぶ。このように、複数の連続した音符が一つの構造を構成することによって感性と統覚(apperception)的に結びついて楽曲が生まれる。それは内耳有毛細胞-蝸牛核-外側毛帯核-下丘核-内側膝状体を経て聴覚大脳皮質に到達した電気的興奮のauditoryI,A-II,Ep(ネコ実験)、更に連合野内の亜区域と連なる段階的/ヒエラルキーをもった聴覚系領域で起こる脳内のエネルギー伝達機構を基盤にしてのことである。聴覚皮質内には、これは他の機能皮質域についても言えることであるが、このような近類性質のニューロンが集団をなして存在している。このように構築された形態的基盤の上に立って、脳の機能として、不連続な単なる音から連続性(旋律)、総合性(和音、協和音、テンポ)がニューロンの集合と結合からなる回路網の活動から形成される。このように構成された一定の曲想の連想作用から、まとまった楽曲が作られる。ワーキングメモリー(作業記憶)の作用を通じて作曲がなされ演奏が行われ聴受される。この際、後連合野で知覚、認知され、統合された視覚・聴覚・体性感覚の複合体が前頭連合野で「組み変え」が行われる(前述4 ]をみよ)。
ここで、暫く、ワーキングメモリー(working memory)、事象関連電位(event-related potentials, ERP)および探索陰性電位について立ち停って考えておくことにする。ワーキングメモリーという言葉は流行語になりつつあるが分かりにくい用語である。ワーキングメモリーとは、一時的に提示された情報の表象を一定時間意図的に(activeに)保持することを言う。情報が一時的にpassiveに保持される記憶に対して、ワーキングメモリーは情報がactiveに保持され、不必要となれば消去(リセット)される。そして目的志向性が強い。因みに、短期記憶では、リハーサルによって長期記憶に転送されて処理される。この保持や処理の働きにより、判断や推論などの高次処理が行われる。ワーキングメモリーの脳内機構については、従来、前頭前野に局在する機能と考えられていたが、最近のfMRI(functional magnetic resonance imaging)やPET(positron emission tomography)で調べた結果、ヒトで大脳皮質連合野に広く、また大脳基底核、小脳、海馬など様々な部位で活性化が観察されている。このことは達成システムとしてワーキングメモリーを考えるとき、目標や課題への志向性が情動や意欲を基盤にして駆動されることを示している。また、事象関連電位は、動物(ここではヒト)が精神作業を行う際に生じる一過性の電位変動で、視覚性探索過程を反映するERP成分として視覚性(記憶)探索(陰性)電位(search negativity)についてはよく調べられている。視覚性符号を用いた研究から、それを反映する活動が視覚系の脳内経路と一致することから、人間の視覚系が外的視覚刺激の処理だけでなく、内的視覚表象の操作にも関与している可能性が示唆されている。聴覚刺激を用いた記憶探索課題(Okita, 1989)においても、視覚刺激を用いたと同様な分布を示す探索陰性電位が報告されている。このことから聴覚系においても、ヒトでは、外的並びに内的な聴覚表象の処理操作に、同様に関与しているのではないかと思われる。
註)Okita; Within-channel selection and event-related potentials during solective auditory attention, Psychophysiol, 26 (1989)127-139.
この節の冒頭で提起した「謎」の問題について整理してみよう。作曲ないし演奏とは、単なる音(素)を連続させることではなく、音(素)を何等かの法則=人間の思惟の運動・秩序などによって「組み変え」、「組み合せ」たものである。組み変えることは音(素)それ自体の存在性とは異なる存在性に移置されることを意味する。「総和以上のものである全体」とは、この「組み変えられた」新たな存在=個としての全体を指すと考えられる。
一方、視覚系においては、点→線→簡単なまとまり(三角、丸、四角)→それらが組み合わされた意味ある形象という風に複雑化されていく。同じように、聴覚系においても、単(純)音→協和音→メロディー→主題音楽というように“総合化”されていく。この諸要素(elements)を融合へと導く過程(プロセス)は、第一視覚野(V1, area 17)や第一聴覚野(AI, area 42)における視素や音素のニューロン群の電気反応的に刻印されて(impregnated, characterized)抽出される特徴づけないし「個(性)」の獲得した次元からスタートする。それがシナプスを替えるに従って、前にみたように、階層性の高いレベルの要素の反応ニューロン群に発展する。
これらのプロセスは電気生理学的に活動電位の測定や波形分析などで解明され、後皮質連合野内で生起する。充分には証明されていないが、logicalにreasoningしてみて、確かな現象として知られている。これらの抽出された「個(性)」-音楽や形質素-は或いは連続し、或いは断絶して、肯定と否定と統合をくり返して、Gestaltの世界が具象化される。このいわば感覚要素(的)ヒエラルキーのascending systemの中に扁桃体を主とする大脳辺縁系の活動であるpathos(情動)と新皮質内の言語野の活動産物であるlogos(言葉、思考)がmixされて(すなわち、皮質皮質間線維により相互にニューロン活動が影響し合って)より更に内容が深められて pathos とlogos が相互的に滲透した、新たな統合である pathologos の状態が作り出されてゆくが、このpathologosの関与の主題は後述されるので、ひとまず、傍らにこれを置くことにする。なお、付言すれば、脳の解剖生理学的研究が教えるところによれば、単純要素反応ニューロン群領域と複雑要素反応ニューロン群領域との間は相互に結合している。つまり皮質内の電気的興奮が一方向性でなく両方向性をもって伝達されることが証明されている。それを式で表すと、AI(V1)⇔belt(V2, V3) ⇔parabelt(V4, MT etc) ⇔post.assoc.cx⇔prefront.cx⇔mot.related areasとなる。
ところで、音楽において18世紀に1つ1つの音の響きを際立たせて演奏し、且つ幾つかの音をまとめてセンテンス=フレーズを形成させていく“語るような演奏”の奏法様式がとりあげられた。音の長さを短く切っていき、「個」としての音符の響きを浮き上がらせる奏法で、休符を用いて音を切ることによって(も)、例えば「ためいき、suspirationes」の表現として、聴く者の感情affectus, Gefuehlに訴える演奏方式である。これは明らかにギリシャ・ローマの古典文芸、およびその復活・再生であるルネサンス芸術を源とする修辞学(的手法)と深く結びつくことによって成立したものと考えられる。affectus をゆり動かすことによって真なるもの、美の根源への覚醒を促すべく用いられたと判断される奏法であり、バッハからモーツァルトの時代(17世紀半ばから18世紀にかけての時代)は「個」としての音素の響きが根源的なart / elementとされており、将にことば(言語)と感性(情動)-Pathologos-との同調に対応する音楽の形態が求められた。それがベートーヴェン、シューベルト、ショパン、さらにブラームスへと引きつがれていくのである。動物進化の立場からみて、音を切る発音(断続音/有節音の発声)は、喉頭の発達によりサルからヒトに進化する段階で可能になったもので、メロディー、もしくは何等かの連続する音の流れを意識的に中断することによって、話の流れにおける「単語」に対応する「個」の響きを明確に認識させるべく挿入させた休符の活用は将に人間的(サルからヒトへ進んだという意味において)なものである。
メモ:歌うように楽器を演奏して響かせる。カンタービレ。楽器に歌の技法を採り入れる。
Discrezione =tempo rubato
メモ:19/8討論による、子音・母音、運動野・発音器官の発達について考察するか
中世に遡るとはいえ、とりわけてルネサンス期の人々はこのような「人間的表現」として、つまり神の国とは異なる人間の世界における人間的世界の表現手法として、古典ギリシャ・ローマ以来のレトリックを導入して、言葉と音の響きを組み合わせることによって、思惟された世界を具体的・感性的響きのGestalt としてミメーシス(写し出)させるべくフマニスムス(人間)の音楽を響かせ、なおかつそれによって神的秩序=神・ロゴスによって法則づけられたコスモス内存在としての人間と、これを治めるロゴス・神との対話の場を作ろうとしたのであろう。
なお、このような音楽(芸術)における聴受と表現(演奏、作曲)およびこれと脳機能との関連について検討していく場合に、それも上記のような音楽史上の事柄としてこれを取り扱う場合にきわめて困難な問題が付随する。それは19世紀末、エジソンによる録音・蓄音機の発明以前のいわゆる音源が存在しないことである。無論、演奏・作曲の歴史を伝える楽譜は存在するが、言語の領域でひとつの単語に、時として多様な意味が内在し、その各れをとるかによってこの単語そのもののみならず、この単語が組み込まれた文章の全体の意味が多様に変化していくのと全く同様に、楽譜に内在する多様な響き・意味をどのように読み取るかによって或る音楽「作品」の伝えようとする意味やその作品の形態・構造は大きく変わって来る。従って、一般的聴受、つまり鑑賞という次元からすれば、或る「作品」は、これを聴く時に行われた「演奏」によって伝えられた意味・形態の域をまずは超え得ない。ひとつの「作品」でも異なる演奏者によってその形態が変化していくのはこのような事由によるが、このことこそが、上記のように史的検討を困難にさせるのである。或る「作品」、例えばBeethovenの「第九交響曲」とはどのような「作品」であるのか、についての検討は、この「作品」のGestaltが明確化され難いところに、その答えを出すことの困難さを介在させている、と言わねばならない。しかし、そうしたことを念頭に置いて記すならば、上にみた、言葉=演奏の形態は概略して20世紀の初頭までは続いていたと判断される。演奏の歴史は、しばしば人脈の系譜の歴史に一致するが、ここでもそれはあてはまる。一例をあげるなら、17世紀の演奏法を受け継いだ、18世紀前半のBachからMozartやBeethovenに流れ、さらにBeethovenの弟子のCzerny、あるいは同時代のウィーンの演奏法を身に付けていたと判断される19世紀のBrahmsとその弟子、もしくはBrahmsの友人でもあったレチェテツキがそれで、Chopinのノクターンに見事な「語る演奏」を遺したレチェテツキの録音にこの流れ・系譜を明らかに聴き取ることが出来る。同時に、このような演奏・表現法は20世紀の初頭に衰退し、新しい演奏・表現法がそれに代わったと考えられる。演奏・表現性の歴史にみられるこの変化・交代は、人間の精神の歴史のうえでの転換、いわゆるニーチェの「神の死」から実存哲学の抬頭の時期に、また一般の世界史では二つの大戦に向かう変動の時代に重なっている。人間の精神活動が脳機能の働きを反映しているのであれば、19世紀から20世紀にかけて人間の脳機能の働きそのものに、変化・断絶があったのであろうか。
一方、作曲の歴史は演奏の歴史と深い結び付を持っており、作曲家は、多分に伝統的なものとして、自分に与えられた、自分で利用し得る表現・演奏法に立脚しながら作曲をまとめていく。自分が伝達したい事柄が、どのように楽譜化されていれば伝達可能であるかを、作曲家は演奏の技法・表現法の点から検討していく、もしくはしていかざるを得ない立場に置かれている。無論、例えば19世紀前半、フランスのBerliozのように種々の新手法を模索し、発明し、これを利用することも出来るが、17・18世紀の作曲家はおおむね与えられた手法の利用によって作曲活動をしていた。このことは、作曲の基盤をなした修辞法の点からも首肯されるであろう。作曲・演奏上の、音楽の修辞は一定のパターン化された表現法の利用・組合せを根幹として行われたからである。当然のことながら、作曲のような能動的活動においてもなお、伝統との結びつきは強く、一般に想像されるような「個性的表現」が求められているわけではなく、Mozartのようないわゆる天才の作品でさえ、その根幹をなすのは既存の表現法、パターン化された手法の、彼なりの工夫であった。もっとも、このような点が「個性・天才性」を強調する方向でなされて来たこれまでの研究でさして問題にされて来なかったのも事実であるが、現在では音楽史そのものに対する洗い直しの作業の必要性が生じ、MozartやBeethovenの作品の中に流れ込んでいる伝統的表現方法、それも中北部ドイツの伝統が北ドイツの精神文化を経由して流れ込んだそれへの関心が高まっている。ウィーン古典派の位置づけそれ自体でさえ、近い将来にその在り方を大きく変えることになるであろう。
視点を変えるならば、このような伝統に立つ作曲と上の演奏・表現法の歴史・系譜とは合わさってひとつの軌跡を描いていることになる。MozartやBeethoven、Chopin、Brahms等の作品が語る演奏によって表現されていたばかりでなく、このことが告げるのは彼等の作品の中に語りかける表現様式がはっきりと認められることである。それも特に聴き手の情動に強く訴えかける緩徐な音の流れの中に好んでこの手法は用いられており、その際に、器楽曲が多くの場合、「テキストの無い歌」の形をとっているのが興味深いところである。この傾向は20世紀になっても認められ、Webern等の作品の中に休符もしくは音楽の流れの意識的中断による表現の深化の意図がみられる。加えて、先述の歴史の転換期にテキスト=言語の扱い方それ自体の中にも大きな変化がもたらされたことは改めて注意されてよいであろう。振り返って要約すれば、ルネサンス以来の歴史の中で、音楽の新しい表現手法として言葉を中心とした形態が打ち出されてきたのであるから、言葉化された人間の思惟・情動が、音楽によって具体的響き化されて来たのは当然の帰結であり、またそれ故にこそ上の世紀の転換点における言葉の響かせ方の変化は注目に値するのである。具体的に言えば、Monteverdi以来のRecitativo=朗誦の手法を発展させて、20世紀オーストリアのSchönbergが新たな朗誦としてSprechgesangと呼ばれる表現法を打出したのがそれである。これは、テキストを歌う場合に、楽譜上に正確な音程、音の位置を与えずに、大体の音高だけを示しておくもので(譜例?)、或る音が実際にどのような音高・響きで歌われるかは歌い手の情動・判断に任されている。古代ギリシア以来追求されて来た、音楽芸術の根幹をなす数比例に基づく調和を究極の美とする考え方とはまさしく正反対に、ここでは異常な、数比例に基づく音高からは捉え得ない響きが求められることになる。Schönbergは作曲家であると同時に、当時のドイツ芸術に顕著であった、表現の深化・意識の深層に根ざす表現を旨とした表現主義の画家としても優れた作品を遺しており、文字通りに、古代ギリシア以来の秩序づけられた法則づけられた宇宙・コスモスの法則に合致し得ない、人間の深層に横たわる各自・個としての存在の響きを音化してSprechgesang は成立したわけである。ニーチェの「神の死」に象徴される異様な事態の中で、人間の世界・コスモスを支えて来た秩序・法則が大戦によって破壊される歴史状況の中で、人間の存在の不安を映して新たに言葉は響いたのである。これもまた、人間の脳機能によって感知された何等かの、宇宙的不協和音の具体化であったと言わなければならない音楽上の現象である。もっとも、歴史における反比例の法則??という点からすれば、芸術の中核、ということは人間の精神活動の中核において数比例に基づく調和が求められたのは、地上的・人間世界における現実が非調和なものであったからであろう。これは実際にそうであった面と、例えばキリスト教によって地上的生活・世界の否定が説かれたことの両面によってもたらされた人間の脳機能を反映させるものであった。それに対して、神の死によって天上的調和が喪失されてこの方、非調和の現実が芸術の中核をなして作品は形成されるようになったが故の一帰結がSprechgesangであったと言い得る。なお、「神の死」はニーチェによって、いささか表現を濃くして説かれ有名になったが、Beethovenと同年の抒情詩人Hölderlinが同じ事柄を説いていることは改めて注目されて然るべきである。フランス革命に象徴される世界の転換の中で、Schönbergと同じくHölderlinが、あわせてBeethovenが世界の不協和音を聴き取って彼等の作品の中に響かせていたからである。その意味からすれば、語る演奏・表現方法そのものの中に、多かれ少なかれ不協和な状況を呈する人間の世界・コスモスは映し出されていたといわなければならないであろうし、改めて人間の精神活動、芸術という名のactiveな表現活動の根本的意味が、脳機能のpassiveな、外界認知の機能・働きとの関連の中で捉えられて来るべき問題を鮮明に映し出すものこそがこの語る演奏・表現様式であるといえる。おそらくは、中世に頂点をなしたキリスト教神学による現実世界の秩序付けがルネサンス以降の近代世界の中で衰退して行ったことの、これは盾のいまひとつの面を告げる歴史の流れである。
甚だ恣意的であるが、類似的興味から1)絵画史からも2)彫刻史からも例が挙げられよう。狭い私的な関心の範囲からみても、絵画史として:フィレンツェ派やベネツィア派のルネサンス期にみられる伝説画や肖像画中心の時代から、長い年月を経てセザンヌに代表される自然印象派の具象・写生の手法が生まれ、ついで個性的な画家としてムンク、ゴッホ、ピカソなどが19世紀から20世紀前半に活躍し、点や線を主体とする単純形象を根源的に求めたジャコメッテー(Giacometti)、モンドリアン(Mondrian)に連続している。また、日本の彫刻史に例をとれば、古代の素朴な土偶・人形は将に単純型の世界で、白鳳期の百済観音、菩薩半迦思惟像があり、美しい協和音とメロディーの響きは薬師寺の薬師如来三尊と東塔の美しさに感じられ、ついで鎌倉時代の、運慶、湛慶、快慶のアレグロ・ビバーチェ的金剛力士像となって円熟し、ずっと降って明治時代となり、ロダンの影響を受けた碌山や光太郎を過ぎて抽象的な凝集した近代彫刻の世界が訪れるのである。Spielのついでに万葉(人麻呂、赤人)→古今集(俊成)→新古今(定家)→宗祇の連歌→芭蕉、蕪村の俳諧→アララギ(子規、茂吉)という日本歌文学の譜系も同様に挙げられる。
晩年Verdi作のタイトルロールのFalstaffに慣って、人生は道化といくら達観してみても、当然至極のことであるが、上記の芸術史の展開と脳内ニューロン群の活動に成就される単純要素と複雑要素との関係を比較したり、類似させたり、ましてや、同一視点から考察しようとしているのではない。性質や次元の異なる事象の比較論は一見有益で面白そうであるが、意外と実(みのり)は少ないものである。
私がここで言いたいことは、我々人間がここまで築いて来た精神文化の創造と継続的開花はbasicallyにlogosとpathosと環境(自然と人間)に働きかける能動的行為(労働)の結合と醸成というヒトの脳の活動にみられる離散と集合、つまり自然がその法則に従って創った秩序ある物質世界の運動を反映したものであるということである。
さらにコミュニケーションの手段として言語をもつに至った動物、すなわち、人間の脳においては、以上の視覚および聴覚のGestalt機構が質的に発展したものとして、視覚的言語(文字)や聴覚的言語(話し言葉)を了解し、抽象化し、概念化するという、感覚的認識から抽象的認識に段階的に発展する概念化のGestalt機構が存在すると考えられる。
以上みてきたGestalt認識に関連する大脳皮質内の領域は後連合野内で互いに隣接しており、短い連合線維(弓状線維)によって密に結合されている。このようにGestalt認識の現代的解釈は、今や最近の進歩した脳科学の研究により科学的に解明することが出来るようになった。
このように言語機構が加わることにより、visual & auditory expressionの単純から複雑へ、または、具象化から抽象化を包含した脳機能は高次の段階へと発展する。すなわち、情動に関する扁桃体と強い結びつきをもつ、腹側側頭葉や側頭葉前方部(極部)は、これらの3 kinds of gestalt cortical areasに直接隣接してその腹側域にあり、この領域で皮質皮質間線維によりGestaltsとemotionの発現が結びつく。しかる後に、先にみたように、音楽(auditory-related)の affectusや絵画(visual-related)のaffectusは前頭連合野に結びついてmotorus affectusを生ぜしめるのである。
私の知る所では、ゲシュタルト認識の脳生理学を研究することが出来るまでのレベルに引き上げた“往時の”ゲシュタルト心理学は、歴史的にその源泉をみてみると、ロック(Locke, 1632-1704)、バークレイ(Berkeley, 1658-1753)、ヒューム(Hume, 1711-1776)などに始まるイギリスの経験論に求められる(ところの)19世紀の感覚主義や連合主義に対する反作用の一つとして位置づけられている。
そして留意すべきことは、ゲシュタルト心理学は本来現象学を源とするものであり、その直接的な哲学的先駆者はフッサールを主な代表とする現象学者たちであった。今日われわれは、上にみてきたように、その内容の精髄をくみとって脳科学の研究成果をもとに科学的に変革し、考察を加えることによって、脳と精神が如何に(how) 何処で(where).結びつくかという問題にチャレンジすることができる。脳と精神をつなぐ扉の鍵がここに隠されているのではないだろうか。
8]情動表現と脳内機構
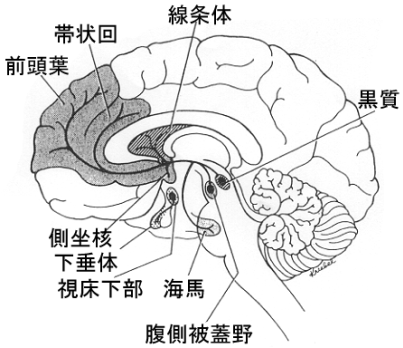
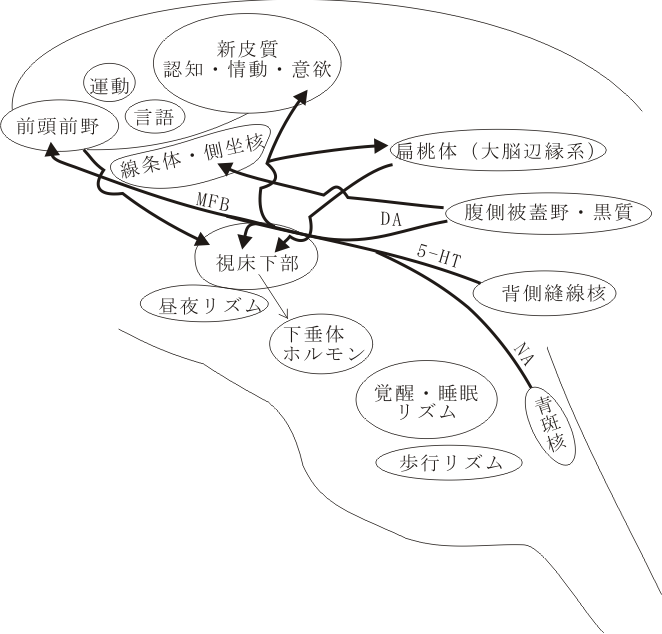
情動機能は認知機能とともに脳の高次神経の所産である主要な要素を構成している二大機能である。われわれは、外部環境および内部環境に関する情報のなかから生体にとって意味のある情報を認知し、過去の体験や記憶情報と照合して、その事象が自分にとってどのような意味をもつのか、報酬性(益、快情動)か嫌悪性(害、不快情動)かを判断している。記憶や情動は海馬や扁桃体を主たる構成領域成分とする大脳辺縁系の機能と密接に関係している。
海馬は原始皮質とよばれ、終脳の蓋板につづく半球内側面の部分が翼板の肥厚によって発生の早い時期に形成される大脳皮質の一部である。哺乳動物の海馬は、発生初期には脳梁の背側に位置しており、脳梁が背尾方に発達してくる時に同伴して次第に発達し、尾方では脳梁膨大の腹側で狭い小帯回となる。さらに腹側前方に進んで数珠玉を並べたような外観を呈する歯状回となり、また側脳室内に突出して海馬足(固有の海馬、アンモン角)を形成する。海馬の尾方発達に伴い吻側部は退化している。海馬はその発生初期から乳頭体と一部結合しているが、この結合部の内に割って入った形で新皮質の発達に伴って脳梁が膨大化し尾方に発達するので、この結合も伸張され脳弓とよばれるアーチが形成される。海馬は短期記憶や外界との関係で場所を認知する機能をもつ部位として注目されている。
一方、哺乳動物の扁桃核は、尾状核や被殻と同様に、半球胞の腹側壁が側脳室の内腔に隆起状に発達した神経節丘の後下部から生じる(マウスでは胎生13~14日頃)。側頭葉が形成されるにつれて神経節丘の腹側が前方に移動し、側脳室下角の前端の前上部に扁桃体が位置するようになる。ヒトの扁桃体は側頭葉前部の海馬旁回鈎(海馬旁回の前端が後外側に曲った部分)のすぐ下にみられる。一般に扁桃体と海馬は "関係が深い" と考えられているが、発生の過程を調べてみると、たまたま最終的に定着した位置が比較的接近しているが、両者は互いに独立分離して発達した構造物である。扁桃体(扁桃核とも言う)幾つかの亜域(亜核)に(内側核、外側核、基底外側核、中心核、皮質核など)区分されている。比較解剖学的にみて、嗅覚との関連で発達してきたと考えられている扁桃体(およびその周辺皮質部)の亜核間の関係が動物が高等になる程、複雑になっていることは興味深い。動物種間の相違はあるが、扁桃体は、皮質内側核群と基底外側核群とに大別される。基底外側核群(基底核と外側核)は動物が高等化するにつれて発達し、ヒトで著明である。一方、内側核・中心核および皮質核は逆にヒトで発達が悪い。なお、鳥類以下の原始線条体(または嗅線条体)は哺乳類の扁桃体に相同とされている。
大脳皮質との関係をみてみると、ネコやサルの所見から腹側側頭皮質、前頭葉眼窩面皮質、帯状回から嗅内野(entorhinal, 28野)や嗅周野(perirhinal, 35野)に、さらに28野と35野から海馬領野に投射することが明らかにされている。また、眼窩面皮質や嗅内野を含む側頭葉皮質と扁桃体との間にも相互間の結合がみられる。種々の感覚性の海馬への入力は嗅内野など海馬周辺皮質を介してみられるが、扁桃体への入力は間脳、中脳のいくつかの神経核(視床亜核、視床下部の腹内側核、黒質、縫線核など)や脳幹内の結合腕傍核、青斑核などから直接の投射が存在する。海馬と扁桃体は発生学的にも機能的にも異なる構造物であるが、この両構造物間の線維連絡は、少なくともサルの段階で明らかに存在する( )。現在、連合野を含む大脳皮質・海馬・扁桃体の特定領域と視床下部亜核との間の連絡についてのより詳細な研究がなされている。
ここで、現在明らかにされている海馬と扁桃体を含めた相互間の線維連絡の関係を簡略化して下図にまとめてみる。
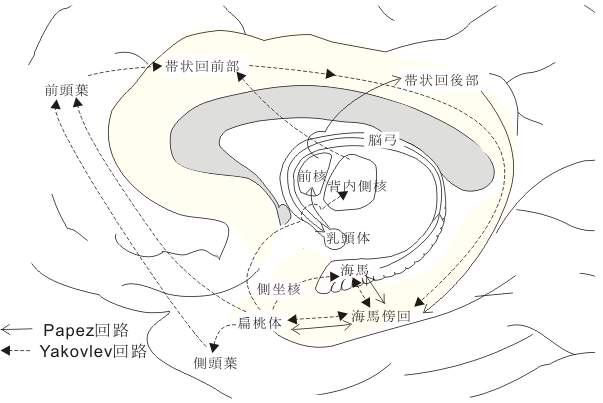
図:情動機構を説明するうえでのキーワードとなる構造物とそれらを連結する神経回路(Papez 回路、Yakovlev 回路)を示す模式図。
新皮質から海馬体への入力/情報の経路は必ず嗅内野・海馬台(ヒトでいう海馬旁回parahippocampal gyrus)を介している。これは連合線維(皮質-皮質間結合)のチェインである。皮質間の相互結合という点からみたとき、固有の海馬ないし海馬体は、"大脳の辺縁" の奥まった所にあって、視床下部を含めた脳幹部から入力される生存に必要な要素と連合野からの高度な情報を一時的にでも結びつける事により、一般記憶の記銘過程や空間記憶の保持に関連した作業に関与しているらしい。これを支持すると思われる行動・生理学的な証拠も提示されている。他方、扁桃体と大脳皮質との関係をみると、扁桃体は側頭葉極や下部側頭葉皮質や前頭葉の腹側部および眼窩面皮質など、感情・情緒に直接または間接的に関連する皮質と比較的強く相互に結合している。その上、扁桃核は味や臭いや自律神経系統の皮質下核と結合しており、亜核内での役割分担も示唆されており、皮質-扁桃核間の神経回路が働くことによって、賞罰、報酬などの意味づけ、動機づけなど形成される所と考えられる。このようにみてくると、哺乳動物の大脳辺縁系は大別して、記憶変換器としての「海馬体系」と、感情表出複合体としての「扁桃体系」とから構成されているとみなし得る。しかしながら、両者とも量質の差があるにしても、ともに大脳皮質と脳幹部からの入力(感覚系についていえば単に嗅覚系のみならずすべての種類のものが入ってくると考えられる)に制御ないし調節されており、この両系の働きを結びつけようとするならば、その主要な接点は嗅内野ないし海馬旁回(海馬台を含めて)にあると考えられる。
つぎに、扁桃体と情動記憶の関連について特記する(西条寿夫、小野武年を参照)。扁桃体を破壊したサルは、対象物の生物学的意味認知が障害され、以前恐れていたヘビやヒトに平気で近づくようになる(Kluver-Bucy症候群)。扁桃体が対象物、あるいは感覚刺激の生物学的意味認知に重要な役割を果たしていることを示唆している。扁桃体は、新奇刺激や感覚刺激の生物学的意味が変化ときに、それらの刺激の生物学的な意味を学習していく過程に関与している。ラットで、音を聞かせて、電気ショックを与えるという聴覚条件付け(恐怖条件付け)を行なうと、刺激条件(音)だけで血圧上昇やすくみ反応が起こるようになる。扁桃体が破壊されると、条件付け反応を用いた課題の学習が障害される。神経生理学的にみて、条件付け学習により扁桃体ニューロンが条件刺激にも応答するようになるからであると考えられる。
一般的に、強く印象を受けた出来事や情動的出来事に関する記憶は長く残りやすいことが知られている。このような情動の記憶増強効果は、扁桃体が記憶固定過程に促進的に作用していることによる。扁桃体は、記憶に重要な中隔-海馬体系における学習性のシナプス応答の変化を増強することが報告されている( )。さらに、海馬体苔状線維のテタヌス刺激により海馬体歯状回でLTP現象が観察されるが、苔状線維のテタヌス刺激時にさらに扁桃体内側部の電気刺激を同時に加えると、歯状回におけるLTPが増強されることが報告されている。
以下にまとめて考察するように、扁桃体は情動記憶の獲得や固定や保持にも関与している。扁桃体には、感覚条件刺激や強化刺激に誘発された快・不快感あるいは嫌悪性の痛覚や報酬性の味覚情報が入力している。ここでは、条件付けによりこれらの情報が同期的に入力し、長期増強などの機序により感覚線維-扁桃体ニューロン間のシナプス結合が強化され、扁桃体ニューロンは条件刺激に応答するようになる。また、扁桃体は、中隔-海馬体系のシナプス結合を強化し、扁桃体で得られた情動記憶を長期記憶として大脳皮質に移行させる。ヒトで、家族性鬱病の患者の鬱状態において、また健康人においても情動的な陳述記憶を思い浮かべる場合に、扁桃体で脳血流が増加していることがPETやfMRIを用いた研究により明らかにされている。
扁桃体には、味覚、嗅覚、内臓感覚、聴覚、視覚、体性感覚などあらゆる種類の感覚機能が直接的または間脳の視床核を介して間接的に入ってくる。この環境からの情報を将にそのまま感覚的に受容する(皮質を経由しない)生の粗なrapid sensationの他に、大脳皮質を経由していわば高次元で処理し、知覚し、認知した結果を時間的に少し遅れて伝達する適正な精密な情報が入ってくる。すなわち、扁桃体の基底外側核は、梨状前皮質、内嗅領皮質(28野)、帯状回(とくに24野)、側頭葉、前頭前野からの線維を受ける。以上の粗と精、原始的と識別的、低次と高次という2種の情報が扁桃核内で遭着する。そのことにより、環境に対して瞬間的、反射的に反応した生得的な生体反応は、成体の思慮深い知恵により、快か不快か、有益か有害かの判断に基づいて環境適応的に補正修正されるのである。ここで行われるこの価値判断の機能の結果は、主要な遠心性ルートである分界条(stria terminalis)を経由して中隔核、視床下部へ伝えられる。そして、小部分が腹側扁桃体遠心路(ventral pathway)により、視床下部に伝えられる。視床下部は食欲、性欲、水分代謝、自律機能、ホルモン調節(内分泌系を支配する脳下垂体の機能を促進又は抑制する)など生命維持に関わる重要な組織で外的または内的な誘因が存在するとき、この領域への興奮が来たとき周囲への働きかけの動因となり、有機体は行動(行為)を起こすことになる。扁桃体からの遠心性投射については、研究中であるが(湯浅ら)マウス胎生14~16日で核集団を形成中のニューロンから軸索が束化を形成してある特定の細胞群(LOT細胞)をmile stoneとして尾状核と視床の境界部に沿って背後部そして前方そして腹方へと走り、形成過程の視床下部領域に侵入する。そして腹内側核(満腹中枢)や外側核(摂食中枢)のニューロンとシナプスを作る。これらの領域にあるニューロンは、グルコースに反応すると共に、肥満因子オプシンに対して、腹内側核には促進的(positive) に、外側核には抑制的(negative)に反応する受容体(レセプター)が存在する。
自律神経系への入力にはすべての感覚があるが、最大のものは体性感覚(内臓感覚を含めて)である。ここで強調しておきたいことは、自律神経系への出口の問題である。その自律系反射は、視床下部を“最高中枢”として、ここから神経線維の連絡系として、延髄の迷走神経背側核や孤束核などに室傍核(PVN)から投射している他、液性(ホルモンや伝達物質)系として、下垂体への流れがある。この切り口は現時点で欠けている重要な点である。
嗅覚や味覚の刺激は直接扁桃体の皮質・内側核群(系統発生的に古い部分)に入力されており(川村、車田)、その情動への関与も無視できない。水棲動物では嗅覚も味覚も同じ化学物質が各感覚細胞に興奮を与える。聴覚は水圧や波動の感覚や一部味覚と同様に側線系がこれに当たっている。ある種の魚(ゴンズイ)では、味覚が非常によく発達して側線葉と呼ばれる膨らみが生じており、その前方は小脳原基と考えられる部分でそこには聴覚系の蝸牛神経核が発達している。イヌが道標として嗅覚系を用いて自分の尿の同定をしていることはよく知られているところである。なお、嗅覚や味覚は、とくに下等動物において、辺縁系/情動と強い関連をもっている。或る知的な女性から「香りに接したとき、様々なことを思い出したり、せつない、悲しい気持ちなったりする」という話を聞かされたことがある。ある味がその人にとって特定の体験をよびさます手段(キュー)になっていることはよくある。そして胸さわぐことがしばしばある。香道やaromatherapyはもっと高級な話で高次の認知機構に関連するものである。
ここで嗅覚と味覚の結ぶつきについて考えてみると、互いに補強しあう関係があるようにみえる。いわゆる「風味」といわれるもので香りと味の組合せである。感覚器から大脳皮質までの投射路をみると、味覚系は延髄から孤束核→視床VPM核→外側溝壁部皮質へ、嗅覚は嗅結節→扁桃体、視床MD核→前頭葉眼窩面皮質へと全く独立した別のルートである。最近 Dalton らはヒトを対象にサッカリンのような香りのない甘味物質を口に含むと、塩味や辛味の物質を口に含んだ場合と比べて、サクランボやアーモンドの独特の香りを嗅ぎ分ける能力が向上するという結果を得た(Nature Neurosci, 2000, May)。この組合せ料理は欧米では使用されることが多く、経験依存的な現象かもしれず小脳の扁桃がこの検知閾値の降下(感度が増すこと)に関係するらしいという。一方、視覚と聴覚との関連については、すでに前、[4]の章で皮質レベルの形態を中心に論じているが、現象面においても、ある種の音楽を聴いて、情景が視覚的に思い浮かぶことは多くの人達が経験するところである。一般に視・聴覚系は、嗅・味覚系にくらべて扁桃体や視床下部との結合は弱く、原始的な生の情動とは関連性が低いと言われている。尤も網膜からの視覚性入力と視床下部域での生体リズムの形成は別の視点からみて重要である。情動と感覚の階層性の問題は別に論ずべきテーマである。
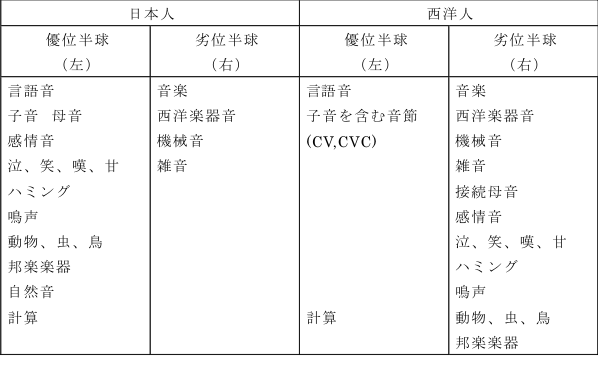
ここでクレッチメル(Ernst Kretschmer, 1888~1964)を登場させたい。チュービンゲン大学精神医学の教授で「敏感関係妄想、1918」と「体格と性格、1921」の著者として、またドイツ文化圏の文学的、芸術的そして哲学的素養をもった学者として有名である。これらの著書のなかでクレッチメルは、敏感関係妄想(Beziehungswahn)と彼が名づけたある種の妄想疾患は、感情移入に富んだ理解により、性格、環境、体験の間の関係を多面的に明らかにするという精神療法を施すことによって治癒させるという”多元的診断と治療、Mehrdimensionale Diagnostic und Therapie" の構想を呈示し、更にその診断の第一次元として体質の研究を行なった。そこで、内因性精神病、ことに分裂病圏と躁欝病圏での病像の生起を遺伝に規定された特定体質の心身両域にわたる表現として理解しようとする立場をとった。彼は、意識機能、欲求機能、情動の3つの機能を心的中枢機能と名づけ、それぞれを脳生理学的見地から考察している。そして彼は、嗅覚や味覚を”下等”(ヒエラルキーが低い)な感覚とみなし、内臓感覚、深部感覚なども含めて体全体に拡がっている情動や衝動に関係する感覚として「情動親近性感覚」と名づけ、生命感情と強く結びついているとした。これに対して、聴覚や視覚や触覚は表象や知覚として使われるもので、情動との結びつきは稀薄であるとし、「反映感覚」と名づけた。われわれの言葉を用いると、それぞれ、古皮質レベルおよび新皮質レベルの高次神経活動の反映ということになる。しかし、事は単純ではない。この問題は感覚の階層性と情動の階層性という両側面から、その相関について考察することを要求する。
クレッチメルは、情動性という概念を用いてその内容を、情動、感情、気分、気質と分けて以下のように定義した。すなわち、「比較的単純な個々の過程」を感情、「特に持続時間が短く範囲は限られているが強力な感情の経過」を情動、反対に感情状態が長時間に亘って全体として一様で漠然とした状態にある場合」を気分、そして最後に「或る個性全体をその情動性の全体的態度をもととして普遍的特徴的な形であらわしたもの」を気質と名づけ、それは個体の液体-神経性基質と関係が深いとしている。そして、体格と気質が内分泌器官および植物神経器官と関係が深く、情動と植物神経系、内分泌系は緊密な共同作業を行っているとして身体と精神の相関の問題を研究した。その成果として現れたものが著書「「体格と性格」」である。私は脳の高次機能の一つとしての情動の問題を、下等動物を含めて系統発生学的立場から、言語(ロゴス)機能を獲得したヒトの脳における情動(パトス)と結びついた精神活動に至るまでを生物学的に論じてみたい。ヒトの脳の聴覚連合皮質に焦点を合わせた活動について語ることは、音楽という芸術、すなわち、人間の精神活動について同時的に、無理なく一致させて語ることに他ならない。20世紀初頭に研究された上述のクレッチマーの成果から、80年経て神経科学の大きな果実を掌中にしているわれわれは、その精神を読みとり、精神医学を発展させなければならないと思う。
話は大きくなるが、摂食、性欲、渇、ストレス(グルココルチコイドとアドレナリン、副腎皮質と髄質)など恒常性(homeostasis)の問題も含めて自律神経系反応の問題が、視覚や聴覚が主になる大脳皮質の機能と関わりをもつ芸術の領域に参加したとき、activeな(運動性、能動性をもった)演出が、logosとPathosの共演として最高に表現されることであろう。ErosとPhagosが、ここにまたキーワードとして加わってくる。Phagosと Erosはどのように亢進し、また、抑制がかかるのか。視床下部に中枢性食欲および性欲に関する調節機構の第一の中枢が存在する。VMH, LHA, POAなどである。この中枢への末梢からのシグナルはニューロン網を介する神経性伝達と、循環血流を介する体液性伝達である。その上位にある高次の二次中枢は大脳皮質連合野(とくに前頭前野)と大脳辺縁系(とくに扁桃体)に在り、従って意志(willen)と感情(pathos)と理性(logos)の影響の下にある。以上が脳を唯物的に考察した結論である。
この他に、扁桃体→(側頭葉)→前頭葉→帯状回(その前部→後部)→海馬傍回→扁桃体および扁桃体→視床背内側核(MD核)→帯状回前部を連絡するYakovlev回路とよばれる情動・意欲に関わる神経回路網が模式化されて存在している。後述するように、中古皮質に属する帯状回は、意欲に関連した行為を行ったとき局所循環血流量が増加する皮質野として知られている領域で、サルの実験でも独自に新しく工夫した有用な行為を施行して報酬を得た時に活動する神経細胞(ニューロン)が存在する(丹治、Science)。
ここで特記しておきたいことがある。それは、アミン系のdiffuseな投射(DopamineやSerotonin)が上記の視床下部、大脳辺縁系(扁桃体、線条体、側坐核、帯状回)さらに前頭前野へ広範囲に存在することである(内側前脳束、MFB)。特に中脳の腹側被蓋野(VTA)から扁桃体、側坐核、帯状回に投射するドーパミン投射線維系は、A-10線維と略称され、新規(新奇)の刺激に(neugierigに)反応する快楽的情動/意欲(emotion, motivation)に関わるものとして関心がもたれている。側坐核はDA/D4R を有し、求心性投射として、venral subiculum および amygdala/BL &Cent nuclei から、それぞれ、脳弓、分界条を経由して、共に、同側性に興奮性入力が存在することが知られている。
ここでドーパミンニューロンの発生について記載しておきたい。ドーパミン産生細胞は中脳の脳室(完成された形で中脳水道)の腹側部神経上皮(基板)においてマウス胎生8-10、10-12日誕生のニューロンが分裂し、radial gliaに沿って腹側に移動、分化、更に接線方向線維(tangential fiber)に乗り変えて、以下の3つの部位に定着する。それらはA8:中脳網様体( midbrain reticular formation), A9:黒質緻密部(SNpc) およびA10:中脳腹側被蓋野(VTA)と名付けられている。A10ニューロンは上述した。A9ニューロンは運動の調節に関連する錐体外路系のもので、線条体(striatum, 尾状核と被殻から成る)に投射する系である(receptor はD1R, D2R)。この線条体は側脳室腹側部の神経上皮から発生(E14-16?)する外側大脳基底核隆起の細胞の集団から成るもので、それらの一部はアセチルコリン含有細胞、SPニューロン、GABAニューロンなどに分化する。Striatumへの最大の入力は大脳皮質からのもので主に運動関連皮質からくると言われているが、高等動物では、広いほとんどの全皮質領野(17野を除く)からの入力があり(前述)、身体全体のトータル的バランスを調節する機能がここにあるとされている。
以上、齧歯類で調べたドーパミン系の構成は、基本的には哺乳動物にもあてはまるといわれている。しかし両者間に進化の発達の程度を反映して、差違があることは当然で、その点も含めて、以下に少し詳しく範囲も広げて考察することにする。
上述したように齧歯類を用いた研究から、黒質緻密部と腹側被蓋野からのドーパミン系、それぞれA9とA10、の支配領域(ターゲット投射野)は異なるとされてきた。すなわち、A9線維は大脳基底核(線条体)へ、A10線維は大脳新皮質と辺縁系へ投射するというものである。しかし霊長類においては、このようなはっきりした区分はみられず、傾向はあるが、共に両者域に投射するように構成されている。ドーパミン細胞の軸索は無髄で活動電位の速度も遅く(1~5m/sec)、興奮性アミノ酸(グルタミン酸、アスパラギン酸)や抑制性アミノ酸(GABA)を伝達物質とする神経回路(投射)系のように情報を素早く伝えるという役割をもつ系ではなく、むしろ、標的ニューロンの活動を「調節」する役割を演じていると思われる。
ドーパミン線維は大脳新皮質に広く分布し、とくに前頭葉への投射はよく研究されている。ドーパミン線維の密度は前頭前野というよりも運動連合野(一次運動野を除く高次運動野)といわれる領野に高いといわれている。皮質へのドーパミン投射は運動の準備、指令を含む随意運動系の制御に関係しているが、線条体投射系は運動の開始と制御に関連している。
ドーパミン受容体(レセプター)は大別して2つ、すなわちD1ファミリー(D1, D5)とD2ファミリー(D2, D3, D4)がある。その主要なタイプは各々D1受容体とD2受容体である。また、澤口らのサルを用いた研究により、前頭葉の運動関連領野が担う随意運動のコントロールにはD1受容体の賦活が関わっていることが明らかにされている。前頭葉の運動関連域の高次神経活動とは自発性、能動性機能の発現に他ならない。表情、感情、意志の発現は、それが言語的表現にしろ、非言語的表現にしろ、調和的運動形態の調節から成立しており、この形態的基盤が、前頭葉におけるドーパミン終末とD1受容体にある。これが他の伝達物質、とくにGABAやグルタミン酸とシナプス結合を作ることにより作用し、投射線維と連合線維により海馬、帯状回、基底核、扁桃体、等々と連絡することにより、GABAやglutamateによる主要な情報伝達系をドーパミンが調節するという形で“精神・神経活動”が発揮されているのである。なお、ラットの線条体ではドーパミンがNMDA受容体による興奮性アミノ酸の作用を高めているというデータが得られている。ところで、精神分裂病にみられる陽性症状(幻覚や妄想)に脳内ドーパミン神経伝達の亢進が関与していると考えられてきた。臨床的事実としても、ドーパミン神経伝達を亢進させるアンフェタミン類(metamphetamine, d-amphetamine)とNMDA受容体を強力に遮断するフェンシクリジンをヒトに投与すると分裂病に類似の精神症状が惹き起こされることも知られている。
脳内ドーパミン作動性ニューロンの投射系
図の説明:脳における主要なドーパミン神経核は、線条体に投射している黒質緻密層、前頭及び帯状皮質、側坐核やその他の辺縁領域に投射している腹側被蓋野ならびに下垂体に対してドーパミン作動性の調節を行う視床下部の弓状核である。
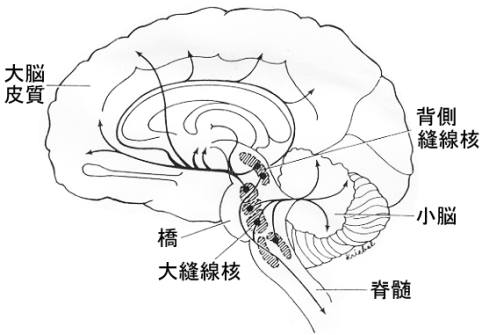
さらに中脳、橋、延髄の正中部に存在する縫線核(raphe nuclei)にはインドールアミン系のセロトニンが含まれ、これも中脳レベル(とくに背側縫線核)のものはカテコールアミン系(ドーパミン、アドレナリン)と一緒になって内側前頭束を作って視床下部領域(の外側部)をそこに分枝を与えながら上行し、大脳基底核(線条体、側坐核)、さらに大脳皮質(前頭前野、帯状回を含む広い領野)に到達する。セロトニンニューロン系は、トリプトファンから神経細胞内でトリプトファン水酸化酵素により合成されるセロトニン(serotonin, 5-hydroxytryptamine, 5-HT)を伝達物質とすニューロン系である。セロトニンはB1~B4と分類される脳幹内のニューロンで合成され、その軸索線維は脳幹、小脳、間脳、終脳に広く投射する。その様子は下の図に示されている。
脳内セロトニン作動性の投射
図の説明:脳でセロトニン作動性の主な核は、脳幹縫線核である。核は、やや拡大して描かれており、その広範な投射は著しく単純化してある。
セロトニン含有細胞群、すなわち縫線核の名称は次の如くである。すなわち、淡蒼縫線核(nucleus raphe pallidus; B1)、不確縫線核(nucleus raphe obscurs; B2)、大縫線核(nucleus raphe magnus),(nucleus paragigantocellularis lateralis;B3)、橋縫線核(nucleus raphe pontis; B5)、背側縫線核(nucleus raphe dorsalis; B6, B7)、正中縫線核・上中心核(nucleus raphe medianus・nucleus centralis superior; B8)、および内側毛帯にはさまれる橋網様被蓋核(nucleus tegmenti reticularis ponti; B9)である。
最近、セロトニンニューロンは、規則的な3~5 Hzの自発的スパイクを発することが明らかとなった(Aghajanian, 1985)。in vivoでの神経活動は覚醒レベルに依存し、この覚醒時に出現する自発的スパイクは、歩行運動、咀嚼運動、呼吸運動などのリズムが繰り返された時に発射頻度が増加し、注意を向ける行動(orientation)で抑制されると報告されている。
また、サルやヒトにおいて、ストレスや攻撃性行為の制御にセロトニンが関係していることが示されている。セロトニンの代謝物である5-hydroxylindol acetic acid (5-HIAA)の脳脊髄液中における量の低下とヒトの攻撃性、衝動行為、自殺行動との関連がみられることは以前より報告されていた。現在「低セロトニン症候群、low serotonin syndrom」といわれているものである(Linnoila & Vikkunen, 1992; J. Clin. Psych. 53:46-51)。遺伝的素因に対して育った個人的(家庭、友人、学校内の稀薄な人間関係など)および/または社会的(銀行の不祥事や警察の腐敗や凶悪犯罪など)環境からのストレスが加わることによって自殺行為、時にはいまわしき殺人行為(他殺行為)への引き金が引かれることにも発展しかねない。自殺者の脳の研究から、5-HTや5-HIAAの低下の他に、脳全体におけるセロトニントランスポーターの低下さらにシナプス後部の5-HT1Aレセプターや5-HT2Aレセプターの上昇(up-regulation)が、前頭眼窩野を含む腹側前頭葉に認められている。
以上見てきたように、辺縁系、前頭葉、液性の伝達物質、自律系を含む高次神経活動の創造的芸術まで指向するポジティブな面と、同時にそれが崩壊したときの精神異常を呈するに至る破壊的な側面とがあり、医学的にも教育的にも芸術・文化的にも子供の発達段階に合わせて直視して考えなければならない重要な問題である。
さらに附言すれば、DAを含むVTA からの遠心性線維は、MFBの中をNA, 5-HT 線維と一緒に通り、視床下部外側部(LH)のニューロンにシナプス結合を作りながら、通過し、側坐核(主にshell 部)にあるGABAニューロンにシナプスする(抑制性)。このGABAニューロンの軸索は、Meynert 核のAch 細胞(projecting to cortex)や腹側淡蒼球(ventral pallidum)のglutamate 細胞(projecting to LH) に抑制性のシナプスを作っているので、VTAからDA 刺激は、抑制を抑制する反応で、Cortex , LH の神経細胞を活性化activateする。性欲中枢に対するVTA fibers の作用機構もこの満腹中枢(LH)への作用と同様と思われる。以上は皮質下レベルの話しであったが、皮質レベルでも、17野に到達した視覚刺激は、VI→Temporal cortex→Amygdala→(Accumbence) → Prefrontal (ventral path) および VI→Parietal cortex→Prefrontal (dorsal path) のルートを介して前頭前野が賦活される。この、腹側被蓋野、側坐核、扁桃体、前頭前野を包括する視覚の世界の出来事は聴覚の世界の事象でも今後明らかにされる可能性が大いにある。
他方また、手網核(nucleus habenulae)、脚間核(nucleus interpeduncularis)、視床下部などには数種のペプチドニューロンが多数存在する。このようにpoint-to-point, rigid なものと soft, diffuse なものとから成るwiring patternとホルモン・ペプチドを含む液性因子により脳の機能、とくに意欲、情動の系は構成されているのである。
ここで、視覚および聴覚刺激に関連して、情動(一部認知を含めた)の問題を進化の立場から論じてみる。広く動物界でみられる現象に類似性があるかどうかを観てみると、単純(突発)音は、例えば、サルが「キー」とか「パン」とか発声して危険(警告)音として仲間に知らせる。この自己を危険から守り、そして逃避する際に発する音声は仲間同士の間のコミュニケーションの手段で、同種のサル(たとえばクモザル)達は、この音声に対して、かって自分自身が応答したものだけに選択的に反応するという。このように音という媒体を用いて、動物は発達した喉頭内により精緻な声帯器官を持つようになり、連続音/混合音/自然音を多用するようになる。そして、ヒトになると、色々な種類の声を出せるようになる。声は声帯で作られた空気の振動そのままではなく、咽頭を通って口腔と鼻腔につたえられ、そこで共鳴されて生まれるものである。声楽家の発声器官はまさに楽器そのものであるとわれわれはしばしば実感する。
また、別の例をあげると、蛾が鳥に襲われたときに、羽をパーンとひろげて”目玉模様”を突如として表出し、相手を一瞬驚かす。これも身の危険から逃避する際に動物が行動を表す単純化した色彩の表出(現)と言えよう。さらにまた、きわめて類似しているが、海中で群遊する小魚に下方向から大きな魚がこれを食せんと襲うときにみせる前者の反応がある。小魚の群は示し合わせたように(同時に)、瞬間的に、不連続的に、白い腹を反射によって輝かせて体をひねり、ひるがえして、方向を変えて逃亡する。
このような下等動物にみられる原始的行動に伴う音や色を用いた単純反射が人間にも本性として備わっていることは、原始社会における危険を知らせる鋭い音や、共同作業への単純な呼びかけ音(声)-「ヤッホー・エッサー」-などの使用に表れている。ゴリラやチンパンジーがコミュニケーションの手段として用いるドラミンゴの旋律は、黒人の踊りや歌にとり入れられたと思える程共感的で音楽的である。人間の場合、それが感性としてますます磨かれて感情・情緒の表現形態としてみられるのである。いわゆる、音を素材とした芸術、すなわち、音楽への原型である。これは、生活の糧を狩猟や漁労から得ていた古代人が、危険や害毒や悪魔から共同して身を守る手段として描いた洞窟の壁画にみられる原始美術の誕生とも軌を一にするものであろう。
ここで、以下に3つの問題を提起したい。第一に情動を今の時点で研究することの重要性について。脳の高次機能は大別して感覚・認知の機能と情動の機能に分けられるが、前者に比して後者の研究が世界的に立ち遅れている。そして、広い意味でも動物の行動、認知・認識の結果の表出の仕方に、情動が深く関わっていること。
第二に、あらゆる動物に情動機構は備わっているが、その脳の発展段階に応じて発現・表情発露が当然のことながら異なること。最も明瞭で本質的な相違または区別は、信号の信号化(パブロフ)の機構を基礎に物事の概念化、抽象的認識すなわち思考を可能にした言語を有している動物(すなわち人間)と有していない(そこまで脳が発達していない)動物とを分けて考える必要がある。と同時に、動物は環境との相互作用を通じて進化してより高次の機能を獲得して発展してきたものであるから、動物一般のもつ基本的、根元的な面から研究を進めることが出来る。また、そういう視点からの研究が必要である。このことは矛盾した2つの事象ではなく統一して考えることができる。
第三に、脳機能を能動的面からみた時の、いわゆる精神(=心)の正常と異常の問題である。この側面の科学的研究は最も立ち遅れており、現在良識ある研究者が苦心して陣地を構築中である。いわゆる「脳と心」の問題であり、「文化・芸術」の継承の問題であり、「精神異常」や「教育」をいかに科学的に捉えるかの問題である。
Mac Leanは、辺縁皮質およびそれと神経結合している皮質下組織を辺縁系(limbic system)と呼び、情動および内臓機能に関与する1つの機能系とする概念を提唱した。彼はまた、辺縁系が自律機能に密接に関連しているので、これを内臓脳(visceral brain)とも呼んだ。マクリーンのモデルにおける情動発現のセンターは広義の"海馬体"(海馬回、歯状回および扁桃体を含んだ領域)である。海馬体からの出力は視床下部を介して情動反応と自律反応を発現させるとした。
また、マクリーンは恒温動物の脳に3型のシステムから構成される階層性(ヒエラルキー)を考えた。いわゆる三位一体脳説(a hierarchy of three brains in one―a triune brain)と言われるもので、原始爬虫類脳(protoreptilian brain)、旧哺乳類脳 (paleomammalian brain) と新哺乳類脳(neomammalian brain)である。原始爬虫類脳は、脳幹、間脳、基底核よりなり、旧哺乳類脳は辺縁系に相当し、新哺乳類脳は新皮質をもっている。ここで、動物の古い脳の上に新しい脳が付加されるという進化の方向の道筋と人間の精神の構成の生物学的基盤が示されている。
原始爬虫類脳(reptilian)は、原始的な学習や記憶に基づいた、型にはまった行動を現わす。そして、この行動は個体維持と種族保存に基本的なものである。魚類や爬虫類では、
大脳基底核が最高の行動運動統合部位で大脳皮質は未発達である。これらの動物の行動反応は辺縁系と視床下部で開始され、そのパターンは柔軟性に乏しいステレオタイプ型である。パブロフ流に言えば、無条件反射の体系によって行動する段階である。
我流に解釈すれば、感覚入力としては、嗅覚のみ前脳の一部の嗅脳と原始的な辺縁脳に直接入るが、味覚、触・圧覚、平衡覚、視覚、聴覚などは、脊髄・脳幹レベルでの反射機構で処理され、能動的出力への仲介(または統合中枢)は原始的な大脳基底核と視床・視床下部であろう。ここで言う基底核は、運動系の他に原始的な側坐核や扁桃核を含んだ未発達段階の構造物と考えられる。
旧哺乳類脳(paleomammalian)は哺乳類において初めて発達した。この脳部位には情動の座があり、rigid/stereotypicな原始爬虫類脳の働きを、ある程度、柔軟に制御している。げっ歯類や下等哺乳類でも辺縁系と視床下部が反応発現の主体となっている。辺縁系と視床下部は行動発現の開始部位の一つである。すなわち、「情動過程」そのものに基づく行動発現と結びついている。この段階の動物は、人間と共通した情動の原型を明瞭にもっていると言えよう。また、パブロフの条件反射学説に従えば、第一信号系による反射活動の体系が備わる段階と言える。
今風に解釈すれば、古い皮質から一部に発生した爬虫類にみられる新皮質(general cortex)は領域的に広がり、大脳感覚野や側頭葉を主体とする後連合野からの辺縁系脳部位に対するコントロールが見られるようになる。
新哺乳類脳(neomammalian)は高等哺乳類においてみられる発達した。前脳の外被である新皮質は、外界環境因子をクールに分析し、高度の精神活動を行なう。霊長類になると、大脳皮質(とくに感覚野と連合野)、小脳、大脳基底核が著しく発達し、ここに行動発現に対する「認知過程」の関与が入ってくる。この過程で、道具の使用を学んだサルたちが表象能力(かって知覚されたことのある像を、その対象が現に感覚器官に与えられているという条件がない場合にも再生する反映能力)をもちはじめる。新皮質と辺縁系との相互連絡は動物が高等になるにつれて発達し、とくに側頭連合野・前頭前野と辺縁系構造物(海馬、扁桃体、視床下部を含む)との間の線維連絡は密になり、情動行動とその基盤はさらに複雑になり、質的にも異なったより発達したものとなる。ヒトでは新たに言語野が発達し(サルからヒトへの大脳皮質言語野の発達については、別に論じる)、言語は個体および集団間のコミュニケーション手段として大きな役割を果たすようになる。つまり、抽象概念を用いた思考が可能となり、情動状態を自省し、洞察できるようになる。言語はいわゆる第二信号系であり、特定の生物学的刺激および対象的経験にしばられている感覚信号系とは区別される。このように意思疎通の手段としての言語が形成された段階までくれば、人間による環境の反映、すなわち人間の意識の発生について語ることができる。それは、脳の発達の成果であると同時に、その脳を持った動物の社会生活の発展の産物でもある。後の項に述べる精神の発達と障害の考察は、この段階に達した脳の保有者、人間、を対象にしたときにのみ成立するものである。
さらに管見を述べれば、ここでは辺縁系に対する前頭前野の支配と「思考過程」の「情動状態」への関与が特徴的で、解剖学的にも連合野と視床下部・扁桃体との直接的または間接的(嗅内野を介して)結びつきは格段に強くなっている。
ところで、サカナに大脳辺縁系、とくに、扁桃核に相当する場所はどこにあるのだろうか。サカナ(メダカ)群集行動schooling behaviorといわれるものがあるが、終脳の特定の領域(ペプチドやTHやNkx2.1 geneの分布、線維の投射などから判断して恐らくマウスの扁桃体に相当する)を傷害すると群集行動ができなくなる。
トリの例で言うと、キメラ移植実験(ニワトリとウズラ)の結果、終脳の特定の核に鳥の種類に特有な鳴き声を規定している領野の存在が予測されている(竹内)。また鳥類の線条体は非常によく発達しており、HVC(High Vocal Center)といわれる上位線条体ないし新線条体(neostriatrum)で春期に神経細胞が毎年新しく生産されて当該領域が増大することがカナリヤで発見されている。“歌を覚える”とういことに関係があるらしい。この領域についての考察は、表現形態(運動系)との関連で大切なのであるが、現在のところ考察が充分になされていない。
以下にわれわれの教室で行われたたサカナとトリの情動発現の実験について、少し詳しく述べる。まず、硬骨魚類の終脳発生の特徴について一言しておく。
哺乳類をはじめとする他の脊椎動物の終脳は、神経管からの発生の過程で脳が側方に膨れて、側脳室を中心に背側から、海馬、大脳皮質、梨状葉の順で発達するpallium領域と、さらに腹側につづいて線条体、扁桃体、中隔が発達するsubpallium領域がみられる。硬骨魚類(メダカ)の場合、神経管の最背側のroof plateが伸長し、脳が外側に逆転した形で形成される。その結果、脳の表面がventricular zoneとなる。
坪川はメダカの終脳で、Nissl染色およびペプチド(CGRP, CCK, NPY, SS, SP)類やTH(カテコールアミン系)の免疫組織化学染色を行い、マウスにおけるそれらの分布と比較して、その類似性からextended amygdalaに相当する領域を想定した。つまりsubpalliumが背側、Dd- Dl region、に位置するところにventral striatum~amygdalaの近傍を推定した)この領域を両側性に破壊すると、恐らくメダカの情動障害のためschooling集合(又は群集)行動が出来なくなる。
話が少し大きくなるが、サカナの生態学的研究により、個体間の大きさの差、体長の同異のような因子に対する反応性の差に基づいて社会形態が形成されることが示されている。「群れ」(順位性群れ、同位性群れ)「縄張り」は、脳の”扁桃体”を主体とする辺縁系構造が有する誘引や反発などの価値評価の結果、それが行動に表現されたものであると考えられる。
次ぎに、話題がサカナからトリのレベルになると、単に、単純行動を示す集合行為などの面からではなく、個体自身が発する音声によってより高次のコミュニケーションが成立し、互いを引き寄せるという異なるタイプの社会行動がみられるようになる。
「鳥の鳴き声」については研究が進んでいる。キンカンチョウ、鶏を含む鳴禽類の研究はコミュニケーションにおいて、外来刺激に対して扁桃体の価値評価が大きな役割を果たすと同時に、種特異的な泣き声も重要であることを示している。鳥の鳴き声は、鳴管(synrix)という気管支のところにある器官の筋の緊張をXII運動神経(舌下)が支配することによって、音が決められている。また、このXII運動神経(舌下)は中脳にあるnuic(dm)の支配を受けており、さらにこの部分が、nurob(鳥の扁桃体相同部分)とHVC(鳥の運動皮質相同部分)の支配を受けていることが明らかにされている。この運動系のどの部分が種の特異性を司っているのか。竹内は種特異性を解析すべくニワトリ/ウズラのキメラを作成するという実験系を作った。
胎仔の脳の一部を微小手術により交換移植して、キメラをつくり、どちらの種特異的な行動が引き起こされるかを調べる方法である。この場合、雄の泣き声を生直後から実験を施行する7日目までテストステロンを投与して性分化を誘導した。ニワトリの場合は一鳴きに一回、ウズラの場合は3回の鳴動がみられることで、種特異性を確認した。孵化後2日目のニワトリの脳胞にウズラの終脳部、終脳+間脳部のみを移植したのでは、鶏の鳴き声のまま変化しなかったが、間脳+中脳、間脳+中脳+菱脳部を移植した場合にはウズラの鳴き声に変化した。
以上の結果から、鶏の鳴き声の種特異性を決めている領域は、先の鳴動運動系で間脳+中脳部分から発生するnurobの一部とnuic (dm)であることが示された。特にnuic (dm)は哺乳類の中脳網様体にあたる部分で、哺乳類では、ここが歩行など反復運動のパタンを形成するパタンジェネレーターであることからも、この部分が鳴き声のパタンを調節している可能性が考えられた。この研究により、社会行動の種特異性は扁桃体における価値評価という面のみならず、中脳網様体のMLR部も鳴き声のパタンの形成において、リズム発現と関連づけて重要であると考えられる。
次に終脳がさらに発達しているマウス(齧歯類)を用いて恐怖(不安)条件反射実験を行い、扁桃体が情動の発現に関与していることを明らかにした実験例を紹介する。
湯浅らは、マウスを用いて扁桃体からの重要な遠心路である分界条 stria terminalis の形成過程を調べ、胎生12日に視床下部に達していることを明らかにした。また、種々の神経伝達物質受容体を刺激あるいは抗精神病薬を投与することによって興奮する神経細胞の分布を最初期遺伝子の1つであるc-fos発現を指標として、情動にかかわる機能的神経回路を調べた。そしてfyn欠損マウスと正常マウスとの間で、扁桃体を中心に興奮神経細胞群の分布と強度に相違があることを明らかにした。すなわち、彼らはFyn欠損ならびに正常マウスにおいて、ブザー音と床の電気ショックを組み合わせて、音刺激に対して回避反応をとるような条件付けをした。そして、情動回路におけるc-fosの発現様式を検討し、過剰反応を示すFyn欠損マウスにおける神経回路機能の異常を見い出した。また、抗精神病薬の影響を検討した。次いで、恐怖条件づけにおいてチロシンリン酸化を受けるシナプス蛋白の同定と、Fyn欠損マウスにおけるチロシンリン酸化パターンの異常を検出した。
われわれは(大山、川村)遺伝子欠損マウス(knock-out mice)たとえばNkx2.1欠損マウスの解析で扁桃体や梨状葉皮質の形成不全や前交連線維の正中部における交叉不全、視床下部、前頭葉間結合の異常増強などの所見を認めた。このNkx2.1遺伝子は、ショウジョウバエのNK-2遺伝子と高い相同性を示すホメオボックス遺伝子でラット胎仔脳においては前脳特異的に発現することが報告されている(Guazzi et al., 1990; Price et al., 1992)。その標的遺伝子としては、thyroglobulin, thyrotropin (TSH) receptorなど甲状腺ホルモンの生合成に関わる遺伝子群が知られている(Civitareale et al., 1989; Kikkawa et al., 1990)。Nkx2.1-/-では、視床下部、下垂体、甲状腺、肺の形成不全が認められ、生後まもなく死亡してしまうので、残念ながら情動行動について観察できない。ヒトNkx2.1遺伝子ヘテロ(+/-)の患者の症例報告によると、精神的な発育に遅延、甲状腺刺激ホルモンの分泌に異常が認められており、いわゆるhaploinsufficiency(遺伝子1コピーでは不充分であること)を意味している可能性が考えられる(木村、私信)。
また川野、川村はPax-6変異体(mutant)ラットを解析して、視床から大脳皮質へ向かう線維が内包を通過することができずに、扁桃体核内に異常侵入するなど、limbic/emotionの異常を認めた。Pax-6は目の形成に重要な役割を持つ遺伝子で、その相同遺伝子はショウジョウバエ、ゼブラフィシュ、マウス、ラット、ヒトでいずれも目の形成に関係する。Pax-6を欠損する突然変異マウスとラットは、ホモ接合体が出生とともに死亡し、ヘテロ接合体では目の形成が未熟であるため、それぞれ小眼球マウス、小眼球ラットと呼ばれている。このPax-6は胎生期の正常脳では、嗅球、大脳皮質、腹側視床、前視蓋領域などに発現し、欠損するとそれらの部位に形成異常が観察される。小眼球ラットのホモ接合体では視床から皮質に向かう軸索の走行に異常が認められ、正常ラット胎仔のように腹側視床の外側部で神経束を作って内包を形成することなく、そのまま伸長して脳底部まで達してしまい、その後、扁桃体原基内に線維が迷入してしまう。
このように、われわれは種々の動物を用いて、情動の研究をし、貴重な情報を得ることができる。動物が高等になると、言語交流の萌芽がみられるサルの段階の情動・行動がある。ここでは、終脳が発達し、皮質間結合が増強され、連合領が広く発達する。そしてヒトに至って言語性皮質が誕生する。これらを理解する上でパブロフの条件反射における言語系と情動系の関連について研究する必要がある。
パブロフは、すべての動物が生得的にもつ、つまり遺伝子により規定された反射(無条件反射)に対して、大脳皮質にあり、実際に存在する情報が扱われる(条件反射の一次情報系)とヒトにのみある大脳皮質の言語性皮質が関与する条件反射の第二信号系の存在を示した。
[小生のパブロフ条件反射について再考のこと]
大脳皮質、とくに連合野が発達したサルの段階になると、価値判断ニューロンの存在する扁桃体の研究の外に、ある程度周りの状況を判断して行動する動物のパタン解析が実験的に可能になる。丸とか四角とかの区別認識が可能になり、短期の記憶と長期の記憶の関連などを研究することが出来るようになるからである。
それでは、ヒトの大脳皮質はサルに比べてどう違うか。それが脳の機能としてどのように反映されているか、パブロフの条件反射第2次信号系、言語系を機能し発揮できるヒトの脳はこの構造の複雑性、結合関係(回路網)の複雑性、発展性においてワニ・トカゲ(新皮質が初めて出現する;general cortex)(→トリ)→ネズミ→ネコ・イヌ→サルと進化して来たstepから飛躍的に発展する。ヒトの脳は、新皮質の幅、ひろがり、線維結合などの面において、サルに比べてはるかに高度である。組織学的にみて、神経細胞が密に分布し、錐体細胞の樹状突起がより高度に分化し、回路網が著しく発達している。ヒトは道具を使用し、労働を共にして生活し、コミュニケーション社会の文化を継続させる。ヒトの脳ではヒエラルキーの高い情動(emotion; Pathos)と言葉/言語(language; Logos)が結びつき、情報を処理する過程で分析力、解析力が格段に発達する。それをこの新皮質の構造が荷っている。ネコとサルとの比較は回路図の新皮質と辺縁系の結びつきの差の図ですでに示した(see above 図挿入のこと)。
神経進歩の図、川村!!!!
それ(ネコとサル)以上の差がサルとヒトでは確実に存在し、それがこの格段なる解析性、明晰性の差/発展であろう。その形態的単位(基盤)はcortexのもつ個々のmass[視覚領や聴覚領、また、各機能域内に各々の異なる機能を荷ったニューロン集団(mass)がある]が、ものすごく分化している。例えばトリでは、このmassが線条体(striatum)に相当し、カナリアなどsong birdでは、季節的に新線条体 (neostriatum) や原始線条体 (archistriatum) の RA(robust nucleus) と呼ばれる神経核のニューロンが増殖し、領域的に増大する。[これはヒトで云えば視床(Thalamus)のmassに相当する。]
またヒトのcortexは領域化の他に層状化にも特徴ある発展がみられ、これが高次脳機能をうけもつ分析性のbasisである。つまり分析的な知覚(諸)野と後連合野、更に、前頭前野への発展というように“上向”的にヒトのcortexをみたとき、下等動物には存在しないヒトのcortexの解析的性質(解析的仕事をすることが可能になる)が浮かび上がってくるのである。たとえばネコの段階ではmotorとsomatic sensory cortical areaは混じていて分かれていない。サル、ヒトとなりcortexの機能域は細かく分かれてくる。つまり解析的・分析的になってくる。それだけでは高次の機能は活動せず、それらを統合し、総括する(組みかえて束ねる)機能をもつ皮質領域がヒトになり出現してくる。この最高の領域(物質的言葉で云えばhighly organized pyramidal cells as well as non-pyramidal cells with efferent and afferent connectionsがみられる皮質部分)が連合野とくに前頭前野(Prefrontal cortex)に相当する。
なお付け加えておくと、1930年代に情動回路として提唱されていたPapezの回路(海馬→脳弓→乳頭体→視床前核→帯状回後部→海馬傍回→海馬)は現在の研究からみて記憶の回路に属するといわれる(小野)。情動の回路と言うべきものは、Yakovlev の回路がこれに当たるという。…小生作図、図14/精神医学テキスト/を加えること!!!
また、案として、このparagraph を前、e.g.,p53の辺に移動することを検討すべし!
最後にヒトの脳について、言語機能、概念形成能力を持った動物とそれらをもたない動物とで、情動の表現にどのような差があるか。Logos におけると同様にpathos においてもヒトと動物との間には質的に異なる性質のものが、脳の発達を反映するかたちで存在する。両者が融合したかたちで精神活動が具象化されてくる。Edward Munch の絵に表現されている「叫び」や「嫉妬」、オペラの中でもろもろに表現されている愛(タンホイザー)や嫉妬(トスカ、オテロ)の表現、さらに、J. S. Bachの受難曲に聴く崇高な響き。改めて言うまでもなく、これらはヒトの脳のみが創り出すことのできる芸術的な産物である。
メモ:自然に対する怖れ、Furcht (limbic level) und Ehrfurcht (association cortex level)
ここで一言、神経科、精神科の領域で、行われている音楽療法について述べてみたい。左程普及してはいないが、音楽が人の情操に訴え、記憶とくに連想を呼び覚ます力が神経系に及ぼすポジティブな効果をもたらすという考えに基づくものである。
恐らくこの背景にはピタゴラス学派や降ってガレノス?の思想がある。そこでは、自然は調和と数の原理によって構成された1つのコスモスとみなされ、倍音構造などの数量的音程関係によって構成される音楽は、ミクロコスモスである人間の精神・身体的な調和や均衡を再構成するための治療原理とみなされた。また、これの高次化された宗教的なものが、たとえば Bach の音楽で、身心の治療=人間に内在する宇宙の秩序の自覚・覚醒によって、その秩序の本源である聖なるもの=神へと人間の秩序=ロゴスを向けさせるべく構築されているのが数的な音楽であるとして捉えることもできよう。
この音楽の治療効果は古代から認識されていたらしく、旧約聖書にも記述されている(サムエル記上、16章23節)。若きダビデは痛みに苦しむイスラエルの初代の王、サウルのためにハープを演奏して、悪霊を取り去って安堵を与えたという話である。ギリシャ神話でもアポロ神は音楽と医療によく結びつけられている。
現在、コミュニケーション障害、気分障害、認知障害のみならず脳卒中や外傷の後遺症、筋ジストロフィー、パーキンソン病や痴呆を含む運動・知覚・記憶障害の患者を対象に音楽(絵画療法も然り)が利用されている。ここで用いられる音楽には、老人ホームで聞かれる演歌のカラオケの類から、精神的に高いレベルの、たとえば、バッハの音楽まで色々ある。知能が低下しつつある老人や精神疾患者には、クラシック音楽よりも大衆的な演歌や民謡が心に沁みて訴えるようである。メロディー、ハーモニーなどの音の連続的な調和のとれた美しさは、皮質レベルに働きかけて、神経症とか精神病に対して有用であると考えられる。精神異常を示す念慮 (Idee) や妄想 (Wahn) の形成は、認知機能、すなわち、ことばを手段とする機能と結びついている。それ故に、ことばを使わずに具体的なイメージも抽象的なイメージも伝えられるので、不安患者にはいわたりの曲をというように、人の情動のレベルと楽曲の情緒性を合わせるというアルトシューラーのいう治療法上の「同質の原理」の上に立って、精神的やすらぎの状態を効果的に誘導しうると考える。この音楽療法の場合は精神不安定者ないし一般の素人庶民を対象とするもので、芸術性を追求するための「ズレ」の概念を含んだ不協和音の効果的用法といった高いレベルの話ではない。
ともあれ、音楽の音響自体、人を快適にする性質を持っているという科学的知見も紹介されている。とくにクラッシク楽曲の音には、人に心身の調和をもたらす1 / fゆらぎと呼ばれる時間的変動が見出されている。
ここでちょっと煩雑になるが、武者によって研究されてきた1 / fゆらぎを理解する手だてとして、ゆらぎ解析の基本となる事項について、グラフの作成法(横軸と縦軸のとり方)と読み方を含めて解説しておきたい。すなわち周波数 (=f) 毎の成分に分解してそこに含まれる各々の強さ[振幅の2乗平均、すなわち各成分のパワー(1ヘルツの幅の中にあるパワーという意味で、パワー・スペクトル密度という)]を調べる。そしてスペクトルの周波数を対数グラフの横軸にとり、振幅の2乗、つまりパワー・スペクトル密度の値を縦軸にとる。このようなグラフを作るとパワー・スペクトル密度の周波数依存性が一目で分かる。
このグラフを説明する。パワー・スペクトルで表された形によって「ゆらぎ」の種類が異なる。横軸に平行な形は周波数とパワーが無関係なことを表す。「無秩序なゆらぎ」「白色ゆらぎ」と言い、すべての周波数成分を一様に含んでいる。両者間に相関関係が見られると右下がりの形となる。
相関が大になる程、パワー・スペクトル密度の価は、低周波数に向かって増加する。すなわちグラフ上でみると、スペクトルの傾向が経ってくる。これは、元のゆらぎに記憶効果があるということを示している。
風のそよぎとか打ち寄せる波などの自然現象には、ある時間単位で一定の規則性と適当な変化性が存在する。音楽音響の場合は、高さを表す周波数にも強さを表す音響パワーにも、1 / fゆらぎ特性が抽出される。あまりにも単純な繰り返しは退屈さを感じさせるし、突然の変化も心を動揺させるが、その点1 / fゆらぎは人に対して安らぎと楽しさを与える快適な波動といえるものと見なされている(下図)。
しかし、事はこのように単純ではないかもしれない。クラシック音楽すべてに当てはまるのだろうか? 1/f における「楽音」それ自体がどの楽器によるどのような「響き」によるのか不明である。 BachからMozart/Beethoven の時代には音楽の「治療」は「健康者」あるいは一部の上流階級に属する人達に対してのものであった。それに対して、19世紀末から20世紀にかけては、音楽の大衆化、すなわち、響きの大衆化が成立し、それが理解されるために必然的に「音楽の大衆レベルでのロゴス化」が起こってきている。この点、検討されて然るべき問題として残されている。
元来、自然界に存在する音は幅(ゆらぎとズレと雑音を含み)を持っており、この幅と正確さ・ゆらぎのなさとのバランスの上に音楽もまた成立しているといえよう。一切のゆらぎを排したものは単に人工的であり、また逆に幅のみによるものは、少なくともヨーロッパ系の音楽芸術の中では、真の美に到達し得ない故に非芸術的である。ルネサンス末/バロック初期にこの両者への関心が改めて高まり、ゆらぎつつも韻律法などを基礎として真の調和へと聴き手を導く、近代の修辞的音楽は成立し、以来、秩序と幅のある音楽が創造されてきたのである。ところで、あらゆる「楽音」には自然倍音が含まれている。その意味で「楽音」は抽象化された、高次化された自然音である。宇宙=自然が数によって抽象化されるのと同一の事柄として、楽音、純正5度は、抽象された自然であるといえる。「ゆらぎ」は低次の情動に属するもので、純正な響きになればなるほど、高次化・ロゴス化され、脳の高次機能に対応する。その意味では、一見奇異に聞こえるが、「ゆらぎ」・自然音が音楽療法に有効である場合、それは、ロゴス化された社会(コスモス)に対応し得ない人間存在の状況を映し出していることになる。
また、歩行障害者の改善(歩行の速度、調和、筋活動のパタンなど)を目指して、聴覚的なリズムを用いる「リズム聴覚刺激」の有用性はもっと強調されて然るべきである。このリズムのもつ脳の活性化の及ぼす効果は、音のリズムと運動のリズムとの同調に注意が向けられるべきで、小脳・中脳を中心とする脳幹レベルの機能賦活が大いに関係しているものと思われる。音楽のリズムは身体に快い刺激を与え、手足をリズムに合わせて動かしたり、リラックスした姿勢やスムーズな動作を起こさせる上で大いに助けとなる。
9]リズム/テンポとズレの生理学
ここで生体とそれをとりまく環境(自然、コスモス、地球)にみられる感覚と運動という範疇に存在する有体(ありてい)の姿としてのズレ/ゆらぎ/不完全さの問題について述べることにする。中脳(視蓋)レベルで、動く物体を視覚的に同定する小脳からの入力をうけたニューロンが見出されているが、網膜からの直接刺激による視覚性入力とのズレの補正をこの部位で行っている。聴覚性の動く音源の同定とズレの修正の問題については明らかではないが、コルチ氏(有毛感覚細胞)器官から蝸牛核、外側毛帯核、下丘外核・周囲核、上丘深層核などの諸神経核が、左右両側性に脳幹から大脳皮質まで興奮が上行する聴覚伝導系の中継核として知られているが、これら広範囲な脳幹内の諸核間の相互関連作用が関与していると思われる。臨床医学的に聴覚系機能を調べる検査法の一つに聴覚誘発電位(聴覚刺激に対する脳の電気活動をコンピューター加算して、頭皮上電極を用いて観察する)や事象関連電位(event related potentials, ERP, 外界の刺激に対して、被験者がなんらかの精神作業を行い、その際の刺激に対する一連の反応に伴って生じる一過性の電位変動、すなわち、認知活動に伴う脳内の神経活動、を頭皮上から電位として記録したもの)の測定があるが、有効なヒントが得られるかもしれない。音源(発音源)の振動数とその持続に関わるリズムの動きは異なる概念ではあるが、ズレ/ゆらぎの現象のもつ、不完全性(むしろ自然の物性を反映している)を連続面に投映したときの快ちよさ(An-genehm-bar-keit)は、視・聴覚系のリズムおよび一定の音組織の中に現れる音高(振動数)の中に、それも特に前者に、内在しているのかもしれない。
ところで、生物の活動には、決まった間隔をもって繰り返すリズムが存在することが知られている。昼夜リズムがその一つで、この生体リズム(biorhysm)が抑制/調整されずにそのままフリーラン(free-running、自由継続)すると、長い時間の方にずれて、1日のリズムが24時間+α(アルファ=±4~5)となることが知られている。ほぼ(circa)1日(dies)という意味でサーカディアンリズム(circadian rhythm, 概日リズム、日内変動ともいう)ともいわれている。このリズムの発振器(ジェネレーター)は体内にあるので体内時計といわれ、間脳の視床下部にある視交叉上核(SCN)を両側性に破壊するとリズムが消失することなどの実験結果からSCNがサーカディアンリズムの主時計、あるいはオッシレーターと解釈されている。24時間の周期で自転している地球上の生物(ゴキブリ、ショウジョーバエ、アプリジア、ラットなどで研究されている)は、24時間の周期リズムで活動している。しかし、外界の制約を取り除いた環境(光を遮断するなど)の状況下で気ままに生活させると、生体内の時計本来の周期(リズムの自由継続、free-running)を表わす。サーカディアンリズムの体内時計は、固有の周期が多少ずれていても、地球の自転周期(24時間)に同調/同期することができる。
サーカディアンリズムの遺伝子もショウジョウバエで発見されperと名づけられた(Bargielloら、1984)。地球上の生物は地球の自転による太陽の照明リズムの影響を受けている。単細胞生物でも細胞内に循環する物質代謝が組み合わさって同期(サイクル)が形成され、リズムを発現することが出来る。神経系のない生物、植物にも、たとえばねむの木の葉の開閉にみられるように、外界に昼夜のリズムがなくなった条件下でもサーカディアンリズムを示すという。哺乳類では網膜に入った光の興奮は脳内体内時計としてリズムのジェネレーターであるSCNに送られる。鳥類では松果体が光の直接照射を受け、ここがリズムに関係する。このように間脳における視交叉上核や松果体が体内時計としてのリズム機能を荷っている。ここは外界の光やホルモン(液性調節)に関係する領域である。
生体リズムとして、この他に性周期、体温、呼吸、循環の自律神経系などがあるが、意識とも関連のある重要なリズムの一つとして睡眠と覚醒のリズムをとりあげてみる。1個の細胞が生体リズムをもち、その集合である組織が同様にリズムを有し、組織が運動系と感覚系に分化し、進化の道筋を昇りつつ、神経系、脳(=中枢神経系の器官)に生体全体を調節する生体リズムのセンターが局在するようになると、その活動のエネルギー量は大したものとなる。大脳新皮質の発達していない魚類、両生類のレベルの眠りは「原始睡眠」と呼ばれていて、脳波上睡眠と定義できるような段階に達していないが、あらゆるレベルの(低次から高次に至る)精神神経活動(認知、判断、決定などの情報処理)をしている大脳皮質には「睡眠」という規則的に効率よく休息するという新しい機能が生まれてくる。
われわれは、脳波を記録することによって脳の活動状態を知ることができる。意識ないし覚醒/睡眠の変化を① 覚醒、② 徐波睡眠(ノンレム睡眠)、③ 逆説睡眠(レム睡眠)(註:急速眼球運動(rapid eye movement)の略号REM;がレム睡眠時に短時間にまとまって急速に動く。この時に夢を見ているという。)の3段階のリズムをポリグラフ上に見ることができる。因みに、このリズムは生体時計の中枢である視交叉上核(SCN)を破壊すると乱れることが知られている。
脳波による人の睡眠段階(表)
睡眠のリズムはどこでどのように生まれるのであろうか?昔から睡眠中枢の学説がある。Pavlov(パブロフ)によれば脳に特別な睡眠の中枢は存在しない。感覚刺激と唾液分泌を伴うその効果の強弱を観察して、彼は睡眠は神経細胞の抑制が大脳皮質全体から脳幹におよぶ現象と考えていた。この抑制は皮質のどこにでも発生し放散する。こう考えると睡眠の問題はリズムの形成とか変動に結び付けて考えるよりは、意識レベル(清明度)の変動の問題との関連が重要なものとなってくるように思われる。
Pavlov以外の学説としては、嗜眠性脳炎患者の脳の病理所見をもとにしたEconomo (1930) は、「睡眠と覚醒の中枢はそれぞれ視床下部の前部と後部にある」と考えた。また、モノアミン系ニューロンの分布をもとに、Jouvet (1964) は「徐波睡眠はセロトニンニューロンの働きで、逆説睡眠はノルアドレナリンニューロンの働きによる」という説を提唱した。この他、ベルギーのBremer (1935) はネコの脳幹の切断実験-上位離断脳 (cerveau isolé) と下位離断脳 (encéphale isolé) -を行い脳幹が睡眠と覚醒の変化にとって重要であると結論づけた。その後、脳幹網様体賦活系の説としてMagounおよびMoruzziにより包括された(1949,1954)。
とにかく、この章では、リズム、ゆらぎ、ズレ、テンポの問題を様々な角度から考察して、何かを探ろうと意図している。しばしば不確定な視点よりの冗漫な記載をすることを許されたい。
ギリシャの神殿の柱は平面図で長方形をとる神殿を囲んで四囲に立てられているが、長手(?)方向の神殿を挟む左右両側の柱は平行をとらず、地面(=床)に対して垂直でなく内側に傾斜して建てられ、その床もまた、平面としてではなく、中央部を高くして作られた。各れも、人間の視覚における錯覚(錯視、visual illusion)を修正するためのズレであり、このようなズレを含んだ建物を遠方から眺めた場合に、柱は垂直に、床は水平に「見える」ことになり、外観的に究極の美を追求したものであると説明されている。同じ印象は、平安神宮の新緑の頃の右近の橘と左近の桜にはさまれた大極殿(神殿)の朱塗りの柱を正面の離れたところから見たときの線の美しさにも通じている。また、一見ハーモニーの完全さを表象している左右対称性(シンメトリー)の問題に関して、精神医学者の島崎敏樹がエレガントに示しているように、写真を人工的に創った左右対称の人の顔貌や風景の作品は、凍りつくような冷たさと不自然さが醸ち出される。自然と人間の創造を融合させた芸術である造園にしても、天空の調和・数比例の美に関連づけられて定規とコンパスを用いてデザインしたようなベルサイユ宮殿の庭園の整然とした幾何学的美しさとは対照的に、池泉回遊式庭園の小川治兵衛作の無鄰菴には、どこからみても規則性をもったハーモニーの美はみられないにもかかわらず、ヒトの手によって作り出された「自然」に内在する秩序を直感させられる。さらに、この秩序を媒介として自然そのもの=宇宙の秩序を透視させて、飛び石の乱れでせせらぎを作り出した造園師の完成の音がひびいて来てわれわれの感動を呼ぶ。このような美的感覚にもとづく精神機能がどのように引き起こされるのかを解釈することはむずかしいが、少なくとも、聴覚系や視覚系レベルの高次機能だけを論じていては解決できないように思われる。言語野、情動系、空間認知系も関与している。考えるに、斜めに物体を傾けて眺めた時に生ずる立体(奥行き)的視覚認知に関係する頭頂連合野の一つであるPG野近辺 (PF, PFG, PG, Opt, IPS, POa, など) は、前に見たように視覚性および聴覚性のdorsal pathway or streamの重要な経路の中継点(いわばmile stone)と考えられている領野で、視空間認知と共にテンポのズレや音空間を含む音楽言語( Musiksprache )は恐らくこの領野およびここから前頭葉の背側運動前野へ投射する連合線維系に関連するものであろう。
視覚心理学において錯覚(illusion)の問題はよく研究されている。有名な錯視の図としては、たとえば、ドアが後方にあるようにみえるか、前方に見えるかという遠近法描画の錯覚を問題にしたGregoryの奥行知覚の図や、欠けた延長線分をGestalt的にみると三角形が浮き出るというvon der Heydtの図、また、同じ大きさの円であっても、周囲が小円で囲まれた時と大円で囲まれた場合とを較べてみたとき、前者の状況の時により大きい円としてみえるというEbbinghausの図などがある。
聴覚心理学において、この種の錯覚、すなわち錯聴についての心理学的研究については、余りないようである。[調べてみたい]が,あるとすれば、音の強さ、長さ、テンポ、音色、リズムなどにどのように関係してあるのだろうか?ここで、強いて、これらの錯視、すなわち、視覚のズレの図に、思いつくままに音楽的類似現象タイプの比較を試みてみよう。Gregoryの図はポリフォニーにおける声部毎の強弱による奥行きに、von der Heydtの図は音としての意味は連続しているが挿入されている休符の役割、たとえば、suspirationesためいき、の表現に、
Ebbinghausの錯視の図を錯聴化したもの
Ebbinghausの図は楽曲の中のおかれた位置により、前後の関係によって、その音符のもつ音の高低、長短、強弱が異なって感じられるという具合に、あるいは、或る音、たとえば上の譜例の♭●を意図的に低く、Ҧ●を高く演奏することによって、次ぎに置かれた音の響き・意味を、通常の場合とは異なって聴こえるようにすることによって、錯聴すなわち聴覚のズレを生ぜしめる、と解説するのは何如であろうか。もっとも、錯聴であるかどうかはともかくとしても、実際の演奏ではこうした音高を意識的に‘低目に’あるいは‘高目に’ずらすことが行われている。おそらく錯聴現象と結びついた‘快’の感情の喚起のためである。このように、「錯覚」というのは、現実にある感覚印象が歪められて知覚される正常の現象である。これとまぎらわしいのが、精神病理学的には異常な、特殊な主観的な知覚障害である「幻覚」である。Esquirolの古典的な定義に従えば「対象なき知覚、perception sans objet」である。しばしば、精神病者にみられる幻覚は、分裂病者にはなぜか幻聴が多い-というより殆ど然り-。多くは「声が聞こえる」と表現する。「頭の中で響く」ということもあり、「外から聞こえてくる」と表現されることもある。意識状態は清明であり、臨床的には意識混濁に伴って起こるいわゆる慢性アルコール中毒者のせん妄状態にみられる幻視と対比される。これは感情的確信によるものである。また幻聴の特殊型として考想化声Gedankenlautwerdenといって、自分の考えていることが声になり、それが人に知られてしまうと訴える患者もいる。幻覚はすべての感覚様態にみられるもので、幻聴、幻視の他に幻味、幻嗅、体感幻覚があるが、われわれがここで問題としている幻聴を神経科学的に考察すると、第一聴覚野(AI)の活動があって(ひきがねとなって)聴覚連合野が興奮するというのではなく、AIからの入力なしに、聴覚性言語野に“異常な”活動が惹起され、情動系と多少とも結びついた現象と見做すことが出来よう。仮説とも言えぬ想像であるが、この聴覚連合野にあるドーパミン投射をうけるニューロンのD2受容体(レセプター)の感受性の亢進などと結びついた研究を行うことにより精神疾患の解明が一層進展すると思われる。またしても、突飛な類推であるが、音を聴くことなしに楽譜を読んだだけで聴覚連合野が活動し音が響くという“訓練された音楽家”の聴覚機構と連がるものはないだろうか?
自然界における一存在である人間の脳(ヒトの脳だけではないだろうが)のもつ「ズレ」の修正機構は大脳辺縁系のもつ感情/情動の不断の起伏(ズレの反映)としても発現される。また、新皮質のうち最も発達したareaである(理性的規律性を本性として持っている)言語野と前頭前野とが互いに強く結合しあって、演奏、表出の過程で「ズレ」の「組みかえ補正」を行う。ここで言語リズムが問題となるが、多くの民族音楽はその民族がもつ固有の(特徴ある)テンポ、リズム、抑揚の規則性がある。古代ギリシャ語の朗誦にはリズムの長短に関わるアルシス、arsisとテーシス、thesis と呼ばれるものがあった。これは本来、足の上げ arsis と下げ thesis から来たものであり、これに強弱、高低のアクセントがどの程度に、どのように結びつくかは容易に決定し難いようである。arsis と thesis を弱強とする考えもあれば、逆に強弱とする考えもある。ほぼ同様の法則はラテン詩にもあてはまるが、ラテン詩・ラテン語の場合にはローマ時代・古典期のそれと、キリスト教的中世文化における場合とではまた判断が異なる。音楽においてarsis を弱拍、thesis を強拍とするのはほぼ19世紀以来の事柄に過ぎない。ともかく、このような歩行の用語を使用しているように脳幹・小脳の歩行運動開始系(後述)と大脳皮質・線条体・視床の運動調節系(前述)が関係して微妙なゆれが言語・音楽表現には含まれている。これが微小になればなる程表現形態としては崇高になり、大になれば美的になるという。このことは既に古典ギリシャにおいて、プラトンが指摘しており、最終的にはゆれ動きの少ないものによって良く整えられたコスモスの法則、創造の秩序に表されると言われている。これもまた芸術の本質的要素のひとつとして長くヨーロッパの歴史の中に生き続けた考えであった。因みに、ナチスドイツの軍隊の歩行はアルシスとテーシスの区別を持たない冷たい感じのする型式であった。
古代ギリシアには、あるいは古代ギリシア以降の歴史の中に、学問の在り方に二つの異なる体系・流れが存在したと言われる。ひとつは数理的なものであり、これは既存の「幅」を可能な限り排除することによって成り立つものであり、数学・物理学によく象徴されるであろう。一方の体系は、これとは逆に「幅」のある世界に依拠するものであり、例えば劇作法・文章作法に関する詩学等はこちらに入る。但し、この二つの体系・流れについては改めて次の二点を確認しておく必要があるといえるであろう。第一の点は、前・後各れの系もその本質においては共通の源泉に、つまり人間を含むコスモス(宇宙・世界)の根源的存在理由についての知的解明に立つということである。また第二の点は、数理的学問の系がさらに二分されて、数学という思惟に拠り立つ学問と、天文学・自然学のような観察に拠り立つものに分けられると言う点である。一般の歴史的区分からすれば、ギリシア以来の学問は、特に中世において明確であった自由七学芸に要約されるように数の学科と文学・修辞の学科に区分されるが、いまはこうした系統的な観点から離れて捉えるのが便利である。
学問の世界において、また芸術の世界において、ギリシア的特質として理性(的なもの)や光輝く明晰性が指摘されるが、これは盾の一面に過ぎない。他面においてギリシア人の非理性の解明(ドッズ他)や怖るべきもの、崇高なものへの畏怖の念(ガスリー他)は指摘されており、また併せて、ギリシア人の特質に、何等かの罪による穢れの念が強く働いていたことも明らかである。従って、ギリシア的なものとして理性的なものが浮かび上るとき、それはこうした非理性的なもの、畏れや穢れの念が強くあってのことであると言わなければならず、数的学問の源泉を成すに足るピュタゴラス(派)の知の体系に、数比例の秩序への注目と浄化の必要性とが併存していたことはその何よりの例証たるであろう。ヒトの認知の働きが脳機能全体の関連づけられた働きのなかで行われることによって例えば音楽と言語が相互的に深く結びついていることが明らかであるように、人間の認知した事柄の体系的表明、即ち学問や芸術の中に人間の多面性、存在の多様な響きは響き出ているのである。
一方、ギリシア的思惟による学の世界を捉えるために、コスモスの三層的構造を理解しておく必要があるであろう。普通に用いられるコスモスという呼称は世界ないし宇宙を指しているが、その世界・宇宙は一定の法則による支配を受けることによって秩序づけられていることを前提としている。その秩序づけについては別記(付論参照)されるが、基本的には人間の属する感覚界(現実世界)とそれを超えたところの宇宙と、さらには前二者を治め法則づける超越界とが問題の三層的コスモスである。もっともこの三層構造は現実(的)構造と人間の思惟によって捉えられたそれとが呼応しあいつつ在るもので、単に思惟によって捉えられたコスモスのみが問題であるとするのはあまりに論理的である。但し、秤の目盛りが後者に傾いていることもまた事実である。
このようなコスモスの全体的構造・在り様を集中的に説いたのはプラトンであるが、しかし彼の学説の特色である、彼以前に先立つ哲学諸士の手になる思索の結晶を総合的に捉えてコスモスに関しても一個の体系を提示したことは記憶されてよいであろう。そこでの根本的な構造として捉えられるコスモスの形姿は上の三層をなすものであるが、その場合にやや困難なのは、プラトンの用いたいわゆるイデアの語を、この三層のどこに位置づけるかである。いま便宜的にイデアを二分してイデア(もしくはイデアのイデア)とこのイデアのイデアに依拠するイデア界としてこれを捉えるならば、1)思惟におけるコスモスの三層のうち下の二層はイデア界であり、これを超える超越界のみがイデアの座所となる、かもしくは、2)三層の全体がイデア界であり、イデアはさらにこれに超絶している、ということになる。各れが正鵠を射ているかは決定し難いが、ここでは1)に従う。
イデアによるコスモスの把握の旨とするところは現実(感覚)界の無批判的容認に対する批判にある。この現実(感覚)界を超えて、理想的世界=イデアによるイデアールな世界に人間の在ること、即ち存在の根拠を求めることがその骨子であり、その場合のイデアールな世界こそ思惟によってよく整えられている筈のものである。それは先ず、哲人王の治める人間の世界と、別の表現をとれば、イデアに直接する叡智によって治められた世界と、それに冠する数比例に拠る宇宙と、さらにはこうした叡智や数比例の源泉・アルケーであるイデアという三層をなすことになるのである。プラトンの著作との対応では、哲人王によるそれは『国家』に、宇宙の構造は『ティマイオス』に、またイデア(とそれによる法則付け)は『法律/法律補遺』及び『パイドロス』にその範を求め得る。とはいえ、ここではこうした三層のコスモスとイデアの関係そのものについて問うことが重要なのではなく、このような構造を思惟し、認知したヒトの脳機能におけるイデアの体系が重要である。何故ならば、人間が日々に生き考え生活しているこの現実(感覚)界への埋没から世界を客観視し、これを論理的に体系づける、いわば世界の概念化がそれによって果たされるからである。またこうして客体化され、概念化されてひとつの論理体系としてイデア=コスモスが成立することによって、これを捉えたヒトの脳に内在してこの体系は存在することになったと同時に、しかしそれがあくまでも思惟されたものであることによって、これがヒトの脳機能によって発見=認知されたものであることによって、この体系づけられたイデア=コスモスはヒトの脳の外に存在するものであるとされた。ギリシア的思惟の特質における何か崇高なものへの畏怖の念は、人間に超えて在るこのイデア界を神的なもの、神聖なものとして感じ取ったが故のものであって、くり返せば、それはイデアの体系から切り離されて別個に人間を捉えたものではない。
プラトンの総合に関していま彼に先立つ二人の名前を挙げてみる。ヘラクレイトスとパルメニデスである。両哲人の世界観をみるに、一方は唯物的で、他方は唯心的である。ヘラクレイトスはこの現実世界、もしくはいわゆる存在しつつあるもの=自然(界)を成す万物の根源に火を見たことで知られている。後代に、アリストテレスが、先人によって捉えられた宇宙を構成する四つの元素、火・空気・水・土とこれらの元素を自然の中に見透した哲学者の名前を挙げているのはよく知られているが、ヘラクレイトスの見た火もしかし必ずしも彼だけがこれを見たのではない。むしろこれもギリシア的思惟のひとつとして理解されるのが正しいと思われるが、各れにもせよ宇宙を形成させつつ火は宇宙の中心に在り、自然=生命=存在を一定量燃焼させ、また一定量成生させる。一方、このような火による支配を一層抽象化させてヘラクレイトスが説いたのはロゴスによって万物は法則づけられている、もしくは法則づけられて在るのが然るべきであるいうことであった。ロゴスに聴き従って生きるのが賢明であるというのがそれであり、このロゴスをイデアに置換してみればプラトンの説いたところもその歴史性の一端を明らかにする。やがて、ヘラクレイトスの眺め捉えたこのようなコスモスの法則はストア哲学によって発展させられて現代に至るまで重要な働きすることになった。ストア的に捉えればロゴスはいわば息吹として万物に生命を与えこれを秩序づける。なればこそロゴスは神としても捉えられたのであった。
一方、パルメニデスは存在するもの、コスモスを四元素との関連においてではなく、これを思惟によって見透したのであった。思惟されるところのもの、それは在るものである。思惟されるところのものと在るところものは同一である、と訳されるパルメニデスの一断片が告げるのは人間の思惟が究極において関わり集中するのは在るところのものだということであり、言葉を代えればその集中においてこそ人間は在るところのものを見出しているのだ、と言うことになるであろう。無いものについて人間は思惟することが出来ない、と言う同じパルメニデスの断片もそのことをよく証しするであろう。この場合も、在るところのものがプラトンのイデアに対応することは改めるまでもないであろうし、ヘラクレイトスのロゴスに一致してこの場合の在るところのものもまた人間を超えて、人間の外に在る。
『ティマイオス』をはじめとする著作の中でプラトンは数比例を説いているが、この考えが何に拠るのかを特定するのは困難である。しばしば指摘されるのはピュタゴラス(派)との関係であるが、これはおそらくプラトンによって統合された彼に先立つ思想体系のひとつでしかなかったであろう。ピュタゴラス(派)によれば数比例の特色、重要な点は次のようなものとして捉え得る。まず一般に数比例とされるのはハルモニアαρμονιαのことであり、ハルモニアには多様な意味が含まれることを注意せねばならない。基本的には、在る点aと別の点bの二点間を結んだ弦(長)の分割としてハルモニアは捉えられ、ab間=1を2,3,4,…で分割した場合に、1:2、2:3のように簡潔な比例による分割によってハルモニア=調和した状態が形成されるのがその原理である。ab=1として表示される基音に対して2分割した場合の弦(長)が響かせるのは基音の1/2=オクターヴの音である。こうしてハルモニアは二つの点、二つの存在の比例=共鳴においてよく調和するもの、美しく共鳴するものを指すことになる故、ハルモニアは二音間の距離=音程としても捉えられる。このように見た場合に、或る音と別の音との距離関係である音程の組合せによって形成される音楽(作品)は、そもそも音程そのものがつまりはハルモニア=AとBという二つの存在の共鳴=ABによって示される数比例化された秩序であるところから明らかなように、デミウルゴスと呼ばれる創造神の手によって、数比例に従って秩序づけられたコスモスの形態を映し出すものとして捉えられることになった。ピュタゴラス(派)によれば、ハルモニアは純粋な形としては宇宙の音楽として先の三層のうちのコスモスの第二層、いわゆる宇宙=天体に響いているものであるから、実際には人間の感覚器官である聴覚には聴こえない。この天空の調和、天空のハルモニアを具体的に響かせるのが音楽(としてのハルモニア)であるということになる。ここに言う、具体的に響く音楽としてのハルモニアのうち、例えば2:3による音は基音に対して(オクターヴ上の)5度であるが、このハルモニア、つまりオクターヴ上の基音と5度は良く調和し、音程を表示する場合に用いられる数値では702セントになる。
これに対して現在一般に演奏されているピアノに用いられている5度の響きは種々の理由から702セントに手を加えて人工的な響きに変えた700セントである。この700セントの5度を用いると、オクターヴ/基音と5度の間にかすかなうなりと呼ばれる、二つの音が完全には溶け合わないにごりが生じる。このことは非常に重要であり、主として19世紀後半から一般に用いられるようになった700セントの5度による、微妙なにごりを含んだ音楽を今日では美しいと感じているのが人間の実状である。このにごりは、目下の論稿で問題にされているゆらぎとは関係のない単なる雑音性に過ぎない。それに対して、もしも思惟されるところのものが在るところのものであり、プラトンがくり返して説いたようにイデアにこそ真の美は求められるのであれば、ヒトの脳によって認知されたハルモニア、即ち秩序づけられたコスモスを映し出す音楽のハルモニアは、例えば702セントに象徴される、にごりのない響きであるのが望ましいであろうし、このにごりのない響きが前提されるときにのみ、音楽におけるゆらぎもまた可能になるといえるであろう。音楽によるいやしの問題について考える際に、このことは改めて注意されて然るべきである。
哲学においてのみならず、人間の感性・感受性に訴える芸術面でもギリシア人はこのようなハルモニアを追求し提示している。既述の建築における見かけと調和の問題もその一例であるが、ここでは他の例を取り上げてギリシア悲劇を簡単に検討する。ギリシアの、それも古典時代のアッティカとう時代的にも場所的にも制限された時空の中で、ギリシア人は悲劇という形をとって調和の問題を説いている。悲劇は、要するにふとしたことからもたらされる状況の逆転によって引き起こされるが、その根底に横たわるのは神(々)によって人間各自に与えられたロゴス=秩序もしくは各自の分を超えてはならないという考えである。分を超えることをギリシア人はヒュブリス=傲慢と呼んでこれを嫌ったが、別の見方をすればこれは汝自身を知れ、ということであり、あるいはロゴスに傾聴して生きるということである。悲劇の結末を通じて、ギリシア人は自己の在るべき姿を学んだわけであるが、その際に、悲劇的結末によって激しく衝き動かされた感情がパトスpathosであり、これをラテン語に訳したのがアフェクト(アフェクトゥス)affectusであることは既に指摘されている通りであるが、しかし悲劇が最終的に求めたものは、このパトス、衝き動かされる感情の揺れ動くことによって却って感情が昇華され明澄化されることであると考え得る。恐らく、このことをギリシア人はカタルシスと呼んだと判断されるが、カタルシスによって到達されるのは、哲学的思惟の働きによってなされるイデアの認知に一致する。現在では、ギリシアで悲劇が実際にどのような形で演じられ、どのように台詞が読まれ歌われていたかを知ることは出来ないが、韻律の形、詩として書かれた台詞を朗誦してその中に流れる、数比例に基づく拍節の秩序はこれを一例として、観衆の心、観衆の目に、悲劇を支配する神の摂理、神のロゴスを響かせ伝えて充分であったに違いない。ともかく、ギリシア悲劇が詩の形をとって、言葉の律動に即して書かれたとき、その台詞・言葉には数比例による宇宙観が映し出されて悲劇=文字・修辞の世界と哲学的省察の数の世界とが一体化されていたことは間違いない。因みに、ギリシア語で演劇は要するにドラマであるが、この動詞形ドラーンは行為・行動することである。ギリシア悲劇は従って、悲劇の形態をとることによって鮮やかに、舞台上に演じられた神の摂理・ロゴスによって法則づけられ秩序づけられたコスモスの一形態に他ならないといえるであろう。ギリシア人はこのような神の法則の演ぜられることを見物するための場所をテアトロンと呼び、テアトロン=見物席において観察されたところのものをテオーリア=認知されたもの=理論と呼んだのである。くり返せば、ここにおいて観察されたものと、自然において観察されたものとはその本質において同一の事柄に、すなわちコスモスの形態、もしくは在るものは何故ここにこのように在るのかという問いに対する答えに属する。
哲学も芸術(ドラマ)も数の学問も、等しく言語ないし文字によって記されたことは、既にギリシア語に、このような事柄を文字化する能力が備わっていたことを証する。文字化することは客観化ないし客体化することに他ならないが、この、言語にそなわる客観化の能力は、人間の思惟における客観化の能力・働きと正確に呼応していると言い得るであろう。思惟が言葉を要求し、言葉が思惟を具体化するからであって、例えばプラトンによって説かれたイデアは、既述もされたように一切の感覚的事物を超えて在る真なるもの・善なるもの・美なるものとして要約されるが、この真に在るものは人間を超えて実在していると同時に人間の思惟がこれを見い出し言語化したものに他ならない。しかもイデアという語が見ることと深く結びついたものであることに象徴されるように、何かを見ることは、既に、人間に向かい合って立っている何か在るものを見い出すこと、すなわち見識ることに他ならない。このことは、人間と向い合い、対峙しているその実在が、何等かの力、作用によって人間に働きかけてくるときに鮮明である。人間が言葉によって表わすものこそ、このような実在するものであり、当然のことながら、言葉を読み、目にするとき、人間はその言葉が表示しているこの実在を、言葉を通して見識るのである。このことを言い換えるならば、例えば人間が人間によって畏怖されるものをカミと呼んで以来、この言葉は人間に神を指し示し、また人間はそこに神を見、聴き、思い描いてカミという言葉において神と出会って来たのだと言うことになる。これをギリシア人の思惟に即して言えば、言葉=音・響きはシュンボロン=象徴である、と言うことになる。象徴・シュンボロンはつまり何かと何か別の或るものとが出会うことの意だからであり、またそれによって形成される第三の、それをこそ象徴と呼び慣わしているものを意味するからである。その意味でまさしくギリシア神話こそ、神と人間の畏怖の念とがどこか中空・中間において出会うことによって成立した象徴の世界に他ならず、これを一層抽象的に思惟し、いわば知の体系としてまとめ上げたのが哲学に他ならない。
このようなギリシア的思惟に照らして改めて言えば、哲学にしろ芸術にしろ、それあるが故にこれらのものが成立し存在している根本的な使命は、人間に虚しいもの・移りゆくもの、人間の所属しているこの現実(感覚)界から真に在るもの、真に美しいもの、イデアへと、またパルメニデスの説いた在るところのものへと、人間の心を向け変えさせることであったと言うことになろう。おそらくは、真に調和して在るもの、あるいはそのような調和=ハルモニアの源泉そのものとしてのアルケーである神(的なもの)こそ人間にとってその存在を支える最も尊いもの、最も美しいものである筈であり、もしも、いわゆるゆらぎによって快さが与えられ、人間の心・精神がいやされるのであれば、それは察するに、ゆらぎそのものがそこで目的とされるからではなく、ゆらぎを生起させているもの、真の美・一切の存在の源泉を成するもの=アルケーに人の心の目が開かれるからである。数比例に基づく音楽は改めるまでもなく、自然(界)の音・響きの中にさえ人はゆらぎを貫いて静かに響く深い調和を、アルケー=神(的なもの)を聴いて密やかな畏怖の念を呼び醒まされていることを忘れてはならないであろう。加えて、哲学・神学や芸術といった高次神経活動の所産、そのうちでも最も高い段階にあるもの、それこそが人間を人間として定義づける最も重要な事柄の知解に前頭前野が密接に関わっているのであれば、ゆらぎにおいてもまた、例えば人間(性)を高めたりこれを回復させたりするために、前頭前野への働きが喚起される種類のゆらぎが求められることになるであろう。ヒトの脳の機能との具体的関連はなお不詳のままに、17・18世紀の音楽の中でゆらぎが積極的に用いられたのも、このような、哲学的思索において探求された真なるもの、真なる調和の美へと人々の精神を促すためであり、そこではテンポ・ルバート(tempo rubato)としての時間のゆらぎや調和を強調するための不協和な響き、音程の微妙な「ズレ」によって作り出されるゆらぎ、響き等が活用された。これを要約すれば、アフェクト=パトスをゆり動かす音楽が求められたのだと言うことになるであろう。そこに、古典古代の学芸をその精神と共に復活再生させたルネサンス以来の音楽の意義は存在すると言い得る。なおこの点に関して詳しくは付論を参照。
われわれは生物学という「ズレ」を多く含む学問上の立場から、本来的機能として「ズレ」を有している脳髄を働かせて、美を対象とする芸術(音楽、絵画、彫刻、演劇など)の分野で、「ズレ」の修正の問題について論じている。「自然のもつゆれ動きつつある実体(実際の存在)」と「ゆれ動くことのない人間の思惟が捉えた自然に関する理論」との間の「ズレ」を視覚系においても、聴覚系においても「修正」して1つのGestaltの範囲内でおさめるという行為が美の追求であると言えないだろうか。人間の脳には、ゆらぎを許容し、「ズレ」を修正する機構が備わっているものと思われる。
身体の動き(体性運動感覚)や形象や音の動きを感覚的に捉える(視覚性運動visuomotor; 聴覚性運動audiomotor)機構やリズムの形成機構などには、歩行とも関連した小脳・中脳および脳幹の領域が関係するというデータが得られている。ネコの脳幹の様々なレベルを切断して自発歩行の開始機構(歩行リズムジェネレーター)の研究を行った結果(森ら、有働ら、1979 in B.Res., 1980 in JNP)、視床下核(nucleus subthalamicus, SUB, この核および近傍の領域をまとめて視床下歩行野、subthalamic locomotor region, SLR,という)からの刺激が中脳にある楔状核(nucleus cuneiformis, NCF,別名中脳歩行誘発野、midbrain locomotion region ,MLR)に加えられることが明らかになった。すなわち、SLRが自発歩行の開始機構に関与しており、その作働を待って歩行運動が駆動され、歩行が実現される。また、この中脳の歩行領域に対して視床下部内の主たる性中枢であるpreoptic are (POA) からの入力も存在する。因みに、POA は情動の中枢である扁桃体にも投射している。
筆者は歩行を合目的に継続する中枢機構は小脳にあり、恐らくは大脳基底核が大きく関与しているのであろうと考えている。これに関連して、動く光源や音源の位置の同定には小脳-視蓋系が関与し、網膜から視蓋に入る視覚情報がズレの補正に関与することが明らかにされている(サル、ネコ)。他方では、静的な形や音の注視(attention)にはサルの弓状溝前方の主溝を中心とした領域に存在する前頭葉のニューロン群が関与しており、後連合野から視覚、聴覚(更に体性知覚)の高度に分析された入力をうけている。この他、情動系や自律系の入力もこの領域に送られてくる。こうしてみると、前頭前野ニューロンは視覚系や聴覚系の活動ないし情報の単なるズレの修正というよりは、情動や内部環境および外部環境からの影響を含めた様々な要因を統括して、行動的意味へ変換(connotational conversion)して記憶、意味づけを伴う修正を加えていると解釈することができる。
リズム、テンポの形成は、脳幹、小脳、中脳、視床下部、線条体、大脳皮質と脳全体のあらゆるレベルでこれに関係し、各レベルでそれぞれ特徴をもっている。そしてこれらが全体として総合されたものが運動系にconvert/conveyし、表現される。1~2才の赤子、幼児が音の調べに応じてリズミカルに全身を動かし、ジャズのメッカNew OrleansのFrench Quarter通りで黒人の子供が手と足を巧みに動かして上手なテンポ/リズムで踊っている姿をみると、皮質下レベルの本能的なものを感じさせる迫力がある。前に述べたように中脳には歩行開始(ジェネレータ)の領域がある。そこから脳幹網様体の中を幾つかのシナプスを替えて、脊髄の運動ニューロンに投射し、これを興奮させる。視床下部には大まかな昼夜リズムのセンターがあり、線条体には、純運動系の他に連合(認知)系、辺縁(情動)系をも包含した調和的、統合的バランス感覚のリズム調節機構がある。また、脳幹から身体感覚、聴覚、視覚、平衡覚、深部感覚などほとんどすべての感覚を受け入れている小脳の機能は無視することができない。考えてみると、リズムというのは音のみに限定して関連づけられるものではなく、光のフラッシュや踊りのステップなど、感覚や運動一般における、その開始と中止(ないし休止、断絶)の間のインターバル(時間の流れ)を言うのである。ヒトで、機能的核磁気共鳴断層法 (functional magnetic resonance imaging, f MRI)を用いて出来事(events)の連続性と間歇性という脳機能を調べた坂井、彦坂ら(1999) によれば、小脳の後葉半球部は運動学習や仮想運動思考(頭の中でテニスのプレイをイメージするなど、virtual action or movement )や発声言語などの他に、ランダムに切られた連続音に反応するというようなリズムの形成過程にも関わっているという。
ここで、この文脈の中心となる小脳に関する神経連絡回路網の形成を中心とした(主としてヒトにおける)脳の発達について記述し、続いて、大脳との関連(いわゆる小脳・大脳関連ループ)についても若干述べることにする(精神医学テキスト、小脳の神経学を参照のこと)。小脳は菱脳胞の背側部の翼板(alar plate)と呼ばれる領域から発生する(但し、少なくとも一部は中脳胞後部の翼板も関与することが知られている)。胎生中期にこの領域が発達し、左右の菱脳唇→正中部で融合して小脳板が形成され、その部位のニューロンが誕生し、移動し、分化して大型のプルキンエ細胞(下オリーブ核から登上線維入力を受ける)や小型の顆粒細胞(橋核、脊髄、網様体その他の下オリーブ核以外の脳幹諸核から苔状線維入力を受ける)、その他の諸細胞(星状細胞、ゴルチ細胞、バスケット細胞など)が所定の位置に定着する。発達中にみられる小脳組織内では、これらの細胞間の連絡が完成し、同時に、脊髄(背側及び腹側脊髄小脳路,DSCT & VSCT)や脳幹の小脳前核からの線維の軸索終末との間にシナプス結合を形成し、同時にプルキンエ細胞からの脳幹および間脳へ向かう出力線維(大部分は小脳核を介して間接的に、一部前庭核へ直接的に)が、発達中の神経細胞へ送られてシナプス結合を形成し、中枢神経系の構成の枠組みがつくられる。ところで、中枢神経系組織は発生学的に神経管neural tubeから、その尾部から吻側へ向かって順次発達する。小脳や下オリーブ核へ投射する脊髄ニューロンの軸索は、DSCT, VSCT および SOT(spinoolivary tract)などの神経路を形成して、下小脳脚に入り、標的に向かって進行する。と同時に、これらの脊髄内ニューロンは、歩行リズムに関連する神経機構(上述)や前庭・平衡・姿勢・運動機能関連領域からの下行性神経路を介した入力を受ける準備をする。同様の出来事が、次いで延髄レベルで、そして橋のレベルで、小脳の発達に平行して起こる。ところで、延髄にある下オリーブ核のニューロンも橋にある橋核ニューロンも発達中の橋・延髄レベルの、すなわち後脳胞の、翼板から生じた神経細胞が腹側方向に移動(migrate)して定着・分化したもので、背側方向に移動した小脳内神経細胞とは同一ないし、連続した起源を有するものである(Ono and Kawamura, 1989,1990)。下オリーブ核へは脊髄や脳幹の種々の神経核(主として赤核、その他に上丘や三叉神経核など)から入力があり、大脳皮質運動領からも僅かながら存在する。橋核への主たる入力は大脳皮質からで、その全新皮質域からの入力を受ける。そして、それぞれ、下小脳路と中小脳路を通って情報/入力は小脳の、それぞれ、プルキンエ細胞と顆粒細胞に伝えられる。より詳しい内容については、専門的になるが、他書(Ito, …,川村,小脳の神経学、医学書院他)を参照されたい。
小脳には随意性および不随意性の運動性の入力の他に、体性感覚、視覚、聴覚を含めて多種の感覚の入力があり、苔状線維と登上線維を介しての入力の処理が行われている。小脳におけるLTP(long term potentiation)の研究は学習機構に関連して詳しく調べられている。これらの運動抑制および感覚性認知の両面にわたる機能の処理過程は、外界との関連において運動と感覚の事象を円滑に協調的にまとめ、活動をスムーズならしめるというタイミングの制御(コントロール)に関与している。つまり、運動、知覚認知のタイミングのコントロールの学習をしているということになる。このように考えてくると、小脳は単に自転車に乗るとか、スキーを滑るとかいう運動学習に関与しているのみならず、感覚性の知覚認識に関する個々の要素(エレメント)を円滑(スムーズ)に結びつけるという“メンタルな機能”の協調的活動の遂行に関与していると考えてよいだろう。この原始的/自動的/無意識的/不随意的な協調、つまり、もともと下等動物レベルから、認められる“個々の要素を関係づける”という協調は、脊髄・小脳前葉・中脳を中心とした系列に包含される。他方、新奇な外的刺激条件を受容して識別的に、意図的に、随意的に行われる円滑な協調は、大脳新皮質・小脳後葉・間脳・新線条体で構成される発生学的にも新しい系が関与していると考えることができよう。以上の二つのシステムを結びつけ統一する形態的な構成として、小脳・大脳関連ループ系の存在をみることが出来る。
この新しい小脳・大脳ループは前脳の発達を待って成熟したものとなる。そして、小脳、基底核、脳幹網様体、脊髄の間を多シンプス的に結びつけている“錐体外路系”を主とした運動系の上に幹線道路としての意識的、随意的運動を司る皮質脊髄路が新しい“錐体路系”として完成する。
図A 最大に簡略化した新旧小脳・大脳関連グループの図
上記の小脳における運動・感覚のタイミングの制御に関連して、小脳が有する(であろう)認知機能(注意、記憶、言語、感覚、刺激受容)に関係するリズムの処理ないし時間情報の処理について考える。この研究は坂井、彦坂ら(1999他にもあり…)により行われた。すなわち、かれらはヒトに7つの音から成る連続リズムを聞かせて、短期記憶させたときの脳内活動をf MRIを用いて研究した。
彼らによればリズムはインターバル比率に依存しており、その構成の比率により二通りのパタンがあるという。すなわち
①左側の運動前野、頭頂葉と右側の小脳前葉半球部は1: 2: 4及び1: 2: 3のリズムに反応し、
②右側の前頭前野、運動前野、頭頂葉及び両側の小脳後葉半球部は1: 2.5: 3.5のリズムに反応する(ニューロン群がある)。彼らは、①を音が存在するかしないのかの2値化を表す(metrical rhythm)階層的な表象構造(hierarchial representation)をもつ処理の系とし、②を一つ一つの時間的間隔を順次記憶する(non-metrical rhythm)直列処理の系(serial chain representation)と区別した。この解釈はかなりむずかしいが、①はわれわれの生活(環境)に自然な馴染みのあるリズムであり、②は、日常にはない一瞬戸惑いを感じさせるリズムである。この“新奇”なリズムは小脳後葉で形成され、それが固定される(馴染み落着く)と小脳前葉の活動となるという。
以上の結果は、
memo:協和、不協和を聴き分ける機構と関連するかもしれず、この項は極めて重要である。
となり、聴覚からの入力によって運動系に刺激が加わり、テンポ/リズムがヒトの身体内に発現すると考えられる。発生学的にみると、線条体/間脳→脳幹→脊髄のルートが始めに、原始的な踊りとして歩行機構と結びついた形で発生し、辺縁系/情動系により駆動され、感情移入された情動の昇華の源が見出されることになる。実際尾状核の視覚性記憶ニューロンは、運動の報酬スケジュールによって強く修飾(modulate)されることが示されている(川越、彦坂ら、1998)。かれらは、サルを用いて視野の中心部を注視させておいて、光を視野の上、下、左、右にランダムに点灯させて眼球運動を起こさせ、大脳基底核ニューロンの活動を記録した。この時、ある特定の方向に眼球を動かした(外眼筋を自己の意志により活動させた)時のみ甘いジュースという報酬が与えられる。この実験のパラダイムは、視覚系が空間を認知し、価値判断や動機(motivation)とリンクさせて運動系に変換させる時、どのように大脳基底核のニューロンが関与しているかを調べようとしたものである。この領域は、扁桃体から情動系の入力が、またさらに新奇物体に対する意欲的行為に関与する中脳の黒質からのドーパミン入力が、終止する場所である。実験結果は、この領域のニューロンは、サルが、形象を自己にとって意味のある空間内に認知し、その方向にむかって意欲的に行動を開始したときに活動しており、基底核(実験として尾状核を用いた)は報酬を期待することによって動機づけを評価し、その結果眼球を動かすという運動系の決定に関与していることを示している。なお、専門的になるが、線条体・視床・大脳皮質の並列系ループ(前述参照)を考えて、新線条体を構成する尾状核と被殻の内側部にいく程、この結果を示す傾向がより強く観察されるかどうか知りたいところである。また、手足や体幹など全身的運動を調べているのではなく、外眼筋の活動で代表して、認識や動機づけによって行動がコントロール(制御)されるか否かを調べているのは、実験データを得る際のニューロン背景(バックグランド)活動による乱れを除去して観察する点ですばらしい手法と言える。すでに、中脳ドーパミン系の大多数のニューロンは報酬期待性の視覚性のみならず、聴覚性の刺激に対してもphasicに反応(活動)するというデータが得られており、情動報酬の過程と接近行動の学習に関わっていることが知られている(Schultz, 1998, JNP. 80:1-27、のすぐれた総説をみよ)。従って、同様に線条体内に聴覚性記憶ニューロンも存在し、情動系によって(リズムをもった運動としての)行動(演奏を含む)が強く影響されると推量される。このように考えると、聴覚系である音の表現にも、視覚系である形や色の表現に類似した脳内機構、すなわち、言語の発音(語ること)のそれとは異なるニューロンのコントロールセンターが存在するように思われる。
この上に、大脳皮質とくに新皮質が発達した動物-哺乳類-の段階になると、その動物の脳髄には、リズム・テンポの表現に加えて、動と静、速と遅、長と短、さらに美と醜、純と不純、協和と不協和など質的に発展した表現方法が発現されるような機構が形成されてくると考えられる。
最も上位にある大脳新皮質はこのprocessの中で下位の線条体の運動を中心とした並列系(上述)ループの中でこの系をコントロールする役目を演じる。聴覚領、視覚領のもつ知覚、認知、認識記憶のシステムは上にみたように、言語系(第二次信号系の確立)と結びついて、後連合野の内でstepwiseに高い段階に昇り、それがdorsal and ventral pathwaysを通り、前頭葉、とくに前頭前野にconnotationのconvert/組換えが起こり、運動系をゆり動かす。このレベルのリズム、テンポは美的な芸にも達し得るものでtempo rubatoの世界も範疇に収め得るものとなる(前述)。彦坂ら(1999)のまとめた図を下に示す(改変)。
最後に蛇足であるが、Area4(第一次運動領)が活動する前に、準備、企画、調和するべく活動する運動関連野は、これらの高次運動野および大脳基底核を中心とする皮質・線条体路、黒質・線条体路、腹側被蓋野・側坐核路、扁桃体遠心路(視床下部、側坐核、中隔核などへの)と密接に関連しているので、芸術や思索に結びつく高次神経活動、すなわち、精神活動を研究する際にこの点を含めて総括的に(その発達過程をしらべることを含めて)考案することは重要な視点であり研究課題である。
一方、鳥海、丸山らの最近のコンピュータ解析による研究によれば、プロの演奏においては、同じ8分音符で示された個々の記号の長さが、楽曲の中の位置的状況により(音程の変化、リズム、ハーモニーなど)メトロノーム時間のようにすべてが厳格に同じ長さの時間で演奏されるのではなく少しづつ長短の差(ズレ)が生じているという。コンピューター演奏などにみられる機械的に時間の長短を正確に乱さずに音を出すという演奏法とは違った美しい演奏が人によってなされるときは、「完全な時間同一性」ではなく、個性あるゆらぎの変異(variation)がみられるとのことである。なお、ある設定されている音符の音の表出に少し長い時間を費やした時には、後続の同じ音符でもこれを修正するかのように無意識が働いてその音の時間を少し短めに演奏するという結果が出ている。これは、演奏者の脳が可塑性をもち、「ズレ」を修正するように働く意志が発動された結果と解釈される。または、ズレをautomaticにmodifyする機構が脳に備っているのかもしれない。
文学的表現を借りれば、音程の落差、音の強度の変化、リズム/テンポの急激な進行、突如として響きわたる不協和音の飛翔、「ことば」と「ひびき」が結びついた時の音空間におけるレトリック(修辞的)奏法の数的比例関係が成立しているロゴス的宇宙(コスモス)内に同調し、また時に乱調するゆらぎ/ズレ等々は聴者の脳に与える外的刺激として意味をもっている。それのみならず、まさにこのような刺激を与えることを旨とした作曲・演奏法が積極的に求められたのが17,18世紀の、バッハからモーツアルトやベートーヴェン、シューベルトに至る時代の音楽であった。
バッハの「マタイ受難曲」にみられるキリストとバラバを対比的に描出した場面など効果的な不協和音の出現によって、とくにロゴス的なものがそこに入ってきたとき、これらの刺激によって脳の活動は全体として(皮質も皮質下も)時に高く(=緊張し)時に低く(=弛緩)する。そして、アフェクト/パトスへの働きかけとしての響きが音空間内に作り出される。このゆらぎを美しく情意を込めて演奏できるかどうかは、音楽芸術家の「頭脳」に内在している共鳴と修正の資質に拠るであろう。これを可能ならしめるものは、脳髄に備わっているズレ/ゆらぎの修正の機構をゆれ動く情動の内で如何に訓練と練習(Uebung)によって、規則を自然の息吹に同調させるかという点に帰する。単に、楽譜上の記号を音に変換するというだけの技法では、音楽用シンセサイザーの定常的発信音にとどまっている。音楽的(または芸術的)効果を期待した演奏をするには、楽音を唯一絶対的な素材として位置ずけることから成熟/昇華させて、ゆらぎをもった持続性の高い音をひびかせる必要がある。この脳内機構の働きは科学的言葉で言えば、脳の働きに条件反射理論をいかに正しく適用するかということになるかと思う。
次のようにまとめられよう。すなわち、脳幹、辺縁系、後連合野、前頭前野を含む脳全体を働かせて、知情意の統一した美しい精神内界を形成させるためには、無条件反射を基にした外界刺激を素材に本能的/生来性とも云うべき脳幹と原始皮質脳を形成させ、その上に、発達期に応じた適当(切)な条件刺激を反復的に与えて、芸術的資質を備えるに足る脳髄を形成しなければならない。そのために要請される第一次条件刺激は、児童期からの聴受(音楽傾聴)を主とする教育と訓練であり、第二次条件刺激は、その上に成立する思春期までに大きく開花する内言語を活動させた思考(索)と響きを融合させることを目的とした訓練と習熟によって到達される高次の演奏技巧の習得であろう。
10]高次神経活動(=ヒトの精神活動)の要諦と精神医学
以下に自由な発想を試みる。書きながら価値あるものを抽出して考えてみたいというのが本音である。素材はいくつかある。それらを一旦ここで整理してみることも無駄ではないだろう。
先ず、視覚系、聴覚系、体性感覚系の興奮が、脊髄・脳幹を明確に分離された形で間脳まで上行して、それが各々、大脳皮質内の第一視覚野(V-1), 第一聴覚野(A-1)、第一体性感覚(S-1)の皮質第4層(内顆粒層)ニューロンに伝達される。この皮質域は柱状に配列された同種の性質のニューロンが集合して柱状構造(columnar organization)をもっている。ここで各感覚様態の細かい分析がなされ、順次段階的に複雑な要素も解析/統合されて、以前に経験された記憶の蓄え(痕跡)と照合される。これが知覚、認知、認識といわれる脳の機能で、後連合野内で行われる。サルの段階になると可成り後連合野は発達してくる。visual, auditory somatosensoryの各領域に異なる機能をもった神経細胞群が同定され、また構造(皮質構築)的にも区別されるので、細かい領域―亜域に区分されることが分かる。それらを連絡する神経回路網の研究も機能解析と並んで成果を挙げている。これらの3種の皮質連合野が重複/隣接する領域に“言語原型皮質”が発達し(サル)、cross-modalの機能をヒトの原始社会で労働とコミュニケーション(情報伝達)のなかで不可避的に発揮させつつ、感覚性言語野が発達してくる。
最も原始的な皮膚や粘膜の感覚から、身体の空間位置関係までを知覚/認知するという体性感覚系の研究も進展しており、頭頂連合野がその場である。この“低次”の感覚は原生動物や魚などでは全身的にダイナミックに皮膚・粘膜(時に嗅い、味も入れる)で反応する。ヒトにおいても祭りの太鼓や笛の響きに皮膚組織がそして内臓上皮の粘膜までも共鳴して踊り出してしまう。
空間認知を視覚性および聴覚性の背側経路(dorsal route)との関係でいえば、視(覚)空間(Gesichtraum)、聴(覚)空間(Gehörsraum)の形成と言うことになろう。視覚性および聴覚性の認知/認識がこれと結びついて高まって行く時には、低次元の知覚・認識から具象的、実体的さらには統一的、抽象的認識に質的に発展を遂げた状態になっている筈である。これは概念を形成することが出来るまでに発展(達)した脳をもった動物すなわち、ヒトにおいてのみ可能である。
この段階の脳とは、次の条件をみたす大脳新皮質が構築(構造的に機能的に、神経回路形成を含めて)されているということである。すなわち、後連合野内に感知された、いわば信号に内在する実体(本質)を抽出して、これをより高次の信号として認識することの出来る後言語野(感覚性言語野)が形成され、上述のように各種の感覚様態の高次機能と密接に結びついた神経回路網が完成されているということである。このステップまではいわば脳の受動的(passive)な機能側面である。動物は外果から得た感覚刺激をその動物の発達段階に応じた形で認知したのち、上記の背側経路(dorsal route)と腹側経路(ventral route)を介して、前頭葉に刺激を伝達し、その能動的(active)機能を発揮すべく賦活する。この際、背側経路は帯状回(中古皮質、大脳辺縁系の一部、意欲に関係する)と、腹側経路は扁桃体、海馬傍回(大脳辺縁系の一部、情動、記憶に関係する)と、相互結合をもって(機能的に言えば相互に情報を交換するように)構成されているので、単なる情報の機械的写像変換ではない、「知」と「情」と一部「意」の第一段階的(原始的)な統合がなされていると考えられる。この後連合野から前連合野に情報を伝える2つのルートをより詳細に考察すると背側経路では、前頭葉の背側運動前野に情報を担った興奮が伝達され、そこで内側面皮質の帯状回前部にある意欲形成関連ニューロンと相互作用する。なお、この帯状回前方域のすぐ背側は6野(補足運動野)にあたる所でPenfieldグループが上言語野と命名した領野を含んでいる。他方、腹側経路は能動運動系に写像変換的に興奮を伝える背側経路とは異なり、どちらかというと、pathosとlogosの融合が顕著で高次機能の発現の素材を粗な形で、より高次の神経活動の分析を可能にする形で包含しているように思われる。
実際このventral routeはサルにおいてさらにヒトにおいて一段と発達した鉤状束(fasciculus uncinatus)を形成しており、その発達はdorsal routeに比較して格段に高い。サルの所見をみて、この皮質間の投射が、運動関連野に直接終わらないで、前頭連合野内の主溝よりも腹側部位(腹外側前頭領域ventro-lateral region)に集中して終わっていることは注目に価する。なぜなら、この領域は進化の面からみてareas46,45など前言語野(運動性言語野)に発展する領域で、後連合野から入った情緒を加味した高度に分析された(言語処理をうけて)情報がここに一次貯えられて、もう一度“能動的”言語分析という情報処理をうけて、しかも、さらに扁桃体の新しい領域(basolateral, lateral nuclei)との相互神経連絡を介して、外界(環境)からの刺激を認識し、価値評価をした高次の情報を文脈の内で捉えて(connotational meaning)、それを変換(convert)し、組みかえた形にして、概念化した抽象化の可能な思考として、前頭前野、補足運動野、帯状運動野など「高次運動野」といわれる領野にその素材を送りこんでいる。高次運動野で運動開始前の準備、指令、連鎖/順序の決定などが企画されて、最終的に一次運動領(4野)に興奮が送られ、そこの運動ニューロンが興奮し、その結果運動がスタートする。
この過程で外界の情報をその有機体が最も多く受けており、“うまく、よく生きていく”機能をもつ皮質連合野からのみ、“たくましく生きていこう(時実)”という機能をもつ辺縁系構造物(皮質と扁桃体などの諸核)に多くの線維が送られているという形態的事実は重要な点である。しかもネコ、サル、ヒトと進化する程、この(恐らく)制御性の投射は強力になっている。前頭葉機能を一括して言えば、能動性の機能で、一次運動野の傷害は運動麻痺として、高次運動野の傷害は運動調節(固縮や振戦も含め)の障害として、前言語野の傷害は運動性失語症として、前頭前野の傷害は精神分裂病者などにみられる自閉症などの能動性障害として発現する。
次の問題として、前連合野から後連合野(後頭、頭頂、側頭の各連合野を合わせてこう呼ぶ)へのいわゆるfeed backループについて一言しておく。要約的に言うと、この投射もdorsal and ventral paths(両方向性成分を含む経路)があるが、殆んどの線維がventral routeを通りdorsal routeの成分は量的に少ない。印象として反対方向成分の半分以下である(サル)。宮下らのサルを用いた実験結果によれば、この“フィードバック”ルートの機能は図形の実物をみることなく、心象(頭の内にある表象イメージ)を基に連想する際に側頭連合野の前方域ニューロンに生起する想起プロセスに関与するとされている。先に考察したように視聴覚の類同性からみて、これは聴覚性の音(符)の順序付けを連想作用としてイメージ的に想起する過程-連想→作曲→演奏→記憶とつながる-においても関与する可能性が充分ある。音楽の演奏者はこの回路を働かせて練習を行っている。
ここで付言しておくことがある。rBF, fMRIを用いたヒトの言語活動の最近の研究によれば、ことばを聞き、語る場合の脳の働きをみると、古典的に考えられていたように(前述)前、後、上の言語野など領野的に3つ部位に限られたものではなく、さらに広範囲な領域(脳全体で少なくとも5-6ヶ所)が動的に活動しているというデータが得られている。この事実は、内言語を働かせて、思考している時にはいわゆる古典的言語野以外の部位も活動しており、脳が全体として活動していることを如実に示している。
これまでの考察の上に立って、ここで音楽を演奏する際に脳がどのように活動しているか、また、響きを捉えて生き生きと演奏するには、どのように心得て、脳の活動の原理にマッチさせたらよいかという点について考えてみよう。
形象を対象とする彫刻家や画家はそれを具象化する前に内的形象のイメージを視空間内に位置づけており、楽曲にたずさわる作曲家や演奏家は内的響きを、音空間内に創造している。専門家は光源や音源が実際に存在しなくとも、形や音のイメージが連合野内に連続的に想起されるように訓練を重ね、そのtechneを身につけている。前に示例したようにfMRIで調べてみると、演奏家の脳では楽譜をみたときに聴覚連合野の(Wernicke)言語野が活動して居り、音の流れを文字の流れとして捉えていることが判っている。この際、視覚性言語野は活動していない点から考えて、楽譜を単に視覚的にイメージングするのではなく、記憶に書き込まれた符号化された連続的な内音のひびきを、小脳・大脳関連ループと大脳・基底核・視床関連ループのなかで学習化された意識下(随意性でないという意味)でスイッチオンして呼び覚ますことによって、運動系を発動させて演奏を可能にさせると考えられる。視覚性芸術家の場合も、たとえば、石を削る前にミケランジェロがすでにその対象・石塊を前にして心の内に削り出されるべき形を予見するとき、恐らく視覚性言語野が活動しているものと推察される。そこには、聴覚、視覚のみならず、一般感覚、内臓感覚をも含む全感覚(の認知)が総動員されて、そこに感動が生まれてくる。そしてこの感動の投射によって、視覚性言語野に創り出されてくるimage(イマージュ)を具象化させるべく、実際の形を石塊から削り出す作業がなされる。その際に、視覚性言語野に形成されている内的イマージュと、削り出されつつある実際の彫像形とが、絶えず照合されてゆくことは改めるまでもないが、このような内的イマージュの外化の過程が芸術における創作であろう。このときの形を響きに置換すれば、それは音楽の創造になる。なお、この内的イマージュと外形との照合が必ずしも内→外の一方向的なものでなく、内↔外の相互的照合・修正であることは、たとえばベートヴェンが残した多数の創作スケッチから知ることができる。内的イマージュが外化されることによってより正確に、もしくはより良いものとして認知し直され、さらにまた、この修正された内形によって外形作り直されるという作業が繰り返されるのである。つまりそこには、内的イマージュとして形象化されている一種の概念の存在が認められる。あるいはそれは言語野としての機能に根ざした言語性の概念であり、それが視覚性もしくは聴覚性言語野に投射されて、ベートヴェンの場合には内的響きのイマージュが抽出されてくることが推定される。いずれにせよ創作においては、創作活動を通じて伝達されるべき事柄に根ざして内的イマージュは形成されてくるので、言語野における概念を作における原イマージュとして捉えることは可能であろう。
この時には、前頭前野(=前頭連合野)→高次運動野→一次運動野(→そして一部の興奮が後連合野にフィードバックされる)として形成される神経回路neural circuitが言語野および情動の回路と結びついて形成される。歌詞の内容を識って聴く、オペラやミサ曲などジャズ、ヴォーカルまで含めた種々の声楽曲において、識らずに聴く場合に較べて情動中枢が強く刺激され、芸術の享受において、それもlogos とpathosに働きかけて芸術作品によって伝えられるべき事柄の認知へと享受者の脳を活性化させるに当たって、言語中枢の関与が重要な役割を演じていることをあらためて意識させる。一つ一つの言葉に含蓄される意味の理解、いわば言葉の内的意味・機能に対する理解の深浅に芸術創作ないし鑑賞が大きく依存することはそれを体験する者が切実に識る処である。芸術の働きかけによってもたらされる美意識、情動性、思想性の涵養、さらにはそれらの総合的働きによって作り上げられる人間性の完成を如何に位置づけるかは、脳全体の活動とそれを支える上記の脳内関連領域を結び付ける神経回路(網)の形成という問題に帰着する。
この際、能動性をもった運動表現の時に考えておくべきことは、最終の出力が単に一次運動野だけにとどまる(初心者は一つ一つ意識的に運動領内のニューロンを活動させることを学ぶが)ものではないことである。これに関与するものは、脳幹内にその機構の中枢が存在する歩行や自律系のリズムに始まり、小脳と大脳との間の相関作用が作られ、中脳ドーパミン系と結びついた大脳皮質―大脳基底核(広義の)の広範囲にわたる働きが考えられる。これにあらゆるレベルで神経伝達物質、ホルモンなどの液性伝達の機構が関与している。
最後に、以上の神経回路は、どのように、何歳くらいまでかかって成熟し、完成するのであろうか。これはある意味では仮説を重ねなければ説明できないものである。この内容を全般的に取り扱うのは無理難題に近いので、ピアノ奏法、情動/概念の形成、自我の発達、そして脳の発達、神経回路形成の異常という順序でそのいくつかの要点に触れながら論を進めることにする。
ピアノを弾くためには指だけでなく、手首、腕、さらに身体全体を使うような色々な運動が必要である。しかし、一番の基本になるのは常に指先を意識して、キー(鍵)をしっかり「つかまえている」という感覚を常に持っていることが大事である。指の運動性について言えば、10本の指がそれぞれ独立してよく動かなければならない。指の大脳皮質運動野における再現域(representative area)はサル、ヒトで拡大する。この手の指を動かすための皮質運動ニューロンが集合している領野(手指の再現域)は舌の運動に関する再現域と運動性言語野に隣接している。これらの領野は短い連合線維(弓状線維)によって結合されている。美しい音を響かせるよい演奏をするには、おそらく言語の発音=舌の運動+発音をコントロールする運動性言語野との相関の中で、指の脱力によってもたらされる指の自由自在な動きをコントロールして、指で語らせる打鍵を促す、言語表現機能を司る運動野からのニューロンの刺激による打鍵をせねばならない。子供にピアノを教えている音楽家に聞くと、上述の大脳皮質構造(髄鞘形成も含めて)の発達や指骨の発育の他にも、指の運動に関連する靱帯、腱、筋、関節および支配神経の習熟的完成が可能になる丁度思春期(15-20歳)の頃にこの技法が身につくそうである。思春期には声帯も形態変化し、ホルモンのバランスも変化し、20歳を過ぎて固定化する。
前頭葉の最後部に、中心溝に接して、ヒトでは第一運動野がある。そして下肢が内側面に、胴体、上肢、顔面、舌、指という順に大脳半球外側面で背側から腹側にかけて、舌、指の複現域などは特に広い面積をしめる形で配列されている。この部位の神経細胞の興奮が即外界に対する能動的活動すなわち運動の開始である。
言語(ロゴス)と情動(パトス)の作用ないし処理により感覚の認識に感情、情緒が込められ、彫のある陰影が加味されて、前頭葉内の前頭前野に入力されて、そこでその情報が、最も適切に処理され、変換され、意味づけられて(convert, connotational processing)例えば芸術では美的表現形態として外界に投射される。
これらを可能ならしめる脳の成熟のうち、上述したように多元的ではあるが、それらの全体を総合するものとしての“神経回路(網)”が存在するという形態的確証はまだ得られていないが、その可能性は充分にある。脳の成熟と人間の成熟とが不即不離であることは言を俟たないが、そのような成熟に関してこの神経回路網の形成とその成熟(完成という言葉は使わない)によって、知、情、意の働きが、人間性を獲得し、思想性を帯びた概念を形成することが出来るようになるのは、この思春期(puberty)においてである。この形成過程の中で青年は他人との共同作業におけるコミュニケーションを通じて、合成語を作り、動詞と名詞と助詞を結び付け、動詞の時制を知り、能動と受動を区別し、動詞や形容詞や分詞さえも中性名詞化し、抽象化した名詞や動詞を使用して、言葉を柔軟にあやつれるようになる。同時に人間社会の中で怒り、愛し、自己主張する中で、喜怒哀楽をコントロールすることを学ぶ。また、仲間との会話、討論、読書、執筆などを行う中で哲学的に思想を表示できるようになる。すなわち、思考の全機構(メカニズム)を想像の所産から切り離して観照することが出来るようになる。勿論このプロセスはnature(遺伝子)とnurture(教育)の合体産物として成熟するものである。内省的に思索し、哲学することが好きになり、自我が確立される時期である。
ところで精神分裂病の多くは思春期以降に発症する。他人とのコミュニケーションが上手く行かず、自閉的になる。能動的志向が徐々に低下し、知的傷害はないが、思考(思路がたどれなく)の傷害や認知の傷害があり、連合弛緩や幻覚、妄想が時に現われる。古人が一言で「精神内界の乖離」と言い当てているが、分裂病はそのままに語義的に表現して、split mind(分離された心/精神)である。神経学の対象となるsplit brainとは異なる。前者は精神のsplitが時々周囲の人達にも分かるように顕在化するのに対して、後者の方は常に存在する明らかな神経学的症状である。
“Split-brain(分割された脳)”が脳の器質的障害で永続的であるのに対して、”split-mind(分離された精神)”は機能的なdisconnection syndrome結合解離性で非永続的な徴候を示す。この分野は精神/こころの領域に属し、多次元的である。多少遊戯的であるが、一方のsplit-brainの症候は19世紀以来の研究である失語(言語障害)、失行(行為障害)、失認(認知障害)に代表される大脳病理学・神経学の範疇で、最近は1960年代にはじまるSperry, Gazzanigaらの大脳交連線維(脳梁、前交連)の切断による症状の研究が有名である。もう一方のsplit-mindはSchizophrenia (shizos=separate; phrenia=spirit, mind)を単に源語的に置き換えただけのもので、元来、脳科学と結び付けて用いられてきた用語ではない。この精神内界の乖離(intrapsychische Spaltung)という精神医学の用語の概念を、ここで、脳機能学の立場から考察してみようと思う。この論理が成り立つとしても、多分に形式的ではあるが。何が、どのようにsplitされるかを考えてみると、
①前脳(終脳と間脳)と中脳・後脳(中脳以下延髄まで)との間のrostro-caudal間(皮質・皮質下間)、
②右大脳半球(略して右脳)と左大脳半球(左脳)との間のleft-right間(左右半球間)、
③大脳皮質前部域(前頭葉)と後部域(頭頂葉・側頭葉・後頭葉)との間のantero-posterior間(前頭葉・非前頭葉間)、
にみられる一時的なsplit(分離)と便宜的に大別されよう。
以下に①、②、③の順に考察する。
①の場合:
外界からの感覚刺激が感覚器官内の感覚細胞を興奮させるが、嗅覚以外の情報処理ができず、外界に働きかける運動形態も反射的である。先のマック・リーン(MacLean)の脳の階層性の三位一体の図式から言えば爬虫類脳のレベルである。奔放な欲動的なものを本質とする情動発現に対して上位からの抑制がかからない。外界から得る感覚入力は嗅覚がほとんどで他の感覚様態と体内からの内臓感覚は中脳のレベルで伝達が機能的・一時的にストップすれば全く無意識的、無条件反射的となる。また、間脳/基底核のレベルでストップすれば、運動処理は線条体で、感覚処理は視床で、情動処理は扁桃体を中心とする辺縁系領域で、自律系・内分泌系の機能は視床下部、下垂体系で、大脳皮質からのコントロールなしで行われることになる。生物体の精神/こころ(と呼ぶとして)の状態像としては、混沌の中の無秩序的統一ということになろう。すなわち、生体機能としては、生活環境の変化に適正に素早く適応することが非常に困難な状態にあると言える。
②の場合:
魚類の左右大脳半球には、嗅脳と呼ばれる終脳の前方部に膨らみがみられ、それが終脳のほとんどど全体を占めている。ヒエラルキーのより高い感覚様態に属する視覚、聴覚系の分析器としての大脳皮質は魚類には存在しない。これらの感覚は中脳レベルで処理されるからである。左右半球間の情報処理は動物の高等化にともって中脳・後脳レベルの段階から終脳に移行する。詳しく言うと、発生学的に古い系統に属する第Ⅷ神経系では延髄で聴神経系も前庭神経系も左右の間に形態学的にも機能的にもコミュニケーションが認められる。体性感覚系は、脊髄レベルで原始的触覚、温痛覚が、延髄レベルで識別的触覚が左右相交差して、それぞれ間脳の視床特殊核まで脳幹網様体内を上行して興奮を伝えている。その伝達経路の途中で多数の側枝を網様体内の神経細胞に神経突起を出してシナプスをつくっている。この側枝と網様体ニューロンの突起を介して左右側間のコミュニケーションが行われている。この大脳皮質レベルに達しない、意識下の(意識にのぼらない)脳幹内全体(延髄、橋、中脳)の左右間を含めた情報処理機構の興奮と抑制の過程は覚醒と睡眠と呼吸循環などのリズムの根底(basis)をなすものである。
興味あることに、感覚情報が集中する、そして哺乳類などの高等動物では、脳の下位中枢から上位中枢すなわち大脳皮質への上行性刺激伝達の内で中継核として位置づけられる視床核においては、左右の交連は機能していない。実際、解剖学的に左右の視床を連絡するかと想像される視床間橋(Adhesio interthalamica)(中間質、Massa intermedia)と呼ばれる構造物がある。内部に神経細胞を多数含んでいるが、交連線維は少数で、しかもヒトでは退化的(20%のヒトに欠如する)である。とっとも、より下等な動物(下等哺乳類以下の)では、運動機能の処理が大脳基底核に属する線条体を含む“錐体外路系”と相呼応して、ここの視床諸核で感覚情報の最終処理が行われる可能性が充分に考えられるのであるが。
この他の交連線維としては、左右の視蓋(中脳背側部)、視蓋前部、Darkschewitsch核、カハール間質核などの諸神経核間を結ぶ後交連Commissura posteriorや、情動・記憶系に属する辺縁系に関連する手網交連(視床上部にある)、それに脳弓交連や海馬交連(原始皮質Archicortexを結ぶ)、前交連(左右の嗅脳部―古皮質Paleocortex―を結合する系統発生学的に古い終脳交連であるが、高等動物ではさらに側頭葉新皮質の一部の交連線維もこれに加わる)がある。
しかしなんと言っても、ヒトで最大に発達し、左右の大脳新皮質neocortexを結合する系統発生学的に最も新しい交連線維は脳梁Corpus callosumである。これまで脳の構成として左右対称性symmetriaと左右各半部の主に相同部分の結合について専門外の研究者にも判るように紙面を費やして来たが、ここでsplit-brainの本論である脳梁切除脳についての研究とこれらから類推される、この範疇のspilit-mindについて以下に考察を進めることにする。
臨床的な観察によると、海馬は一側を切除しても、他側が正常に働いている限り、日常の生活に支障をきたすことはない。しかし、両側を切除すると過去の意識の流れを再現する機構が失われてしまうという。大脳新皮質の半側切除についての手術後の経過報告については平尾の研究があるが、一側半球だけでも-例えば右半球のみ残された患者においても-認知機能とくに条件反射第二信号系(言語系)に関与する機能低下はみられるものの、通常人格を認められるだけの能力は保持してることが知られている。(更に検討すべし)。
以上のような切除や本論で問題とする脳梁の切断の手術は、20世紀の前半に、腫瘍患者やてんかん患者の症状の進行を防止する治療目的で行なわれた。Sperry, Gazzanigaらは、脳梁(一部前交連も加えて)を切断した症例、いわゆるsplit-brain患者を対照として種々の検索を行った。そして、それらの患者において、左視野の物品呼称障害、左視野失読、左手の失書、右手の構成失行などが観察された。重要な点は、この大脳交連線維切断例の研究によってヒトの左右半球間に機能分化が存在すること、それは如何なる性質のものか、左右半球間の話し合いすなわち、情報処理交換の作用、ひいてはいわゆる精神活動としてどのように統合されているのか等々について、具体的事実(データ)に基づいて、またそれらを参照して、少なくとも大きくは誤らない範囲の考察をすることが出来るようになったことである。
考えてみると、中脳以下のレベルでは、恐らく間脳以下のレベルにおいても明らかな形においては、解剖・生理学的に左右対称symmetryで差違はない。脳が進化・発展して、ヒトの大脳半球に発達した時にはじめて、左右半球に機能の差違が生じてきたように思われる。その証拠にサルにおいては認知機能の左右差と言語機能の問題の研究を行って結果が出せるという実験計画が組めないということである。
ヒトとサルの大脳皮質の発達で著しく異なる点は、皮質内の細胞(ニューロン)の形態や配置などの皮質構築を度外視すれば(本来は最も大切であるのかもしれないが)、①後連合野および前連合野内に各々に発達した言語野の有無、②前頭葉とくに前頭連合野(=前頭前野)の著しい発達、および③連合線維(=同側性皮質・皮質間結合線維)の発達とくに前頭葉と頭頂・側頭葉との間の線維結合の発達の差というようにまとめられよう。
このような条件が整った所に両半球間に優位性が見られるようになる。しかし、この優位性はあくまでも相対的なものである。この点で言えば、左半球は、物品呼称、話し言葉の理解、音読、読解、書字など、明らかに言語機能の処理過程において右半球より優れている。
他方、右半球は、視覚的に図形や模様を認知し、断片化したものを合成・形成していく機能や、直接知覚的統合的あるいはゲシュタルト的な過程において(Sperry)、また、手を使用して空間的情報を模索し変容させる機能において(Gazzaniga)左半球よりも優れていると考えられている。
ここで言語における記号(文字、音声)の処理過程について心理学的考察をしてみたいと思う。繰り返すが、左右間に差(優位性)は存在するが、絶対的でなく相対的である。そして互いに情報を交連線維系を介して交換し合っている。以上を念頭においての考察である。右半球の働きは、記号によって伝達される意味内容の具体的像と経験的・情動的にむすびついており、その処理様式はイメージの助けを必要とする推理課題を解くのに適している。それに対して、左半球の働きは、意図的な言語学習に適しており、意味をになう記号と記号との間のいわば法則的な関係、あるいは、その対象が備えている属性の間の関連を結び付けるように機能しているいるように思われる。こうしてみると、脳梁線維の働きによって、右半球が獲得した知識は、やがてその具体的で直接経験的な色彩を失い、一般化されて左半球に移っていく。この特殊化から一般化へ、また、未分化から総合的分化への道程は将に言語機能の発達の過程と結びついている。「あゝ、あの時、あのようなことがあった」というようなイメージ的な非言語的知識に基づくエピソード記憶はイメージ能力が低下するその年齢的推移と共に、「この意味は斯々如々」という言語的知識に基づいた意味記憶に比重を移していく。
最後に、ではあるがわれわれの研究課題に関連して最も重要な問題である、言語(ことば)とくに音声言語と音(楽)(言語性と非言語性)・聴覚の問題について角田忠信氏の研究について、またその理論についてとりあげてみたい。耳鼻咽喉科学の臨床家であり、すぐれた研究者である角田は発達期の脳の可塑性、つまり脳は形態学的・機能学的・化学的に環境の変化により大きな影響を受けるということに注目して、人類に特徴づけられる言語修(or獲)得と聴覚系(言語と非言語情報に分ける)の関係およびそれらの大脳半球優位性について調べた。そしてその結果を『日本人の脳』(1978年)として発表した。日本人の聴覚系の脳機構としての興味ある結果を以下に紹介しておく。角田によれば、『日本人について、二つ以上のバンドノイズの組合せ音について、その周波数比を変えて優位性を求めると、整数比にある音は左耳(右半球)が優位になるが、非整数比の関係にある音は右耳(左半球)が優位となることが見出された。このように、日本人の聴覚系は言語と非言語情報に分ける前段階で、外界の音を自動的に音形に従ってharmonicとinharmonicの音に振り分ける機構をもつと考えられる。このように左右の聴覚系の優位性はinharmonicとharmonicとに整然と分類されるが、あまりにも複雑多様な働きを示す脳の別の明解な面を表わしていて面白い。』
●表1 日本人と西洋人の言語音、自然音、楽器音の大脳優位性パターンの比較
表2 電鍵打叩法で得られた日本人の聴覚の優位性
また、角田による表1と表2を引用してここに掲載し、後日の考察の資料としたい(他の結果も含めて、聴覚系/2音の整数比、非整数比の選別について(角田1426号)、など(勝木先生論文も入手したいし1464-6号)再検討。また植木幸明(1468-70号)。(以上メモ)。
いろいろ、ここで、中心課題である精神病とくに精神分裂病のsplit-mind(大胆に言えば分裂病脳の一時的・機能的切断)の問題について考察する準備が揃ってきたように思われる。しかし多分に仮説的で顰蹙・噴飯ものであるかもしれない。
①の皮質・皮質下がsplit(切断)されたときは、皮質からの抑制のきかない情動系の障害で能動的には統一がきかない、定型的、反射的、衝動的な面が現れてくるであろう。②の両大脳半球間のsplitとしては、言語系と非言語系の情報の交換処理が上手くいかないので言葉や思考の連合弛緩や奔逸がみられ、認知の概念化が傷害されるであろうから幻覚、妄想(気分)、自我意識の障害、自我や自己空間位置の確立の障害として、させられ体験や両価性といった症状が、③の前頭葉と頭頂・側頭葉間のsplitである能動性障害、自閉性、意欲減退、感情鈍麻などの徴候と結びついて出現することが考えられる。臺弘がくりかえし指摘するように、分裂病のmindはいつもsplitしているわけではなく、時々分裂splitしてその症状が表面化するだけである。決して脳組織に穴があいたとか、破壊されているとかの類の恒常的な非可逆的な、まして遺伝的なもののみによって規定されているような“恐ろしい”“狂った”病気ではない。治療により、良くなる疾患である。治癒不能のもの也、と世人の素人を相手に信じこませるような俗説に惑わされてはならない。
③で問題にされている、前頭葉と頭頂・側頭葉との間の、いわゆる前後の大脳皮質の機能的splitの関係は、近年Ingvarら(1974)によって開発された脳局所血流(rCBF)の研究によって、慢性分裂病で調べられた。それによると、脳血流量が前頭連合野では正常脳で調べた平均値より少なく、頭頂・側頭連合野では逆に多いことがあきらかにされている。
付言するが、最近に至るまで、種々のデータを示して片側半球だけで-とくに右半球について-意識を持っているか否かというような議論があるが、元来人間の意識というものは、空間認知、自我の存在の認知、言語機能の働きなどを包含した社会的、communicative/geseligで全体的なものであるから、そのような視点に立脚して物事を考え語ることが求められるのではないだろうか?
ラットやサルにアンフェタミン(ヒロポン)を注射して実験的に分裂病モデル動物を作成することが行われてきた(臺 弘)。投与を受けた動物たちは“分裂病類似の症状”を示し、群から離れて、おどおどしく行動する。アンフェタミンは過剰ドーパミンの作用と同様に(アゴニスト)、ドーパミンの受容体(レセプター)を刺激し、過敏な状態にする。分裂病患者の陽性症状を軽減するクロルプロマジン系の薬物は、ドーパミン受容体の遮断薬(ブロッカー、アンタゴニスト)として作用し、これらの実験モデル動物の症状も緩和する。
最近西川らによって、この分裂病実験モデルを作成する際に、薬物を投与をうける動物の年齢(時期)と薬物への依存性が検討された。その結果将にヒトの思春期に相当するネズミの幼若期に薬物を投与された時、動物は分裂病様症状が誘発されやすくなることが示された。
専門的記載ではあるが、重要なことなので、以下に西川らの研究に関連した事項について考察することにする。
(文献)
西川徹ら 精神科治療学 12(1997)617-623
車地暁生 臨床精神医学29/4(2000)381-389
精神分裂病の複雑多岐にわたる病態を単純に2分してこれを幻覚、妄想などを主体とする陽性症状と、自閉、感情空虚、意欲の減退などの陰性症状と便宜的に名付けられている。陽性症状には脳内ドーパミン神経伝達の亢進(D2,D4受容体の過敏性反応hypersensitivity)が、陰性症状にはグルタミン酸神経伝達の低下(NMDA受容体の遮断)が関与すると考えられている。なお、分裂病の病因として、脳の器質的変化すなわち、形態学的ならびに組織病理学的変化が最近数多く報告されており、その内で、神経発達障害仮説が提唱され、胎生期に始まり、生後から思春期、青年期に至る神経発達過程における障害(異常遺伝子の発現、様々な要因によるストレスの液性伝達機構を介した影響などを含む)が考えられている。実験分裂病モデル動物を作成する方法として、分裂病様症状発現薬のうち、アンフェタミン類(メトアンフェタミンなど)やコカインは抗精神病薬反応性の陽性症状を発現させる。他方、フェンサイクリジン(PCP)をはじめとするNMDA型グルタミン酸受容体(NMDA受容体)遮断薬は抗精神病薬に抵抗性の陰性症状を引き起こす。また、実験動物ではこのような薬物を投与することによって生じる行動ならびに分子生物学的変化が、その発達段階によって大きく変化することが判っている。例えば、幼若期(生後1~21日)のラットでは成熟期(生後25日以降)とは異なりメトアンフェタミンによる逆耐性現象が生じない。
このような事実をふまえて、西川らはメトアンフェタミンに対して発達依存的で抗精神薬によって抑制される応答性をもつ遺伝子を検索した。分裂病の多くが脳が成熟する思春期以降に発症するという臨床的事実と上記の動物実験の結果を勘案して、分裂病ではある種の情報システム系に何らかの障害が起きていると考えた。この情報システム系を支えるものは、ある一定の神経回路(網)の活動に他ならない。このシステムないし神経回路は幼児期から思春期にかけて脳が発達する時期に一致して発達し、成熟を遂げるものに関連すると考えられる。西川らは、ラットにメトアンフェタミンを急性投与した際に生じる最初期遺伝子の一つであるc-fosの脳内発現パターンが、幼若期と成熟期で異なることを見いだし、とくにその発現の仕方が大きく異なる大脳皮質と線条体に着目し、逆耐性現象に深く関与する遺伝子群を検出して調べている。このように薬物応答に差のある遺伝子をクローニングし、選択し、それらの候補遺伝子の全構造を決定し、抗体を作製し、それらの脳内分布を発達の段階を追って調べることが出来る。
今後共、①精神活動と高次神経活動との関係、②伝達物質や受容体と遺伝子レベルの解析、神経組織や神経回路の形成など多次元レベルの統一、③脳の正常発達とその障害が現わす精神異常、等々、を広い概念として捉えていくべきである。とくに精神的に向上するプロセスの脳内機構を支える神経回路(網) -このなかに記憶、情動、意欲に関わるPapez回路、Yakovlev回路や、認識や能動的発現に関わる皮質レベルを結ぶ背側経路や腹側経路なども共に関与する- が同じ思春期に成熟発達をとげるということは非常に示唆に富む事実であり、これを結びつける形で、発展性ある見解に到達できればと思っている。
幼児期,児童期,思春期、成人期と発達、成熟する脳内機構としては、ニューロンの突起、とくに樹状突起の著しい発達、髄鞘形成(グリヤとくに希突起膠細胞の発育)などにみられる遺伝子発現に依拠した内的要因と、条件反射第二次信号系にもとづく言語機構と家庭内および社会における共同作業、知的および情操教育などの環境要因の両者が相俟って、種種のタイプの“神経回路網”が機能的に活性化され強化される。これがヒトの高次神経活動すなわち精神活動を支えるものである。このように発達する“神経回路網”が条件反射機構にもとづいて教育(知育、徳育、体育、情育)される過程で、適切な時期に関連遺伝子が発現し、物質的基盤が整い液性伝達系と神経伝導系が動的に共生cohabitする。芸術、学問における創造活動は、これを基にした脳の能動的活動に他ならない。
上述の高次神経活動が如何なるふうにか生じる何らかの原因により -近い将来解明されることが期待される- 異常をきたし、偏った(狂った)状態になったときに精神の異常をわれわれは観察する。精神異常者、とくに分裂病において、妄想知覚や幻聴などの認知障害、思考障害が如何にどのような形で、起こるのであろうか。手に余るむずかしい問題であるが、考えてみることにする。素人眼にみても分裂病者の場合、脊髄、脳幹レベルの上行性伝導路システムや大脳皮質の第一(次)感覚野-視覚領(V1)、聴覚領(A1)など-の障害ではなく、連合野の障害であるように思われる。しかし、本当にそうであろうか?むずかしい問題である。感覚知覚作用の認知障害としてみた場合、視覚性刺激にしろ、聴覚性刺激にしろ、それをパタン化して、Gestalt的に全体的、総合的に対象(objet)を捉える(認識する)ことの障害であると思われる。外界刺激を知覚するとき、それを認識し、意味づけし、価値判断し、以前の記憶(条件反射により得られたこれまでの痕跡)と照合させる。人間は事象を認識するとき、部分から構成される対象を捉える際に、外界からの刺激をパタン化して、全体的に把握しようとする。この際に、表象化し、概念化し、それらを連関(連携)づけて思考を組み立てるのであるから、ことば、言語中枢を活動させていることになる。
安永浩によれば、正常者のもつ認識の原則パタン:a≧bが、分裂病者ではひっくり返っており、病者が自ら述べる体験はあたかも、その逆転(a<bと描き得る)を示しているかのようだと言う。いわゆる「パタンの逆転」の理論である。これについて考察することは小生の理解を多分に越えているが、一応解説する。「パタン」とは、自-他、質-量、全体-部分、因-果、生-死などにみられるA-B構造のことである。安永が呈示するこの「パタン」なる基本概念は観念的色彩が濃い。彼は以下のように説明する。すなわち、
[ 正常人の知覚は一般にa=bと記号し得る体験である。即ち主体的能動性がまさに外界の存在圧力とつりあっている。しかもaの論理的先行という前提は保たれているから、その知覚像は自分にとってだけのことであって、もしかするとまぼろしであるかもしれぬというしっかりした検討の態度をとることができる。分裂病型幻覚(主に幻聴)では像そのもののあいまいさや非常識性にもかかわらず、また意識や知性の健存にもかかわらず、ふしぎなくらい病識をもてない。これはその外来性(b性)の方が自明な前提と化し、主体性(a性)の方がそれにかろうじて対応する、条件的偶然的なものになっている、という把握をすれば、どうにかこの体験の「実情」をうかがい知ることができる。]
すなわち、分裂病者の知覚像・認識パタンは実体に欠ける幻影的(phantom)なもので、正常者のそれがA-B構造とすれば、B-A構造になるというように認識の原則であるところの「パタン」の形成に逆転が起こっているという「ファントーム仮説」である。
これに関連して臺弘は、彼が提唱する「瞬間意識」の中では正常者においても「パタン」の逆転は起こりうるものであると考えている。臺は「瞬間意識」について次のように述べている(私信)。
[ 私の設問はいつも臨床の現場から出てきます。物差し落しによる単純反応時間テストによると、対照正常者の平均から5~10cm (25~45 msec) 遅い患者の行動や言葉には一瞬の遅れが感じられます。これをWertheimerの仮現運動視:TVや映画のコマ送り時間の1/25, 30秒や、flicker fusionの閾値の50msecや、Efronの実験:red and greenのflashの20msecづつの連続がyellowと知覚されること、Michotteの「因果性の知覚」実験:2点の無関係の運動が因果関係があるように近くされるのは50msec以内の出来事、などと一緒に考えると、時間意識の認識は量子的で、その単位は25~50msecであるらしい、これを「瞬間意識」と呼びたいというのが私の意見です。この意見が精神病理学的に重要と思われるのは、分裂病に見られる妄想知覚や幻聴などの体験が「瞬間意識」内に起こる現象と見なされるからです。これは分裂病者に起こるバウム・テストの際のGestalt崩壊や、精神生理実験で知覚入力の関門瀘過機能の障害(prepulse innhibitionの欠落、backward maskingの過剰)などが、どれも50msec以内の現象であることと関連します。幻覚がla perception sans objetと云われるのも無理ありません。それは知覚と同じレベルの現象で、それだけに現実味をおびて病識を得難く、生活史記憶の中に取り込まれて長く後に影響を及ぼすと考えられるからです。]
まとめに入るが、これまで①情動機構、②認知機構、③総合的能動機構(小生の新語)という脳の高次機能について述べて来た。構造的には①脳幹(延髄、橋、中脳)、②前脳の低位中枢部(間脳、基底核、大脳辺縁系)、③前脳の最も発達した部分である新皮質(そのうち、後連合野と前連合野に特に注目する)に区分する。この区分は、マックリーンの①原始爬虫類脳(protoreptilian brain)、②旧哺乳類脳(paleomammalian brain)、③新哺乳類(neomammalian brain)に相応する。また、時実の①脳幹・脊髄系(生きている、反射活動、調節活動)、②大脳辺縁系(“たくましく”生きてゆく、本能行動、情動行動)、③新皮質系(“うまく”そして“よく”生きてゆく、適応行動と創造行動)にも対応する。また、神経心理学的に新しい考え方としてルリアの三つの基本システム:第一(二、三)ブロックまたは第一(二、三)機能系、すなわち、①視床、視床下部、網様体、辺縁系皮質なるシステムで上向性並びに下向性網様体賦活系の働きが下層にあり、欲求・動機・意識を支えている。次に②後部(中心溝より後ろ)大脳皮質(頭頂葉、側頭葉、および後頭葉)の機能としての情報の受容・分析・貯蔵と関係したシステムがあり、知識体系の基礎を作っている。③大脳の前頭部、とりわけ前頭葉皮質の領域で、人間でとくに著しく発達している部分で、人間の活動を意図し(意欲)、プログラムし(計画)、実行する(活動)、さらにその上に、その行われた活動の成果を最初の意図と比較照合し、結果に価値判断を下す。場合によっては、再度プログラムを組み直す。というような複雑な「計画と創造」の基礎となるプログラムと決定の系が見られる。
以上の複数の研究者たちによって提唱されてきている3層のブロック構造ないし機能系が、神経回路網を通じて、および、液性の伝達系を介して、密に連携して、脳の活動を全体として行っている。高次神経活動すなわち人間の精神活動はこれら全体の結果として表現されるものである。この際、情動系にも、認識系にも、能動的決定の系にも、パブロフの上記3つの条件反射系(1つの無条件反射系と第一および第二条件反射系)に照応した、本能から言語思考にいたるまでのヒエラルキーが存在することに注意しておく必要がある。
さらに、精神神経医学における諸機能の階層的秩序(ヒエラルキ)とその崩壊については、イギリスの神経学者ジャンクソンの「階層理論」すなわち「神経系統の進化・進展(evolution)と解体・退行(dissolution)の思想がある。Spencerの進化論を継承してあてはめたものであるが、ジャクソンは神経系の進化をより単純で次元の低い機能(組織化された、自動的な、低次の中枢)から、より複雑で高次元の機能(組織化されていないで、出生後に組織化され続けることになる、随意的な、高次の中枢)へと移行・発展するものであるとした。そして、その頂点をなすのが「精神の器官」(organ of mind)、または意識の身体的基盤であると考えた。
そしてその進化の過程を逆行するものとして神経系統の病的解体ないし退行を説明した。退行によって起こった上位の水準の機能の欠損症状として「陰性症状」を、また、病的過程によって破壊をまぬがれた下位の水準の機能の解放症状を「陽性症状」とみなすた。ジャクソンは神経疾患と精神病とを統一した原理によって理解しようと試みた。ジャクソンの解体の概念は後世に大きな影響を与え、エー(Henri Ey, 1900-1977)によって新ジャクソニズム(neo-jacksonisme)として発展された。エーはジャクソンの神経機能の進化と解体の理論を精神医学に適用したといわれている。
今後、脳の研究、とくに高次機能の研究が発展し、精神活動の科学的解明に有効なメスが入り、児童、青年の脳の発達に適合した教育(知的なものだけでなく、情操もそれ以上に大切であるという認識をもって)の指針が示され実行されることが切に望まれる。同時に芸術面での教育、芸術活動、精神医学の治療面での進展に大きく寄与することになろう。
11]条件反射と心の問題
自然科学者は物質からなる自然を対象として、物質の性質、合成、発展について、(ありのまままの)観察と(人為的に操作を加えるという)実験の結果を(自然の法則に合っているかを)考察し、それを発表する。その際、自分が得た所見結果を、これまでに得られている過去の知見/業績と比較・検討する。両者が合致している場合には、新しく得た結果が学問的に発展し、深化されていることを強調して公表する。不一致の場合には、その点を再度実験などにより再検討して、確めた内容を、過去の業績との相違点に主たる焦点をあてて、先人の仕事に正当かつ適切な敬意を払いながら主張する。
脳という物質の最高段階に発展した集合体を対象にして、謙虚に且つ、厳格に、自然の法則に従って研究する限りにおいて、研究者(=人)は大きな過ちを犯さず、道を踏みはづすことはない。研究機器、技術が時代と共に進歩し、次々に新しい知見が生み出され、それを基にして研究対象がより明確に再考され、議論される。
新しい概念・志向がこのように形成されて科学は発展する。時代の制約があるので、その点を考慮して先人の業績を評価し、正しい部分を引継ぐことは後人の義務である。それと同時に、不備な点を率直に指摘し、批判することも、たとえそれが未熟な段階の見解であったとしても、重要なことであり、任務であると考える。われわれはこのようにして、偉大な先達を乗り越えて行くべく、研究を推進させようと努力している。
科学者にとって、自分で納得した事実に基づいた事象の上に組み立てられた見解を主張することが何よりも大切で資質を問われる事柄である。中枢神経系(脳と脊髄を合わせてこう呼ばれる)の物質の構成、性状、変化を観察し、考察している限り、解剖・生理・生化学・分子生物・病理の各分野において、自然科学者は、自然に問いかけてその反響(エコー)を聞いている限り、思考の結果を発表してもその思想の内に迷ったり、誤ったりすることは大局において少ない。
ところで、人間の脳の活動の産物の総体である心や精神の問題になると実験によって検証することが容易に出来ない対象であるので、話はそう簡単にはいかない。人間の精神活動は言語機能と深い関連をもっている。この点、とくに注意を払って考察しなければならない。ところで、脊髄や脳幹のレベルにはない認識、思考、判断、言語、情操、等々の高次神経機能の座である大脳皮質(とくにヒトの)の高次機能を問題にするとき、偉大な先達たちの中には、舵を失ってしまう脳研究者もいた。残念なことに、世人は著名な学者の思想を無批判に受け入れてしまう。無理のないことではあるが、学問の発展を妨げること甚だしいと言わざるを得ない。
高次神経機能を脳内の神経細胞の働きとして自然科学的手法で捉えようと努力した脳外科医ペンフィールド(Wilder Penfield)や神経生理学者エックルス(J.C. Eccles)という偉大な脳研究の先達でさえ、晩年に心的エネルギーの存在を述べたり、心的第3世界への連絡脳(Liaison Brain)の存在を考えた。そして、今日の神経科学の知識では精神を脳の高次機能の所産として捉えることができないとの結論に達し、二元論的に心身問題を解決しようとした。更に、分離脳を用いて左脳・右脳の機能の研究を行ったスペリー(Roger W. Sperry)や皮質下レベルとくに脊髄で運動制御の生理学的研究を行ったRagnar Granitも、精神が物質的変化をもたらすとか、脳内ニューロンは目的をもって働いているというような表現をし、晩年は唯物論を離れて観念論の立場で主張した。上述の碩学たちはシェリントン(Charles Sherrington, 1861-1952)学派に属する人達である。シェリントンは「脳は多数の反射を有機的に統合して複雑な運動を作り上げる作用をもっている」というように反射学を学問として確立した近代神経生理学の基礎を築いた学者で、EcclesとGranitは愛弟子である。
丁度これに対立する立場に、唯物論の立場から「脳の反射」(1863)を発表したロシアの生理学者セーチェノフ(1829~1905、シェリントンの32歳年輩)がいる。彼はクロード・ベルナールの研究室で反射運動の中枢抑制機構の研究を行い、帰国後この著書を発表した。セーチェノフの研究上の指導原理はデテルミニスム(決定論:ビタリスム生気論に対する用語で、生物体においても無生物におけると同様、すべての現象の存在条件は絶対的に決定されていると考える立場)であり、脳の活動に反射原理をひろげ、動物およびヒトの心理活動の反射理論の発端となり、パブロフの高次神経活動の学説、すなわち、条件反射学説を生み出す道をひらいた。
以下に高次神経活動(=脳の高次機能)すなわち、精神活動を研究するに当たって現在、必づしも適正に評価されてはいない条件反射の有用性について考えてみようと思う。専門的に研究している訳ではないので、筆者らにとっても大胆な試論である。考察が表面的にならざるを得ないが、可能な限り誤りなく客観的な態度に終始したいと思う。
脳髄における反射の概念を最初に確立したのはデカルトであった。外界の刺激が神経の伝導によって脳に達し、その結果としておこる生物体の自動的反応を彼は「反射」と名づけた。その内容は機械論的観念論であった。すなわち、人間の理性的霊魂という原理をもちだし、人間の心理を反射概念によって説明しようという方向へ進めることはできなかった。後世の人たちの実験結果がつみ重ねられ、次第に科学的に理解されるようになるのであるが、セーチェノフ、シェリントン、パブロフと辿る反射の理論の変遷については他の拙論にやや詳しく述べられているのでこれを参照されたい(1999)。ここではただ一言。以下に述べる条件反射学説が心理現象を弁証法的決定論に従って研究しかつ理解する道を開いたという事実を指摘しておく。
脊髄レベルの反射弓を出発点としてこれの複雑な組合せ、またはその総合として中枢神経系の機能を説明しようとしたシェリントン流の反射理論は、大脳皮質の働き、すなわち、精神活動の領域においては無力であった。というより思考の脳髄に対する関係という問題は彼の生理学的立場からは、無意味(ignorabimus、われわれはそれを知らぬであろう)な解釈不能のものとみなされていた。シェリントンはパブロフの条件反射(理論)は「唯物論の香りがする」と言って認めようとしなかった。より正確には理解できなかった。
パブロフの学問上の師であるセイチェノフは、著書「脳の反射」の中で「生体は環境なくして生活していくことは出来ない。」「意識的、無意識的生活におけるあらゆる行動は、そのおこり方からみて、広い意味の反射である。」「感覚器の刺激感受性が停止すれば精神過程も停止するだろう。」と述べている。「脳の反射」が発表された1863年を己の研究の出発点と位置づけたパブロフは己の反射説の根本原理として「原因のない、動機のない、刺激のない作用は存在しない」と言っている。そして、彼はいかなる神経作用、いかなる精神現象も客観的に存在する実在性をもっていて-それらはこの実在性の反映である-、それに対して、客観的な生理学的研究の仕方を通じて、唯物論的な解釈を行うことができると考えていた。そして、「条件反射」を個々の生活のなかで獲得された生体の反射と定義づけ、「無条件反射」を「生体が生まれつき持っている一定の反射活動」「大脳皮質と関係なしに行われる」「遺伝的に無条件に固定されており、生存してゆく間に、条件反射として形成されていく色々の行動反応の基礎をなすもの」と考えた。
条件反射には、第一(次)信号系と第二(次)信号系がある。第一(次)信号系とは、直接われわれの感覚器官に感ぜられる直接的な刺激で、第二(次)信号は、第一(次)信号(単純な刺激)の符号化されたもので、新しい質の独特な信号、すなわち、間接的に表現される言葉である。第一(次)信号系を感覚信号系、第二(次)信号系を言語信号系とも言う。人間の場合には、第一(次)信号系の他に、他の信号系として言葉が追加される。言葉が特殊な信号系として追加されることによって、新しい神経活動の原理、すなわち抽象と普遍化が行われるようになる。
人間の大脳皮質の機能は、その構造を反映して動物のそれよりも非常に複雑化し特殊化している。従って動物実験によって得られた結論を人間にあてはめるにあたり、慎重であるべきなのは当然である。大脳皮質で生じる神経過程の基本的法則には高等動物(サルなど)と人間に共通するものがあるが、人間の大脳皮質はより高度に組織化されており、人間が誕生する過程で共同作業などのコミュニケーションを通じて言語が発生し大脳皮質内で知覚・認識の領域と運動・自発的能動性の領域が互に関連をもちながら発達する過程の中で人間の大脳皮質の連合野が著しく発達し、その特殊性が構成されてきた。
ここで、大脳皮質連合野と条件反射第二信号系(言語野)の関連についての問題が次の大きなテーマとなる。動物は外界からの感覚刺激を知覚し、認知する。皮質レベルで連合線維間のシナプスが替わるにつれて、一般に神経細胞はより複雑に細工され、処理された情報としての刺激を受けとるようになる。サルにおいて、後連合野内の特定の領域、すなわち、上側頭溝 (superior temporal sulcus)後部の周囲皮質(posterior STs 域) に体性感覚、視覚、聴覚などの入力が、一定のパタンをもって集中して終わっている。同様の皮質レベルの感覚刺激伝達の集中性は、ネコにおいても認められ、サルのSTs域とネコの中S上溝 (middle suprasylvian sulcus) 周囲皮質 (MSs域)が対応している。サルの脳にみられるこのSTs域は、視覚性、聴覚性および体性感覚性の性質をもつ各連合野に囲まれており、ヒトの大脳皮質の発達段階にくらべて、たとえ質的に "低次"で萌芽的であるとしても、皮質内の位置的関係や線維結合上の類似性からして、進化論的にみて、ヒトの39野(角回)や40野(縁上回)-感覚性言語中枢域(Wernicke中枢)- に発達し分化する可能性を内蔵している領域と考えてよいように思われる。この段階はパブロフの条件反射理論でいえば第一信号系から第二信号系への移行段階であるといえよう。
動物は符号/信号を用いてお互いの意志を伝達し合う。伝達手段として、単なる感情を主体とした伝達の上位に、身振り語、音声言語、文字言語がある。「あぶない!」とか「逃げろ」とかの言葉を情動的に発した場合、サルが相手であってもヒトが相手であっても、本能的行動ないし無条件反射的に関係した反応(一種の反射運動)を示すと考えられる。音声によって特定の行動への指示が与えられているように思われるが、音声によって情報が伝達されているからといって、サルやチンパンジーの「ことば」(使用しない方がよい!)が人間の言葉と本質的に同じであるとは考えられない。
いわゆるbody languageといわれる身振り言語は、①サルやチンパンジーの間、②外国語の初期学習期、③聾唖者間の手話、④幼児の初期言語習得期などにみられる。①~④でおのおの当然条件が違うが、共通している所は、身ぶりという信号は意味をもたせることが出来るので、「身ぶり観念」とでもいうべきものを作り出すことが出来るということである。これは身ぶり言語によって一般的表象が個体間で伝達される可能性を示している。つまり、サルが(または最初期幼児が)観念を形成し、かつ、それを伝達するという初期の段階に位置することを我々は知るのである。これは後連合野内の原始的言語野の形成過程に、進化的にみて(拙論-言語野皮質の発生-をみよ)、対応するものである。
この次にくるものは、音声言語を用いることによる観念の形成と伝達の段階である。観念を形成するということは、知覚像が音声記号のなかに客観化されて個人の心理過程から相対的に自立性を得て、持続性をもったものになることである。そのためには記号と結びついて客観化される必要がある。音声記号は、身ぶり記号とちがって、記号の任意性をもっており、あらかじめ個体間ないし集団(生活)内で約束をとり結んでおく必要がある。このような任意性をもつ記号を使用することはサルにとってむずかしい。殆ど不可能である。サルの脳は視覚的に三角形と四角形を判別したり、相手の表情を読むことは可能であるが、文字を認識することは不可能である。同様に聴覚的に異なる周波数の音を弁別できるが、情動に訴えない声の内容を理解することはできない。これを担当する(に照応する)言語野は発達していないからである。記号の任意性をもたない身ぶり記号から記号の任意性をもつ音声記号への発展の過程には質的な変化がある。サルとヒトの大脳皮質の間に認められる差、すなわちヒトの大脳皮質のもっている複雑性と特殊性がその変化を説明する機能・形態学的な基盤である。このような質的変化がヒトへの進化の過程で成立したのである。サルからヒトへの進化の過程で、ヒトは身ぶりではなく音声を媒介として観念を形成するようになり、更に具体的観念から抽象的観念を表すことができるという段階に発展するのである。この発展は概念の捉え方における質的な変化を意味する。すなわちこの抽象的概念とはそれについて一般的表象が成立しない対象についての概念である。どの民族の言語の歴史を調べてみても、聴覚連合野に依存する音声言語に次いで、視覚連合野に強く依存する文字言語が創造されてくる。音声言語に較べて文字言語が加わると、言語体系がより思考過程に深く入り込んでくる。文法構造が確立し、時称の区別、能動と受動と中動(ギリシャ語をみよ)の区別、断定と推定の区別、現実と仮定と希求の区別等々の文法構造が言語の発展過程で形成され、備えられる。思考・認識の方法も多様化し深化する。この段階に到達しているということは、脳の働きから考えて、後連合野と前連合野の中に広く存在する言語中枢が全体として活動していると考えられる。
このように、高次の符号である言語を媒介として抽象的思考の段階に進むためには、発達した前(頭)連合野(前頭前野)への入力が必要となる。この過程のなかで前頭前野内に能動的性質をもった言語に関する領野(Broca 中枢)が新しく形成される。概念を形成し判断を下し、推理を試みるという思考能力を備えた前頭葉皮質が形成され発達してくる。こうして、言語が特殊な信号系として追加されることによって、新しい神経活動の原理、すなわち抽象と普遍化が行われるようになる。すなわち、質的に発展を遂げた高次の言語条件反射といわれる第二信号系が形成されるのである。
子供にみる言葉の習得・発達をみるに、サルからヒトへの進化の段階で言語を獲得していったであろう過程が類似していることに気づく。言語の習得と概念形成上の使用を説明することに、パブロフの発見した条件反射の原理が適応されている。児童期から思春期にかけて、先にみたようにヒトの脳髄は完成する。言語中枢の発達と聴覚・視覚などに代表される感覚連合野および能動機能を発現させる前頭前野と密接な関連をもった大脳皮質の成熟は音楽や美術の鑑賞と表現を高度なレベルにまで高める。児童期から思春期における脳科学の進歩を採り入れた科学的な教育方針が文化、芸術、科学、一般社会の福祉などあらゆる分野で現在求められている。われわれは条件反射の理論を学び、求められている上記の分野に創造的にこれを適用する努力を怠ってはならないと思う。
脳の正常発達と教育との問題に関連して、性格異常、社会的ストレスの影響、ノイローゼ、精神病の発病と予防・治癒への科学的アプローチについて、晩年のパブロフが彼の条件反射理論を動物からヒトに拡大して精神医学の分野で如何に貢献しようとしたか。この点できるだけ具体的に考察してみたい。
以下に、学童・青年の教育の問題と精神異常の問題について条件反射理論を武器に掘り下げてみたい。
人間は、いかにして言語を獲得したのか?ネコ、サル、ヒトの連合野の発達、とくにサルの後連合野皮質における種々の感覚(視覚、聴覚、体性感覚など)の集中性とその領域のヒトへの感覚性言語中枢への発展については、脳の皮質皮質間結合の研究(川村、1970~)で明らかにしてきた。脳内に符号(信号)を理解する領野、ついで符号の最高のもの、符号の符号、つまり言語を理解する領域がヒトになって発達するのである。言語の発達が示すように、例えばサルの生活の内での感情的呼びかけが身ぶりと一緒になり、反復しているうちに集団の内である程度のとりきめが発生して誕生するのである。つまり、認知/認識の理解とその伝達表現には、その言語発達と共に、emotion(情動)の表出が、伴っているのである。下層から言語伝達を支えていると言ってもよい。脳の構造とその発達は大脳辺縁系と密接に関連しながら、その旧い皮質の上に発達した新皮質とその最も発達した連合皮質内の言語野の発達とその過程を系統発生的(進化論的)にみることが出来る。
ペダンティックの謗を免れないが、言語起源論の最古のものは、神(創造主)が与えてくれたとする言語神授説であろう。旧約聖書には「エホバ神、土をもて野のすべての獣と天空のすべての鳥を造りたまいて、アダムのこれを何と名づくるかを見んとてこれを彼の所に率いいたりたまえり。アダムが生物に名づけたるところは皆その名となりぬ。アダムすべての家畜と天空の鳥と野のすべての獣に名を与えたり。」と記述されている。
これに反して、古代ギリシャの唯物論者たちは、言語の起源が「人間の自然的本性」(エピクロス)であるとし、また、その流れをくむ哲学詩人ルクレティウスは、「万物に名を与え、最初の言葉を教えてくれた発明者がいたなどと信ずるのは馬鹿もよいところだ」(「物の本性について」)と、言語の起源を神に求める考えを一笑に付している。
時代はずっと降るがダーウィンは種の起源(1859)でうち出した進化論を発展させて、「人間の由来」(1873)の中で、人間への進化も、他の動物間と同様、その心の諸能力も含めて、動物との連続性をもっているとして、「私は、言語はその起源を、いろいろな自然の音、他の動物の音声および人間自身の本能的な叫びを、手まねや身ぶりに助けられて模倣し、修正したことに負っている、ということを疑うことができないのである。」
「人類の初期の先祖が音声を最初にもちいたのは、おそらく現生のある種のテナガザルがやっているように、真の音楽的抑揚を発すること、つまりは歌うことだったろうということが明らかになるであろう。広範囲にわたってみられる類例から、この能力は雌雄の求愛期間にとくに用いられ――愛、嫉妬、勝利などのさまざまな感情を表現したことだろう。――また、競争相手への挑戦としても役だったろうと推論できるのである。したがって有節音によって音楽的な叫び声をまねることが、いろいろな複雑な感情を表現する・ ・ ・ことばになったのだということは、ありうることなのである。」
と述べ、人間と動物とは連続的であり、その差は程度の問題にすぎぬと主張する。ダーウィンの言語の起源に関する見解もこの域を出ていない。
また、ジャン・ジャック・ルソーは、「情念」を言語発生の原点と考え、次のような言葉を残している。
「言語の起源は人間の最初の欲求に負うものでは少しもない。…というのは、(欲求という)人びとを遠ざけている原因から、(言語という)人びとを結びつける手段が出てくるというのは不条理だろうから。それでは一体、この起源はどこから発しているのだろうか。精神的欲求、つまり情念からである。あらゆる情念は、生きようと求める必要のためおたがいに避けあわざるをえない人間たちを近づけるのである。彼らに最初の声を出させるのは飢えでも渇きでもなくて、愛であり、憎しみであり、憐れみであり、怒りである。」つまりルソーが主張したのは、対人関係における欲求・感情に結びついたコミュニケーションであった。
マルクスとエンゲルスはダーウィンの進化論を更に発展させた。「意識的な生活活動が人間を動物の生活活動から端的に区別する」(「経済学・哲学手稿、マルクス」)と人間と動物の本質的な差違を指摘している。誤解を避けるために付言するが本質的差違は意識の有無であると言っているのではなく、生活の中で意識的に社会性、共同性をもって活動をするかどうかという点にある。この生活活動のうちで、基礎的、主導的なものとして、労働の役割を捉え、労働こそが人間が動物から自らを区別して人間化していく最初の推進力であるとしたのである。この労働活動こそが、人間と動物の連続性における飛躍の原動力となるものであるとの観点が、後年エンゲルスによって「猿が人間化するにあたっての労働の役割」(1876)という自然弁証法の中におさめられている小論文として結実された。この著作は、進化、労働、言語などの関連問題を多面的な角度から解明しており、現在、高次機能に関心をもっている脳研究者を含む科学者にとって是非とも読まれるべき労作である。実際、これまでみてきたように後連合野の働き、言語中枢の発達、運動系・能動系皮質の活動など、これらの相互関連が現代神経科学の広範囲な分野の内で解明されて、上述の科学的見解が実証されてきている。
次に幼児、児童の脳の発達について学習意欲と教育について考えてみたい。上述したように根本的には、欲求は脳幹や辺縁系皮質が関係するもので、それが脳幹網様体賦活系を通して大脳皮質の後半部さらに前半部に活動を与える。その結果、高次元の欲求、意欲に昇華・発展させ、方向づけ、意識的で計画的な行動に変換させ、前頭前野の働きを以て、行動を意図し、計画し、実行し、評価する。高次元の意欲とは、このように、脳幹・辺縁系の自然的欲求と後連合野の認知体系の活動に対して、前連合野からのフィードバックが働いて、その能動的な働きにより制御(コントロール)されて再編成が行われるという高次脳活動の現象である。このような経過をとって子供の精神発達がなされる。この精神発達にとって重要なものに、神経膠細胞(オリゴデンドログリア)が神経興奮活動を円滑ならしめるために神経回路を構成する神経突起(軸索)のまわりを絶縁体の鞘で被覆するという髄鞘化現象がある。この髄鞘化は生後漸次完成するものである。新生児では、第一運動領や第一感覚野では髄鞘化は完成しているが、後連合野や前頭葉ではその発達が遅れていて髄鞘化はみられない。6ヶ月になっても前頭前野の髄鞘化はほとんど進行していない。3歳児の脳で後連合野の髄鞘化は中程度、前頭前野は初期段階である。7歳児では、後連合野でほぼ完成、前頭前野は中程度。思春期にまでかかって大人の髄鞘化された脳が完成すると言われている。
脳の発達には髄鞘化の他に神経細胞(ニューロン)の樹状突起の発達、ニューロン間の互いの連絡すなわち回路網の形成、条件反射に基づく伝達効率が進んだ機能的連合回路の発達、それに、カテコールアミン系(ドーパミン、ノルアドレナリン)、インドールアミン系(セロトニン)の大脳皮質への入力と樹状突起へのシナプス形成と過剰入力へのelimination, trimming(除去、トリミング)、皮質と皮質下との構造物間の刺激と抑制のバランスの確立、それに磨きをかけるかのように画竜点晴とも云うべき言語中枢を含む認知機能、能動性発揮の機能と情動機能との有機的な連携が、適正な外界からの刺激によって情と意と知の豊かな成熟した発達が青年期までの長いスパンで緩やかに形成されていくのである。この発達期に情操を豊かに育てることと子供を“プラス”の行為に動機づけることが重要である。それには、「理解されているだけの動機」を「実際に有効な動機」に転化させていくことが大切である。子供は大人が傍らにいるだけで、子供の自己中心言語は増え、また子供の問題解決の成績は向上するという。ヴィゴッキーは次のように説明する。「子供は本来社会的存在であるから、その社会的関係の中に置かれたならば発達課題をものにし、大人の助けを借りて達成していくなかで自分のものにする。」そして、彼は「子供が現在いる関係領域よりもより高い発達段階で、子供が占めることのできる関係領域」を「発達の最近接領域」と呼んだ。この際、社会が、大人が子供に要求する課題は、子供の発達の「最近接領域」に属するものでなければならないと主張し、そこで生じた動機は理解され、課題に取り組んでいるなかから、実際に有効な動機が生じてくると言う。社会的環境の内で子供の意欲を発達させ、その意欲を意識的に行為に作りかえるには、ルリアによれば言語系である第二信号系による行動の自己調節的な働きが重要である。その時期は、子供が自己中心言語の時期を経て、言葉を自分の思考の手段として表現できるようになるのは、内言語を使用することができるようになる6-7歳の時期であるという。内言語とは外言語が内化したもので、この時期はその移行期に当たる。その後、12歳頃までは言語が次第に論理的になり、計画的行為も具体的事実と結び付けられるようになる。その行為が具体的な操作から一般的で形式的な操作の段階になるのは児童期の後期に相当する。具体的なやり方を経験するなかで、抽象的な解決法が生まれてくる。このことは数の概念の把握の仕方をみれば明らかで、具体的事物に置き換えるという操作を通じて、抽象的な概念が後連合野内に形成されると思われる。これが運動系皮質を活動させると学習課題にとっての計画性をもった行為として表現されることになる。このような計画的活動が反復されて、行為は一般化され条件反射第二信号系の働きにより意識的なものとなる。この段階で「行為への意図」を意志作用として定められるようになる。ここでは具体的行為は言語的に正しく評価されるようになる。この段階は前頭前野(帯状回前方部も含めて)の活動が主体となる。この意志作用は内言語の助けなしには成立しない。次に青年期(思春期)に移っていくが、行為の主体である己を客観的に対象化し、言語評価が与えれらるようになる。いわゆる「自我」が確立されていく過程に相当する。
以上の生後から青年期に至る(正常の)神経発達過程に障害が起こると様々な神経症状や精神症状が現れる。ところで精神障害の原因究明が叫ばれているが、その一つの立場として、胎生期に始まる神経発達の障害仮説が提唱されている。
分裂病患者の死後脳について、観察され、研究結果が報告されているが、母集団を均一化することがむずかしいためか、所見が不確定で定まっていない。ヒトを対象とするこの種の研究はすべてが一致して、動物実験結果のように、再現性を求めて結論づけると言う性質のものではないことは万人が承知している所である。というのは、精神症状の縦断的経過、遺伝負荷要因などの多様性があるのが精神分裂病の特徴で、普段は一見正常者と言動にさしたる差違がみられない。また、思春期に多くが発病すると言われているが、突如として、興奮、幻覚、妄想、などの人目を驚かす症状が出現するのではなく、それまでには、学童期、場合によっては幼児期に周囲から「チョット変ジャナイカシラ」と思われる程度のいわゆる前駆症状がみられるものが大部分である。
死後脳で、側脳室や第三脳室の拡大、皮質体積の減少、海馬の変化[縮小、神経細胞の大きさの減少や配列の異常、形成異常、シナプスマーカー(synapsin, synaptophysin syntaxin, SNAP-25)や樹状突起マーカー(MAP2) ]および前頭前野、帯状回、内嗅野などの皮質にみられる海馬の変化と同様な病変が数多く報告されている(cf.車地、2000,review臨床精神医学29/4.;381-389)。
以上の病変に、一般にグリオーシス(炎症反応の時にみられるグリアの増殖)の組織変化が認められないことから、グリアの発生する以前のニューロン産生や移動の時期に起こる胎生期の発達障害説が提唱されている。しかし、よく調べてみると(病理学者に聞くと)、生後の変化で、とくに海馬などではグリオーシスを伴わない病的変化がしばしばみられるので、完全に納得される説明とは言えない。とにかく、精神病の発症を解明することは胎生期、幼児期、児童期、青年期における脳の発達を研究することに直接連がっている。この際、神経要素(ニューロン、グリア)、栄養因子(分子)、神経細胞接着因子(NCAM, L1など)、神経成長因子、脳の発育に関係する遺伝子(転写因子、シグナル伝達因子、シナプス形成や神経回路形成に関するものetc)の動態を相互に関連づけて動的に研究することが重要である。さらに、神経回路の問題では、機能的に異なる大脳皮質の諸領域が如何に連合線維によって結び付けられているかということを単に知ることにとどまることなく、人間の脳と精神の問題を研究する者は、ヒトをヒトたらしめる唯一の特質である言語(ロゴス)中枢の問題を情動(パトス)や意欲に関連する(脳内)領域(これまでに再三述べてきている)と結び付けて、物質の言葉で具体的に語っていかねばならぬ責任を負うであろう。音楽家、美術家は、己の脳内で活動するロゴスとパトスを聴覚系ないし視覚系の連合野における認知・認識系(ヒエラルキーを備えている)と連合線維(皮質皮質間結合線維)を通じて機能的に結合を強化し、前頭前野の能動機能を美しく“人間的”に発揮させようと努力し、練習に励む。この機能的連合結合の形成は、上述してきたように、条件反射の機構に基づくものであり、芸術の分野でも、教育の分野でも脳研究の分野におけると同様、条件反射理論を各分野で評価して応用していかねばならないと考える。
最後に、これまでの記述で欠けている重要な点を考察の対象に選び、この稿をまとめたいと思う。それは前にクレッチマーの項で現象学的に触れているが、気質、性格またその異常の問題である。それに関連してパブロフのイヌの性格と実験的神経症の研究と、それを基にしたヒトの神経症(Neurose)の考察、更に晩年のI.P.Pavlovが関心をもった精神異常/精神医学(ヒステリー、強迫神経症、分裂病など)への科学的とりくみについて話題としてとりあげたいと思う。
先ずパブロフのイヌの実験神経症に至る過程について述べる。パブロフの高次神経活動の「型」の理論は、1921年頃から晩年に至るまで長期にわったって展開された。彼はその活動の基本的特徴を3つに大別し、それに基づいて神経系を型によって分類した。第一は、興奮、制止という神経過程の<強さ>(the force)、第二は、こうした過程の<平衡>(equilibrium)、第三は、それらの<易動性>(mobility)である。強さとは、環境が強いる緊張に皮質細胞が耐えるという特性である。平衡とは、興奮と制止の過程に同一の強さと易動性を維持する特性である。易動性とは、興奮と制止の過程が環境の変化にこたえて変化する特性である。そして、パブロフはイヌの行動パタンにもとづいてこのような神経系の3つの組合せから、4つの神経系の型に分類した。すなわち、強くて不均衡な(抑えのない)タイプ、強くて均衡がとれ、敏活な(活発な)タイプ、強くて、均衡がとれ、緩慢な(落着いた)タイプ、そして弱いタイプである。これらはそれぞれ、胆汁質、多血質、粘液質、憂うつ質(黒胆汁質)と対応している。このタイプは、気質の問題として、人間存在の天才的観察者であったギリシアの哲学者(ヒポクラテスをはじめとして)たちが、経験主義的に“芸術的”に、4つに分類したタイプに近いものである。
ここでちょっと重要な割り込みをさせて頂く。パブロフの実験的神経症を評価して紹介するなかで内村祐之(精神医学の基本問題、医学書院、1972)は精神障害発生に対する二つの伝統的思考として以下の文章をわれわれに残している。すなわち、
「この二つの思想の第一は、人間の精神機能は階層的構造をもつというものであり、第二は、精神機能は生理学的反射機構と深い関連をもつというものである。神経系統の機能、また心的機能が、未成熟の段階から、種々の発達段階を経て、最終的に成人の成熟した段階にまで達するという進化論的見地は、個体発生史または系統発生史を注意深く観察し、しかも自然科学的に考える人々にとっては自明の理である。そしてこれはまた近世の精神医学者の誰しもが、精神異常の症状を理解しようとする際に、脳中に描いた前提であったと思う。このことを具体的に記述し、しかも精神異常の解釈を、この進化の反対方向、つまり逆行の結果として捉えようという試みを最初に敢行したのがH・ジャクソンであったことは、前に詳しく述べた通りであるが、その解釈のニュアンスや内容こそ違え、異常な精神症状と低位精神機能とを結び付けて研究を進めた研究者はジャクソン以後にも多かった。本展望で紹介した研究者の中から拾っただけでも、クレペリン、ジャネ、フロイト、クレッチュマー、エイなどの名が挙げられる。層次論と退行論とは、精神病構造論を貫く一つの中心的思想である。
次に、第二の思想たる、反射と精神機能との関連であるが、これは、脊髄反射の生理学的機構の知見にヒントを得て、複雑した精神現象もまた進化した大脳の反射作用として理解できると考えたグリージンガーに始まり、失語症の発見から精神的反射弓の作業仮説を発展させ、これによって精神症状の発生を理解しようとしたウェルニッケを経て、最後に、条件反射の発見に基づき、精神機能とその異常もこの条件反射の枠内で理解できると考えたパブロフに至るまで、長い道程を経た思想である。」
話をパブロフの「実験的神経症」に戻す。パブロフの研究グループは条件刺激を漸次困難なものにして、イヌを葛藤やストレスの状況下におとし入れるという実験を行った。たとえば条件刺激として円を示した時に唾液分泌が起こり、長楕円では分泌が起こらないように条件付けをしておいてから、円と楕円を少しずつ似たものに近づけていくと、ある点で、イヌは二者を弁別できなくなり、混乱し、「神経病」の状態を起こす。この際、イヌの行動を観察して明らかになったことは、「神経症」を起こす条件は動物の個性/性格(行動の型として現われる)により左右されるということである。
パブロフは神経系の型のタイプを人間にあてはめる際には、非常に複雑であるとして、極度の注意が必要であると考えていた。この複雑性は人間の環境が社会的な性格をもつことからくる。つまり、神経系の3つの特性-強さ・平衡・易動性-は、人間の型の決定にも適用されるが、それが社会的環境のなかで形成される。くどいが、もって生まれた神経系の特性の上に-それよりも基本的な意味をもつものとして、性格/気質のもつ社会的価値、期待、困難の克服、組織への忠誠、問題解決の知識や実践能力、生活上の打撃や緊張など複雑である。
以上のように洞察して、パブロフは、人間の神経の型を決定する二つの新しい基準を導入した。すなわち、人間の型の問題は強さ、平衡、易動性に加えて、一方では皮質と皮質下の活動の間の、他方では第一信号系と第二信号系の間の、相互関係の特殊性という面からも研究されなければならないとした。要するに、人間の型を決定するためには、ある個人が受けたすべての影響が考察されなければならないのである。パブロフは、人間の神経系を非常に一般的な三つの型に分類する。<生活経験のなかで>、ある人々は一方の極として第二信号系よりも第一信号系をよりよく発達させ、他方の極として第一信号系よりも第二信号系をよりよく発達させる。あるいは、ほとんど大部分の場合がそうであるが、言語系に調整者の役割を果しつつ、二つの体系の、均衡のとれた相互関係を発達させている。人間の型に関するこの理論を理解するためには、われわれはさらに、両信号系の性質と両者の関係とを研究しなければならない。問題の中心は情動と思考の性質と役割にある。
情動と観念およびそれらの相互関係の基礎にある高次神経活動は、三つの体系を含んでいる。それは、皮質にある感覚系(第一信号系)と言語系(第二信号系)、皮質下にある無条件反射系である。観念はつねに言語による抽象作用であるので、第二信号系あるいは言語信号系が観念の基礎をなす神経活動となる。他方、情動は、主として第一信号系あるいは感覚信号系と皮質下(扁桃体、中隔核、分界条底核、側坐核/腹側線条体を含む大脳辺縁系を構成する部分)の無条件反射系の相互作用である。このように観念(認識の機構)と情念(情動機構)の基礎にある神経回路を含む諸々の過程は、互いに密接に関連している。このことは、これまで解剖・生理学的に詳しくみてきたところである。繰り返すが人間にあっては、認識(観念)と情動は、実在を意識内に反映する不可欠、不可分の二側面である。その意味で、動物と違って言語中枢を備えている人間にとっては、情動は意味をもっており、<観念なしに情動はなく、逆に情動のない観念もない>。
晩年のパブロフは精神病院での研究を通して強迫症(obsessive)、妄想症(delusional)、心気症(hypochondrial)、抗うつ症(depressive)など種々のタイプの精神異常(神経症や精神病)の患者を分析的に診断した。われわれは、これら精神疾患の分析研究の成果を、パブロフ自身の詳しい報告を、読んで吟味することができる(高次神経活動の客観的研究、I.P.パブロフ著、岡田・横山訳、岩崎学術出版、1979)。彼は、高次神経活動の諸法則を利用して、それらが第二信号系にあてはまる場合の特殊性を証明するように努めた。パブロフは約15年にわたって実験神経症と精神病の研究をしたが、それを通して、有効な治療法について探求し、彼の好んで口にしていた「科学的に健全な心理療法」に向かって歩を進めたいと思っていた。今日の精神医学者(臨床医と研究者は不可分で共に)はこれを受け継ぐ用意をしなくてはならない。現在脳科学は大きく進展し高度の知識をわれわれは持っている。この財産を国民共有のものとして、芸術、教育、医学の分野に科学的な方法を生み出して適合させて、豊かな芸術、正しい教育、安心できる信頼される医療を築いて行かねばならない。
12]脳と精神の科学の発展と思春期における自我の形成と情操教育
18世紀末にピネルが鉄鎖から開放するまでは、精神病者は正当な人格も認められない、悪霊にとりつかれた罪人のように扱われていた。精神病が脳の病気として自然科学的方法をもって医学の研究対象となったのは、グリージンゲル書の「精神病の病理と治療」(1861年)以降といわれている。当時の自然ならびに社会科学的事項をピックアップして時代背景をみてみよう。この10年前後の間にダーウィンが「種の起源」を出版し、パスツールが生物の自然発生説を否定し、メンデルが遺伝法則を発表し、マルクスが資本論第1巻を刊行している。考えてみると、価値観というか世界観の変革をせまられたすごい時代であった。
神経の回路網や構成についてはかなり明らかにされてきたが、脳病の成立機序が解明されるのは21世紀であろう。あらためて歴史をひもといてみる。1838-39年に、シュライデンとシュワンによって細胞説が確立され、その20年後にはウイルヒョウが「細胞病理学」なる大書を著わしているが、精神障害の身体的基盤が信じられるようになったのは、クレペリンが、他の身体疾患と同じく、精神病にも原因、経過、転帰、病理解剖の同一性をもつ疾患単位があると考察した1883年(「精神医学書」を出版)頃からで、この頃には神経細胞(ニューロン)の概念もほぼ固まりかけていた。
19世紀の遺産をひきついで、20世紀前半には輝かしい成果が産まれた。パブロフの大脳皮質の働きを中心とする条件反射理論、カハールのニューロン説、シェリントンの神経生理学、モルガン一派による近代遺伝学の基盤の確立、シュペーマンによる形成体の発見、ベルガーによる脳波の導出、ヤスパースによる精神病理学の方法論の確立等々、神経学生物学の領域に限っても数え挙げれば切りがない。今世紀後半になり生命科学に対する関心が高まり、ワトソン・クリックによる核酸 DNAの二重らせん構造説(1953年)に始まる分子遺伝学の発展成果を神経発生生物学をはじめ脳の科学に結びつけようと、それぞれの専門分野で大きな仕事を成し遂げてきた理論物理学者や分子生物学者達がこの神経科学の分野にどっと流れ込んできた。非生命体を対象としてきた物理、化学の成果を存分に取り入れて、物質によって構成される生きた細胞の進化の産物である有機体(=生物)を対象として、それまでになかった分子、遺伝子という物質の言葉を用いて基本的な仕組みを語れる研究を進めることができるようになった。因みに、「免疫」、「癌」の研究の次に来るものとして、21世紀は「脳の世紀」と位置づけられている。
音楽、美術においても、19世紀から20世紀にかけて、価値観の変革と創造をせまられる時代であった。人類はかつて経験したことのない第一次、第二次の世界大戦という精神社会の荒廃と頽廃を身をもって体験する。不確定要素に囲まれた時代を過ごしている。以下に、芸術面よりの概括的な記述が欲しいところである。(丸山先生に考察をお願いする)
この項では、脳の活動の所産である精神活動と複雑な社会の中で能動的に生活する、社会的動物である人間の発達について、人間を人間たらしめる自我の形成時期である青年期(=思春期)の精神活動にだぶらせながら、解析を試みることにする。
上の図はこれまでの記述をまとめたものである。大脳新皮質と大脳辺縁系に代表される2つの大きな高次機能である受動系としての認知機能と情動機能、それに能動系としての運動と意志の発動がある。これらの高次神経活動の総体=精神を下部構造が支えている。それらは、覚醒・睡眠リズム、歩行リズム、昼夜リズムなどの自動調節システムである。複雑な神経回路網が完成されて精緻に脳が活動するには、これらの脳幹レベルに存在する諸々のリズム発進の機構の他に、縫線核由来のセロトニン系(5-HT)、青斑核由来のノルアドレナリンあるいはノルエピネフリン系(NAあるいはNE)および腹側被蓋野/黒質由来のドーパミン系といった視床下部、大脳皮質など脳の広い範囲にdiffuseに分布する神経伝達物質のシステムがある。勿論その他に興奮性アミノ酸や抑制系のGABA(ギャバ)、それに知能活動に関係のあるアセチルコリン系(Ach)がある。大脳皮質後連合野で外界からの刺激を知覚・認知・認識する一方、他方で感情・情緒的に外界からの諸刺激が、扁桃体でその意味ないし価値(益か害か、プラスかマイナスか)が判断される。それら受け入れた全体のものが知的に、意志的に、感情を込めて、運動系と感情表出系を介して外界に出力される。前者は主に運動皮質・線条体系で、後者は視床下部・下垂体系である。大脳新皮質とくに前頭前野が関連する最高次の機能については別に論じた。ここでは多数のアミン、ペプチド、ホルモンなど液性系が関与する感情表出系に焦点を当てて、その機能について発達論的に論を進め、そのメカニズムについても考えてみたい。人間の精神発達と関連させて、思春期における自我の確立という特別な視点から光を当ててみたい。
ところで、精神分裂病を含めて、思春期(広義、狭義により幅があるが、11才~15~18~22才)に発病する精神疾患は多い。考えてみると、人間の生涯を通じて、思春期は身体的にも精神的にも発育・発達の変化が最も著しい時期に相当する。まづ、身体的側面では、第二次性徴(男子で陰茎、喉頭軟骨、声変わり、陰毛など;女子で乳房、陰毛、初潮、脂肪沈着など)として特徴的な変化が認められる。11才頃から15才頃までに生ずるこの身体内の著変は意識的にも無意識的にも、当然自己の精神面に大きな影響を与える。一言で云えば、この身体的エネルギーの内的な緊張を伴う急増に対して、どう対処するかが問題となる。物質的な基盤としては、上述の神経伝達物質からホルモンを含めた自律神経系、情動系、液性系を主体に包括される変動と言えよう。心的活動の微分係数が突如として大きくなる。外的および内的刺激に対する過敏性(Irritabilität)が認められ、情緒が不安定になる。他者との相互的な関わり合いの中に置かれている人間は、この青年期(=思春期)になって初めて、両親・教師への従属的な存在から、未成熟さを自覚し、過度の自意識をもって周囲に対して過敏に反応しながら自己の存在を確認しようとする。この時の脳内機制を考えてみる。すなわち、感覚野と知覚連合野のニューロンは感覚刺激を処理する。そして、その情報を前頭葉に伝達する。その過程でまた前頭前野で受け入れる過程で、外界の刺激の価値判断を行う扁桃体を中心とする辺縁系と情動の表出を行う視床下部を中心とする感情出力系の二大システムを、大脳新皮質連合野が調節・コントロールする機構が発達する。この新皮質とくに前頭連合野から辺縁系や間脳領域に送られる制御・調節系が適正に形成されることが大切で、このシステムの主役である前頭前野(PFC)が成熟するのが(髄鞘化を含めて)丁度、思春期に相当する。この発達が障害されるといわゆる思春期の精神異常の障害としてみられる症状が現れる。そこには青年独特の攻撃性、反抗性、思い上がりがみられる。そしてそれは、社会から容認されるような形の判断基準(価値観)を身につけ、大人の社会からその存在が容認されるものとして自覚的に定められたものとはなっていない。そしてそこには精神医学あるいは心理学でいわれる両価性ambivalenceが顕著に認められる。これは知性の面でも情動の面でもみられるが、とくに後者で明らかに認められる。いわゆる正と負という価値の対立した概念が同一個体の中で同時に存在している。すなわち、尊大さと劣等感、依存と自負、尊敬と反抗/侮蔑、愛情と憎しみ、純愛と肉欲愛といったものである。この両価性は分裂病における第一級症状の一つである。思春期の障害としてはこの情動面の動揺や変化に対応できず、安定した基盤をもった自我が確立できず、精神分析学でいう社会的同一性(social identity)の形成が未熟な段階に留まっているが、社会は大人使いをする。
このように思春期=青年期は、自分の同一性identity,自我を形成する上で、精神形成の上で恐ろしく重大な時期である。他面では、家族や小学校で教えられた価値観が社会との接触を通して、矛盾に直面し、妥協を強いられ、必要悪を認めねばならぬことになる。親や教師に従属的に規制されていた独自性に欠けた「良い子」が、ここで試練に直面することになる。社会からはじめて主体としての営みを求められるのである。この時期における社会との言語および行為を通じての対応やその処理の仕方が、独立した個人の責任として求められるので、社会環境が悪くて混沌(カオス)として乱れているときには、その直接の影響として蒙るものは青年期の人格形成である。倫理的、道徳的なものを含むが故に、前頭前野の健全な働きが問題になる。
生物体の日常行動は健全なリズムの上に通常成り立っており、このリズムが大きく偏位するときには身体的、精神的異常が現れる。昼夜リズムの障害は、視床下部を中心とする気分の障害やうつ状態を出現させる。睡眠・覚醒のリズムの障害は生命機能が宿る間脳から延髄までを含んだ脳幹の機能調節の変調で著しい身体への疲弊と精神の不安、焦燥感を伴う。近年、薬物依存の障害で、覚醒剤中毒や睡眠薬中毒(ひいては抗精神病薬依存)が問題となっているが、これらは脳幹レベルにおける神経伝達物質とその受容体と液性伝達系、自律神経系、ひいては免疫系の障害として捉えることが出来る。薬物依存の問題は、人格障害、性格異常を引き起こし社会犯罪とも結びついてくる。また、広くリズムの障害は食欲(拒食や過食)や性欲の行動異常としても現れるが、これらの機能中枢が視床下部に存在していることと関係している。とくに摂食障害は思春期の疾患として重要である。
先にみたように心身両面に顕在化する性衝動の発現は思春期に特徴的で(そのまま)自我に同化しきれないもの、自分ではどうすることもできなものと感じさせられる厄介なものである。とくに、直接的に性衝動を行為化することがタブーとされている近代社会の中ではその行動は規制される。性犯罪との関係は教育と結びついて考察されねばならない。
これらを全体的に調節しているものは、前頭前野を含む大脳皮質から辺縁系へのregulationである。それと同時に、脳幹レベルから上行性に脳の上位中枢に広範囲に影響を与えている、ドーパミン(DA)、セロトニン(5-HT)、ノルアドレナリン(NA)のカテコール/インドールアミン系の液性のregulationがある。思春期は上記の事柄が脳の機能としてunreifからreifの状態に完成していくプロセスの時期である。健全な脳機能の発達という視点から考えて、この時期における身体的並びに精神的「栄養」の授与は大切なことである。
この「栄養補給」をどのように与えたらよいのか。ここで、筆者ら、すなわち脳を研究する科学者とキリスト教思想史を研究する音楽家が、成熟形成期の脳の所有者の身体と精神が生活をする上で青少年非行や学校教育や社会全般の問題について考えてみたい。
上述したように、脳の最高部位中枢である前頭前野が、下部構造をコントロールしながら活動する過程が、発達心理学的に自我形成・自己同一性の確立に一致する。とすれば脳を中心に考えてみても、思春期は内的にも外的にも疾風怒濤(Sturm und Drang)の時期である。心身共に変動する時期に、家族における親子の関係、学校における教師・生徒の関係の価値観が動揺し、時には崩壊し、再編を迫られる。少年期から青年期へと発育するにつれて、自我を確立すると同時に人間は社会的存在であることを自覚し、「私」の中に「私たち」を持たねばならぬようになる。つまり、人との交流、社会での常識や集団の中でのルール(これは家族内、学校内の規制/ルールの延長であるが、より発展したgeseligなものである)や責任を身につける。「共同体」こそが自己を確立する媒質であることを認知するのである。脳の後連合野の発達段階から前連合野の発達段階に発展・成熟し、内省の基盤である言語機能が完成に近づく。そして「内省」により「内面の他者」を発見し、自我の形成が促される。
いつの時代もそうであったように、現代の社会の環境は教育をめぐる状況を含めて深刻である。つめこみと競争の激化、不登校や「いじめ」、校内暴力、少年犯罪と教育環境は憂慮されるべき状態にある。この中で、生徒の目標を自分の成績順位にかけて争うという競争の場に教室をしてしまった管理、選別の教育システムが行われている。また、偏差値という単純な形で青少年を選択するから、彼らは勉強することに興味が湧かず、その意欲も欠如するようになる。こうして「学校に行くに値するのか」、「この世の中は生きるに値するのか」と言う彼らの問いに社会は益々答えられなくなってきている。現在、学校崩壊や少年犯罪が問題になっている。この原因は、少年の素質や教師の質の問題としてのみ捉えるのではなく、社会環境の問題として捉える必要がある。20世紀には、かつてない犠牲をもたらした二度の世界大戦があった。民族の抑圧、ファシズムの暴虐、化学兵器・核兵器の出現による大量殺人、侵略と虐殺などの戦争の違法化に対する人々の良心は麻痺してしまった。
大戦後も局地戦争は世界の各地でつづいている。モラルを説く大臣が倫理的問題を起こし、現代の社会は、長時間労働が野放しにされ、リストラと大量失業、貧富の格差、環境破壊等々多くの矛盾を抱えている。罪を犯した少年への教育的、福祉的措置を中心にした対応をしなければならないが、この犯罪の原因の多くは少年をとりまく社会環境の拙悪さや少年が未熟な発達段階にあることに端を発している。加害者である子供たちは、過去において学校で「いじめ」や体罰を受けたり、虐待にあったり、被害者である場合が多いことも事実である。厳罰主義では少年犯罪は無くせない。子供たちを追いつめているのは大人の責任である。
青少年期に彼らは、自・他の関係の不協和に悩み、自分を表に出さない傾向を保つ。他の「優等生、よい子」に対して無視・「切り捨て」・「いじめ」をする一方、自分としては「優等生、よい子」でいなければならないという矛盾を持つ。己の欲求や不満を表すことができずに、常に本当の自分でなく仮の自分であり、ずっと張りつめた状態を続けている。この精神状態が続けば、不安、焦燥、強迫心性、不眠、吐き気、自律神経失調症に陥ることは確実である。
最近、西鉄高速バスジャック事件、豊川の主婦刺殺、秋田の母親絞殺などの少年犯罪が頻発しているが、これらの犯罪行動には、さまざまな社会環境のゆがみが映し出されている。これらの環境を改善し、整えていくのは大人を含む社会全体の責任である。身体的には、脳幹の液性伝達物質の調節障害、大脳新皮質から辺縁系や視床下部へのregulationの障害を思い起こさせる。精神的には青少年自身に自分の犯罪行為の重みを自覚させる教育と同時に医療の充実が必要である。
以上にみた現代社会の害毒を取り除くにはどのような「栄養」をどのように補給しなければならないだろうか?そして、その方策は?近年、わが国においても教育改革が叫ばれており、「奉仕活動」、情報技術革命(IT革命)、体罰、効率的学校統廃合などが提唱されているが、それでよいのであろうか?
それよりも、「美」的情操教育、音楽、美術
前頭葉の活性化「真」「善」(丸山)
そして、医療においては、
人間の精神生活や生き生きした交わりから生まれる情動を大切にし、患者をトータルな人格をもった全体像として捉える医学教育が望まれる。
脳活動(高次神経活動)の産物の総体として動物の心、ヒトの精神と呼ばれるものを位置づけることが出来る。そして、絵画、彫刻、音楽、文学、科学、教育を含めた文学と芸術と学問によって、つまり広く人間の精神の活動とのの所産によって、人間社会の中で形成され抽象化された概念(Begriff)を理解し創造しつつ人間はこれを次の世代へと伝達することができる。繰り返すが、脳の活動の所産が精神活動といわれるものであり、その具現化されたものが文化・文明の社会を形成する。そして、文化が継承される。また、精神の異常ないし病的状態を生物学的に研究することが、精神医学を科学的に研究することに他ならない。このような広い視野に立って、将来の脳と精神の科学を結びつけて発展させていく義務と責任が良心的知識人、研究者、芸術家に荷せられている。
人間とは何であるかを問い定義し、自らの手によって、それがあればこそ人間の人間たる所以であるところのものを証しつつ生きることを義務づけられた動物であるからであり、ひとたび、それこそが人間をして人間たらしめているところのものが定位されたときに、人間は各自の責任において、これを遵守しつつ生きてゆかねばならない。何故ならば、そうでない限り、人間の各自が、人間であることを自ら放棄してゆくことになるからであり、その意味で脳研究や芸術探求の目的がどこに向けられるべきかに改めて論議の的は絞られるように思われる。人間の高められた精神の中核が脳であるならば、生物学的意味での存在をも含めた一個の全体存在としての人間を人間として在るべき形姿=存在の目的へと導き得るものこそ脳を措いて他に在りはしないからである。
13]エピローグ(Epilogue)
よい芸居というものは、主たる役者が終幕でノコノコと出て来て、くどくどと述べたりはしないもの(お気に召すまま、As you like it. シェークスピア)であるが、この草稿にはここに言い残した文句が必要なようである。
Spoken by Rosalind: It is not the fashion to see the lady the epilogue; but it is no more unhandsome than to see the lord the prologue. If it be true that good wine needs no bush, 'tis true that a good play needs no epilogue; yet to good wine they do use good bushes, and good plays prove the better by the help of good epilogues. What a case am I in then, that…
丸山と川村の共同執筆となった経緯について
筆者らの一人川村は、精神医学と神経科学とくに神経解剖学を医学部卒業後約10年間学んだ。その前半は7:3,後半は2:8位の割合で臨床と実験をつづけてきた。長年多くの先輩や同僚に教えを受けてきたが、その中で最も尊敬する2人の学者について特記しておきたい。お一人は精神科医の臺 弘先生で、先生からは1990年を過ぎた頃から「精神に進みなさい」と時折励まされていた。また、留学中(1971-73年)からの恩師で神経解剖学者のAlf Brodal教授からは、「機能学は形態学をベースにしなくてはならない。神経科学の分野は広範囲であり、狭い分野に閉じこもらずに研究できるのは羨ましい。臨床医学とくに神経精神医学の研究を捨てるべきではない。」と暖かい忠告をうけた。1987年、先生は、癌で亡くなられる前年、ご自宅に一週間招いて下さって教え諭して下さった。
1999年正月に「脳と精神はどこで出会うか」を2000年正月に、「Gestalt心理学がおもしろい」という年賀状のメッセージを臺先生からいただいた。こうして川村は大学生活の定年(2000年3月)を迎えるにあたり、脳と精神の問題について考えをまとめ、それを最終講義で学生たちに披瀝して批判して貰うことに秘かに決めた。そして実際2000年2月17日(木)、川村は慶應義塾大学医学部に於いて『脳と精神は如何に出会うか』と題して講演を行った。
川村は約40年間、大脳皮質連合野、小脳求心系、脳の可塑性・移動、小脳、間脳、大脳皮質の発生の研究を共同研究者に恵まれて、伸び伸びと行うことが出来た。精神医学についても関心を持ち続け、時々ではあるが精神鑑定医、指定医として病院で働いてきた。
脳と精神を結びつけるに際して、川村は、脳の形態学、発生学的研究の成果の上に立脚して、できるだけ自分が関与した研究を通じて思考し、発表した内容を取り入れて論考すべく努めようとした。脳を研究する自然科学者として、事実に基づかない観念論的主張は排除しなければならないと考えつづけてきた。
このように考えて、1999年から2000年にかけて、なるべく多く、自分の専門以外の分野の発表を聴くことにした。討論にも積極的に参加した。そのような集会の中で、最も実りのあったのは、国際視覚シンポジウム(2000年3月8~10日、岡崎)であった。私自身も脳の機能と構成における「聴覚系と視覚系の特殊性と類似性」という題で発表した。外国から招待された何人かの旧友にも逢って最先端の研究内容をじかに知ることが出来た。この数年、実感として川村が得たものは、①視覚系に比して聴覚系の研究が遅れていること、②高次神経活動のうち認識系に比して情動系の研究が遅れていること、③音の響きと情操という要素の上に立つ音楽という芸術/学問について、音楽家、作曲家、演奏家の関心に応えられる確かな研究を自然科学者はほとんど持っていないことであった。それ故、この分野で現時点で得られているものを考察していく過程で、脳と精神の問題をまとめてみようと思った。何故なら、音楽という芸術の表現は、脳の高次神経活動の生み出した精神の活動に一致するものであると考えたからである。
川村は、脳の形態をベースに機能と結びつけて、この20-30年に関わりあった研究を再学習しながら、この著の大枠をまとめ上げるという仕事を楽しみながら2ヶ月余りで終わらせた。精神医学関連の内容については、臨床的感覚が鈍っていた。原著論文を調べて考察することが、にわかに出来なかったので、医学部卒業後10余年間精神医学教室や病院で、勉強した書物やノートの記録や同僚に教えを受けたりして、大筋だけは誤らないように努めて書いた。川村が最も苦心したのは趣味程度の教養すら身につけていない音楽に関する知識であった。しかし、これは芸術という切り口から精神についてまで容喙しようとする欲張った本書の趣意上、欠かせない部分なので、自力で小学生並の作文でよいから、ひとまず書いて形を整えてから先生を探すことにした。
川村が丸山に教えを乞うたのは、偶然ではなかった。川村は丸山のバッハ、ベートベン、モーツァルトに関する教養講座や、音楽大学における丸山のレッスンの聴講を受けていた。また、ヨーロッパ精神史についても、都内の教会で聴講を受けていた。川村は丸山にヒトの脳を実際に提示して教示し、しばしば討論を重ねた。丸山は川村を東京音楽大学のゼミナールに招き、脳と精神と音楽(音の響き)を中心とした討議を2000年5月20日に行なった。また、丸山は川村の定年退職記念研究会「脳と精神」に招かれて、「知覚と音の響きの二つの方向性について」と題して講演し、脳研究者との間に活発な討論が行われた。
丸山は、(personal historyなどを中心に書いて下さい。音楽家、宗教家/神学者として、等々)
以上のように、経歴も背景も、従って物の捉え方も異なる人生を歩んできた二人が、互いに専門の知識を教え合い、理解し合って、このような共同執筆という作業にまでこぎつけたのである。脳と精神と芸術(とひいては教育)について、非常にしばしば、各論的内容については勿論のこと、全体的・統括的な意見の交換を行った。共に50年~60年の人生を唯物論的、実証主義的思考を続けて来た川村と、精神と宗教と芸術の世界に生きてきて、精神の内的世界を思索してきた丸山との間に、この書で話題にしている問題に関しての共通した精神の志向があり、互いに違和感を感じてはいない。しかし鋭い読者は、Materialistの書いた文章とSpiritualistが書いた文章は容易に見分けることができるであろう。互いの専門性が滲み出ていて、文章のスタイルも、従って個性も、とくに結論の提示の仕方などにおいて異なってはいようが、この点に関しては、足して二で割る式の平板化をして脱個性的な統一した文体を作るということを敢えて試みなかった。むしろ、読者に、このような毛色の違った二人の、しかし同じ志向性をもった精神の持ち主の、書いた文体を多少ともとまどいながら笑みを浮かべてfollowして下さるであろうことを期待したい。それは分担執筆という形でまとめ得る本とは異なる風味を提供するものとなった。
[M.とS.の対話]:無しとする